Vista previa parcial del texto
¡Descarga Universidad Geneser libro y más Traducciones en PDF de Medicina solo en Docsity!
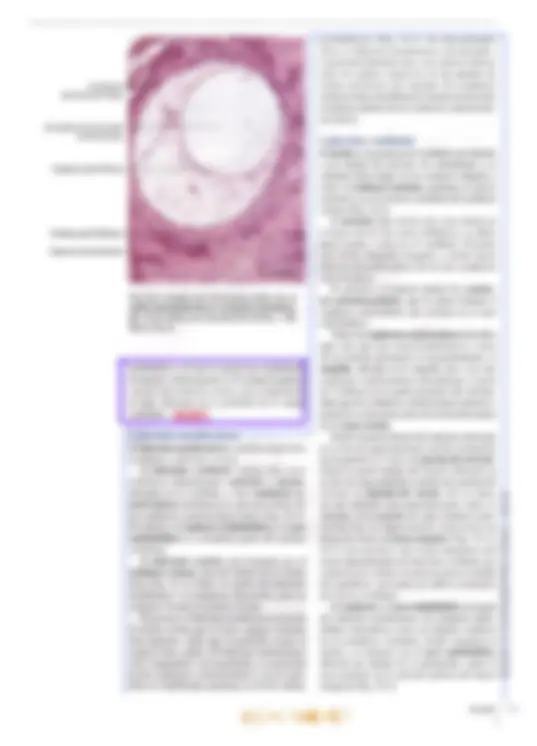
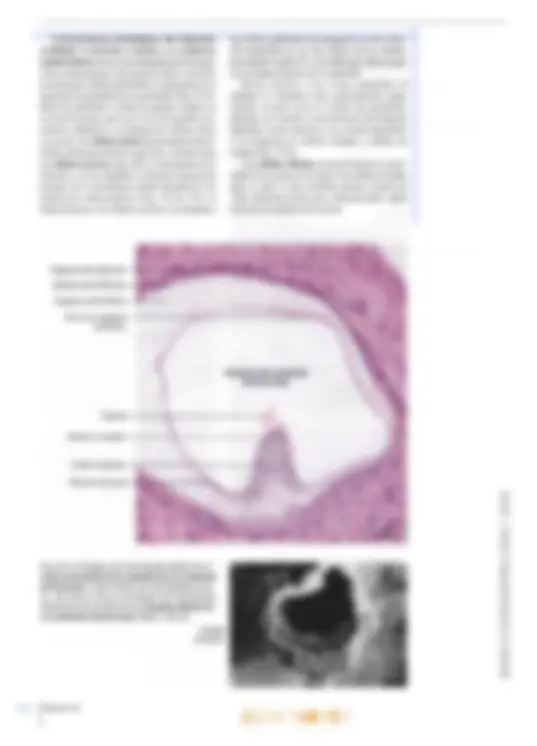
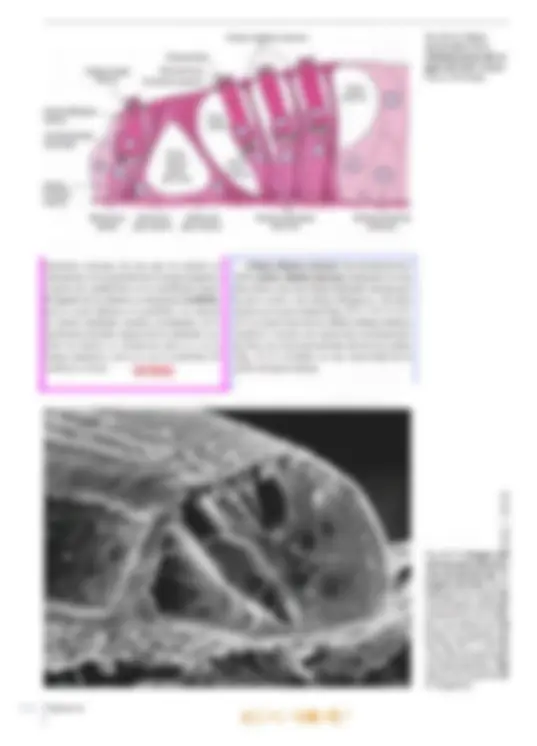
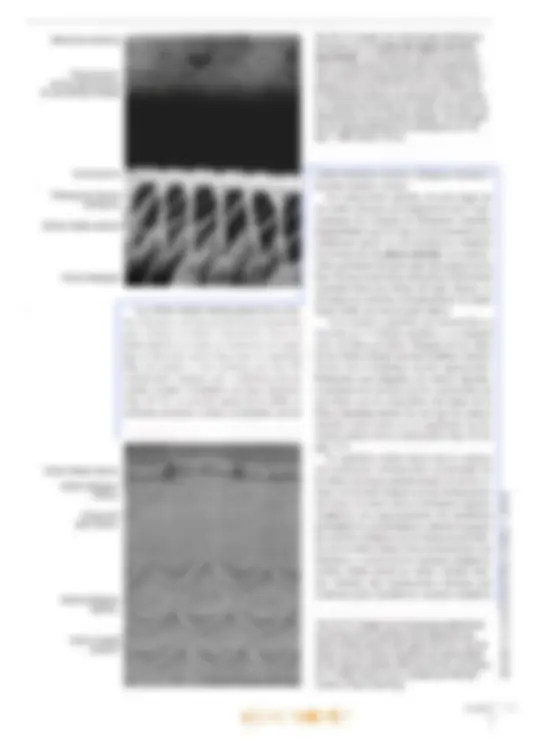
brusca disminución de la audición (“tapón en los oídos”). La disminución de la audición se debe a que la membrana timpánica se curva hi adentro o hacia afuera. ya sea que la presión del aire sea más baja o más alta que aquella en la caja del tímpano. Cuando la membrana timpánica se tensa, se modifican sus propiedades mecánicas, por lo que se transmiten sonidos más bajos a través del oído medio. Las infecciones pueden diseminarse desde la nasofaringe hacia el oído medio a través de las trompas auditivas y causar otitis media aguda, de gran importancia clínic: Oído interno El oído interno también se denomina laberinto, por su conformación compleja. Está compues- to por una porción ósea y otra membranosa. El laberinto óseo es un sistema continuo de cavidades y conductos en la porción petrosa del hueso temporal que rodea un sistema equivalente de sacos y conductos: el laberinto membranoso. Laberinto óseo El laberinto óseo se compone de tres partes: un vestíbulo que se comunica hacia atrás con los con- ductos semicirculares óseos, mientras que hacia adelante se continúa en la cóclea (Figs. 25-1 y 25-2). El vestíbulo y los conductos semicirculares pertenecen al órgano del equilibrio, mientras que la cóclea se relaciona con el órgano del oído. El laberinto óseo tiene una longitud de unos 2 cm, y el eje longitudinal coincide con el del hueso. En consecuencia, los conductos semicirculares se ubican en posición posterolateral, la cóclea en posición anteromedial y en el centro se inserta el vestíbulo. El laberinto membranoso está rodeado por el laberinto óseo. El espacio entre los laberintos óseo y membranoso se denomina espacio perilinfático y contiene la perilinfa. El vestíbulo es casi oval y en su pared lateral, orientada hacia la cavidad timpánica, se encuentra la ventana oval o vestibular, cerrada por la base del estribo. Los conductos semicirculares son tres, de los cuales el conducto semicircular anterior (o superior) se orienta perpendicular al eje lon- gitudinal del hueso, mientras que el conducto semicircular posterior se orienta a lo largo de ese eje. Así, ambos forman un ángulo de unos 457 con el plano medio. Por último, el conducto semicircular lateral se ubica en un plano perpen- dicular a los otros dos, con un ángulo abierto hacia adelante de unos 30? con el plano horizontal. Por al [def Má; aq E can perpendiculares entre 10 de los extremos de cada conducto posee una dilatación o ampolla ubicada justo después de la unión con el vestíbulo. La cóclea (lat. cochlea, caracol) es una dad ósea en espiral similar a un caracol. La ba tiene un diámetro de unos 9 mm y se orienta en dirección posterior y medial hacia el fondo del conducto auditivo interno. La curvatura basal de la cóclea sobresale dentro de la cavidad timpánica, donde forma el promontorio. Mide unos 5 mm de altura, y el vértice o cúpula se dirige hacia adelante y en dirección lateral, hacia la parte anterosuperior de la pared medial de la cavidad timpánica. En consecuencia, el eje de la cóclea es perpendicular al eje longitudinal del peñasco. La cóclea se compone de un pilar central cónico, el modiolo (lat., dim. de modius, buje de una rueda), alrededor del cual se enrosca un conducto óseo que describe dos giros y medio (Figs. 25-2 y 25-3). La base del modiolo se orien- ta hacia el fondo del conducto auditivo interno y presenta varias perforaciones que se continúan en delgados conductillos por donde transcurren las ramas del nervio auditivo, el nervio coclear. Desde el modiolo parte hacia la luz de la cóclea una delgada repisa ósea, la lámina espiral ósea, que sigue por toda la espiral, desde la base hasta el vértice. Los conductillos longitudinales del modiolo se curvan sucesivamente hacia afuera y llegan hasta el borde fijo de la lámina espiral ósea, donde cada uno presenta un ensanchamien- to. En conjunto, los ensanchamientos forman el conducto espiral del modiolo o conducto de Rosenthal, donde se encuentra el ganglio espiral o ganglio de Corti (Fig. 25-3). Los con- ductillos continúan desde los ensanchamientos del conducto espiral hasta el borde libre de la lámina espiral. La lámina espiral ósea (Fig. 25-3) penetra sólo hasta la mitad de la luz coclear, por lo que divide en forma incompleta el conducto en dos seccio- nes: una superior, el conducto coclear o rampa media, y una inferior, la rampa timpánica. Una membrana de tejido conectivo, la membrana basilar, se extiende desde el borde libre de la lámina espiral Ósea hasta la cresta basilar, un engrosamiento del ligamento espiral en la pared lateral de la cóclea, por lo que divide por com- pleto el conducto coclear de la rampa timpánica. Sobre el conducto coclear se encuentra la rampa vestibular. Las rampas timpánica y vestibular se comunican en el vértice del caracol a través de un orificio denominado helicotrema (gr. helix, caracol; trema, abertura). La rampa vestibular comienza en la parte basal de la cóclea en el vestíbulo, frente a la ventana oval (cerrada por la base del estribo), vi El oído 699 700 mientras que la rampa timpánica comienza a nivel de la ventana redonda (cerrada por la membrana timpánica secundaria). Un delga- do conducto, el acueducto coclear, comienza cerca de la rampa timpánica y desemboca sobre la cara interna de la porción petrosa del tem- poral, donde la perilinfa se comunica en forma inconstante con el líquido cefalorraquídeo del espacio subaracnoideo. Desde el punto de vista histológico, el hue- so compacto de la porción petrosa forma una condensación compacta especial de 2-3 mm de espesor alrededor de las cavidades del laberinto, la cápsula del laberinto, por lo que esa estructura puede disecarse. Las cavidades del laberinto óseo (el espacio perilinfático) están revestidas por periostio, pero en su interior se encuentra una capa de células muy aplanadas, las células perilinfáticas, de origen mesodérmico y con características de fibroblastos aplanados (Fig. 25-4). Una capa similar reviste también la superficie externa del laberinto membranoso. Finas prolongaciones cito- plasmáticas de las células perilinfáticas recorren el espacio perilinfático en el vestíbulo y parte de los conductos semicirculares y lo transforman en un sistema continuo de hendiduras intercelulares que contiene perilinfa. La perilinfa tiene una composición bastante similar a la del líquido cefalorraquídeo (conteni- do bajo de K* y elevado de Na*). Posiblemente provenga en gran medida de un ultrafiltrado del plasma sanguíneo por trasudación desde los capi- lares del tejido conectivo bajo la capa de células Ligamento espiral Membrana basilar Ventana coclear con membrana timpánica secundaria Ganglio espiral (de Corti) Lámina espiral ósea: Capítulo 25 an ras OPICENTRO USAC / 2016 Fig. 25-3. Imagen con O microscopio óptico de y) un corte mediomodiow- lar de la cóclea (de urO cobayo). Corte teñido O con hematoxilina-eosi- na. x37. Barra: 200 m. Características histológicas del laberinto vestibular, El utrículo, el sáculo y los conductos semicirculares poseen una delgada pared transpa- rente compuesta por una capa de tejido conectivo revestida por células perilinfáticas aplanadas en la superficie orientada hacia la perilinfa (Fig. 25-4). Hacia la endolinfa, se observa epitelio simple, en su mayoría plano, pero que cerca del epitelio sen- sorial es cilíndrico y se compone de células claras y oscuras. Las células claras no presentan caracte- ¡cas ultraestructurales especiales, mientras que las células oscuras (que sólo se encuentran en el utrículo y en las ampollas) contienen numerosos pliegues de la membrana celular basolateral con numerosas mitocondrias (Fig. 25-5a). Por su ultraestructura, las células oscuras se asemejan a Cápsula del laberinto Células perilinfáticas Espacio perilintático las células epiteliales de transporte en otros sitios del organismo (p. ej., las células de los túbulos proximales renales) y es posible que intervengan en el mantenimiento de la endolinfa. En las máculas y las crestas ampulares, el epitelio es cilíndrico alto especializado como epitelio receptor para el sentido del equilibrio; presenta las mismas características histológicas generales en las máculas y las crestas ampulares y se compone de células ciliadas y células de sostén (Fig. 25-6). Las células ciliadas son las receptoras senso- riales; se reconocen dos tipos: las células ciliadas tipo 1 y tipo 2, muy similares desde el punto de vista ultraestructural pero diferenciadas sobre todo por la distinta inervación. Zona con epitelio cilíndrico Epitelio receptor- Cresta ampular- Células oscuras: Fig. 25-5. a Imagen con microscopio óptico de un corte transversal de la ampolla de un conducto semicircular. Corte teñido con hematoxilina-eosi- ma. x90. Barra: 100 um. b Imagen con microscopia electrónica de barrido de una ampolla abierta en un conducto semicircular. Barra: 100 um. Capítulo 25 Ampolla del conducto Cúpula A semicircular anar BOOK'S COPICENTRO USAC / 2016 Las células ciliadas tipo 1 tienen forma de pera, con un estrecho cuello apical y una ancha base redondeada que contiene un núcleo redondo (Fig. 25-7a). Salvo en la parte apical, la célula está rodeada por una terminación nerviosa aferente con forma de copa, el eáliz, pero también es inervada por terminaciones eferentes menores. La superficie libre orientada hacia la endolinfa está recubierta por abundantes estereocilios (Figs. 25-Sb, 25-7b y 25-8b). Con microscopio electrónico se distinguen numerosas mitocondrias, un aparato de Golgi su- pranuclear bien desarrollado y muchas vesículas pequeñas (Fig. 25-7). La superficie libre luminal orientada hacia la endolinfa está recubierta por unos 30-50 estereo- cilios (microvellosidades largas inmóviles) y un cilio único denominado cinocilio. Vistos desde la luz, los estereocilios se disponen en un patrón hexagonal regular, pero además están polarizados, dado que crecen gradualmente en longitud en di- rección al cinocilio, que nace en uno de los bordes de la célula (Fig. 25-7a). Los estereocilios son más estrechos en la base y contienen un haz longitudinal de filamentos centrales, anclados a una red terminal en el citoplasma apical. El cinocilio presenta la habitual estructura interna de microtúbulos (patrón 9 +2) y está anclado en un cuerpo basal apical. El citoplasma del cáliz contiene abundantes mitocondrias y varias vesículas. Los cálices son los ensanchamientos terminales de las fibras ner- viosas vestibulares aferentes y el cáliz representa una forma de contacto sináptico con la célula ciliada tipo 1. Los cálices pueden rodear algunas células ciliadas tipo 1. Se ha demostrado que las Cúpula — Células. ciliadas Células de sostén am" e OA Fig. 25-6. Imagen con microscopio óptico del epitelio receptor especializado en una cresta ampular. Corte teñido con hematoxilina-eosina. x735. Barra: 10 ym. s EN anar terminaciones en cáliz están formadas por las fibras nerviosas vestibulares gruesas de conduc- ción rápida, En consecuencia, se considera que las células ciliadas tipo 1 tienen mayor capacidad de discriminación que las células ciliadas tipo 2. Las células ciliadas tipo 2 son cilíndricas, con núcleos redondeados ubicados a distintos niveles, pero por lo general más cerca de la luz que los de las células tipo 1. Desde el punto de vista citológico, los dos tipos celulares son casi idénticos, con una superficie luminal recubierta por estereocilios y un cinocilio. Las células ciliadas tipo 2 se diferencian de las células tipo 1 por el tipo de inervación. Sólo la parte basal de las células ciliadas tipo 2 está en contacto con las terminaciones nerviosas, pero cada célula es inervada por numerosas termina- ciones aferentes y eferentes. Las células de sostén son cilíndricas altas y se extienden desde la membrana basal hasta la superfi- cie luminal (Figs. 25-6 y 25-7a). Los núcleos redon- dos tienen ubicación basal y el citoplasma contiene un aparato de Golgi bien desarrollado, numerosas vesículas (posiblemente de secreción) y abundantes mitocondrias. Además, se observa una densa red microtubular longitudinal que corre desde la base hasta una red terminal por debajo de la superficie apical. Las superficies laterales se comunican con las células de sostén vecinas y con células ciliadas mediante complejos de unión. Las células de sostén confieren sostén mecánico a las células ciliadas y también tendrían importancia nutricional. Las prolongaciones (estereocilios y cinocilios) de las células ciliadas se extienden hacia arriba en una sustancia gelatinosa con glucosaminoglu- canos abundantes que recubre la cara superficial de las máculas y las crestas ampulares. En las máculas, esta masa es bastante aplanada y se denomina membrana otolítica (Fig. 25-8), que en su parte superior presenta numerosos cuerpos cristalinos denominados otolitos o estatoconios (lat. status, posición, equilibrio; gr. konia, polvo). En su mayor parte, se componen de carbonato de calcio y su tamaño varía hasta un máximo de 30 um. Su peso específico es casi tres veces su- perior al promedio de la membrana otolítica, por lo que aumenta notablemente su masa. A lo largo de ambas máculas, por la línea media transcurre (Z una cresta con forma de semiluna (ing. snowdrift 7 line, línea de nieve) que se corresponde con una Ú] banda angosta subyacente en la capa de células Y epiteliales sensoriales denominada estriola (Fig. 25-8c), donde la densidad de células sensoriales O es un 20% inferior a la del resto de la mácula. 0) Cc /2016 O US) osa con abundancia de glucosaminoglucano: 's mucho más gruesa y se denomina cúpul: Figs. 25-5a y 25-6). Se extiende con forma El oído Fig. 25-8. a Dibujo esquemático de una mácula en el laberinto vestibular. (Según lurato). b Imagen con microscopio electró- nico de barrido de la membrana otolítica y el epitelio receptor en el utrículo. Barra: 100 jm. (Cedido por Michael Larsen y Klaus Qvortrup). e Diagrama esquemático que mues- tra las características morfológicas de las máculas del utrículo y el sáculo, respectiva- mente. Nótese la distinta conformación de las dos máculas y la diferente orientación de los haces de estereocilios: en el utrículo, los estereocilios están orientados hacia las estriolas, y en el sáculo, están orientados en dirección opuesta alas estriolas. (Según Gray, en Williams). Membrana otolítica Célula ciiada —N tipo 1 Célula de sostén Fibras nerviosas Terminación eferente Capa: gelatinosa Fibra nerviosa vestibular c El saco endolinfático contiene en su luz una sustancia homogénea y, con microscopia óptica, se observa que se asemeja a un folículo tiroi- deo. Está revestido por epitelio cilíndrico alto compuesto por dos tipos de células, a saber, con abundancia de ribosomas y con abundancia de mitocondrias. Las células con abundancia de ribosomas contienen gran cantidad de retículo anar Membrana otolítica Célula ciliada Utrículo Célula ciliada tipo 2 Terminación eferente Terminación: aferente Línea de “nieve” a Otolitos DE 7 Ep] Células lolo) Moe (6) sensoriales JEREO vestibulares endoplasmático rugoso, varios aparatos de Gol; y vesículas de secreción, cuyo contenido es ss cretado a la sustancia homogénea intraluminal Al igual que las células tiroideas, las célul principales muestran indicios de capacidad d endocitosis, dado que la sustancia homogén es captada por endocitosis según necesidad. capa de tejido conectivo subyacente del saco ex BOOK'S COPICENTRO USAC / 2016 El oído lolinfático está muy vascularizada, con capilares fenestrados, Por lo tanto, por su ultraestructura el saco endolinfático se asemeja a una glándula endocrina y se cree que las células con abundancia de ribosomas producen una hormona que podría tener importancia para la regulación del contenido de electrolitos de la endolinfa. Posiblemente la hormona se almacene como prohormona en la sustancia homogénea. Las células con abundan- cia de mitocondrias están densamente ocupadas por mitocondrias, y la superficie luminal está re- las, las células ciliadas son estimu- ladas po iceleración líneal y la fuerza de gravedad. que la membrana otolítica tiene mayor peso Ífico que la endolinfa, una aceleración li cualquier dirección, por de la cabeza, induce un ina otolítica respecto lo que se inclinan Tean diferencias ejemplo la inclin4 movimiento de la de las células ciliada los cilios. De este mod de potencial en las célula mencionó, el cinocilio tiex de la longitud en dirección al ci ha demostrado que la inclinación de reocilios en dirección del cinocilio call despolarización de la célula ciliada, seguid una mayor frecuencia de impulsos en la IB nerviosa aferente, mientras que la inclinació de los estereocilios en dirección opuest cinocilio genera una hiperpolarizació: célula ciliada y la consecuente redug 'anal iónico, cuyo menta con la incli- odo de apertura disminuye hacia el lado opuesto al ci- linfa se produce un aumento del flujo es potasio hacia el interior por el período Capítulo 25 cubierta por numerosas microvellosidades largas que confieren a las células una gran superficie y un aspecto correspondiente a las células absortivas. Se cree que absorben y transportan la endolinfa hacia los capilares de la capa de tejido conectivo. La endolinfa ocupa todos los espacios co- municantes del laberinto membranoso y posee una composición química particular, dado que es el único líquido extracelular del organismo con muy elevada concentración de iones potasio (140 mM) y bajo contenido de sodio (5 mM). En de los ¡ones potasio, tambié calcio, lo cual se cree que jj de un transmisor sináp En las crestas a das son estimul: AA ria fa ampular y la pared de los conductos rculares, el extremo libre se desplaza en :ción opuesta al movimiento de la cabeza de los conductos semicirculares), puesto ¡e la endolinfa presiona a la cúpula en la g dirección, por efecto de la inercia. En uencia, se desplaza como una puerta, ¡e los cilios se inclinan en la dirección movimiento de la cabeza. La in- los cilios causa despolarización e ambién en las crestas ampula- idad de reposo en las fibras movimiento rotatorio, duce en la cúpula un respectiva se modifica entonc8 opuesta. Como la cúpula tiene exactame! mismo peso específico que la endolinfa, ciliadas en la cresta ampular no son al das por las aceleraciones lineales o la fuel gravedad. anar BOOK'S COPICENTRO USAC / 2016 708 Br El vértigo post igaroxístico benigno (BPPV, ing. benign PR pal positional vertigo) se considera la fork fiecuente de vértigo periférico, es decir, gent € nervio vestibulococlear o el órgano del equ brio. Como regla general, el vértigo pg se reconoce por su características MÓnes de movimiento con típica 59 Prde rotación (vértigo giratorio). 4 Riel paciente o el entorno. El BJ Jere comenzar sin motivo alguno PISO vértigo rotatorio cuando se sal Pábeza en una posición determinada. través del conducto reuniens. El extremo ciego se ubica junto al helicotrema. En un corte transversal, el conducto coclear tiene forma triangular y se describen un techo, una pared lateral y un piso (Figs. 25-9 y 25-104). El recho separa el conducto coclear de la rampa vestibular y está formado por la delgada mem- brana vestibular (membrana de Reissner), que se extiende desde el periostio engrosado sobre la lámina espiral hasta el ligamento espiral sobre la cara interna de la pared externa de la cóclea. La pared lateral del conducto coclear está formada por el ligamento espiral, que es el pe- riostio engrosado sobre la superficie interna de la cóclea a nivel del conducto coclear. La mayor parte del ligamento espiral está especializado en forma de la estría vascular, que se extiende desde la fijación de la membrana vestibular hasta una elevación redondeada en el conducto coclear: la prominencia espiral. El piso del conducto coclear está formado por la lámina espiral ósea y la membrana basilar fijada en la parte medial al borde de la lámina espiral. En dirección basal de la cóclea, el ligamento espiral presenta un saliente parcialmente óseo en forma de peine, la eresta basilar, en cuya cara lateral se inserta la membrana basilar. La cara inferior del piso se orienta hacia la rampa timpánica. La parte superior presenta una estructura muy espe- cializada, el órgano espiral (órgano de Corti), que es el órgano receptor del sentido del oído. Al igual que las demás partes del laberinto membranoso, el conducto coclear contiene endolinfa. Estructura del laberinto coclear. A continua- ción se describirán con mayor detalle las carac- terísticas histológicas de las distintas estructuras de las paredes del conducto coclear. La membrana vestibular (membrana de Reissner) se compone de dos capas epiteliales Capítulo 25 Es probable que el BD Ríéba a que restos de otolitos qus dos en la cúpula de la Cresta aa fé Uno de los conductos semici cul IST estimulan los nervios aferentes sin RIVO. La causa más frecuente de BBPV es el sismo cefálico. El tratamiento mediante la mA ds Epley (procedimiento de reubi- cación de TW alíticos), donde se mueve la cabeza en secúW fp cuatro posiciones definidas durante 30 sc iguede liberar los restos de otolitos capturada Lo que cesa el vértigo. aplanadas (Figs. 25-9 y 25-10). El epitelio sim+ ple plano muy delgado orientado hacia la rampa: vestibular está formado por células perilinfáticas de origen mesodérmico que carecen de lámina basal. Hacia el conducto coclear, también hay epitelio simple plano (de origen ectodérmico), con características ultraestructurales que sugieren una función de transporte de líquido y electrolitos; El epitelio tiene una lámina basal que lo separa del mesotelio, El ligamento espiral no es un ligamento ver= dadero, sino una gruesa capa de periostio en la cóclea ósea. Hacia el hueso, el ligamento espiral se compone de tejido conectivo denso colágeno! mientras que hacia el interior del conducto coclear su estructura es más laxa y está más vascularizado; Conforma la mayor parte de la pared lateral del conducto coclear. Estría vascular. Hacia el conducto coclear, el ligamento espiral está recubierto por un epitelid estratificado denominado estría vascular (Figs; 25-9 y 25-104). El epitelio se compone de tres capas de células, denominadas respectivamente células marginales, células intermedias y células basales. Las células marginales tienen origen ectodérmico. Se desconoce el origen dé las células basales, pero se cree que sería mes+ odérmico o que provendrían de la cresta neural. Algunas de las células intermedias se asemejan d los melanocitos, por lo que se cree que derivarían de la cresta neural. No hay lámina basal entre las células basales y el tejido conectivo subyacente en la estría vascular, Hacia arriba, las células marginales se continúan en el epitelio de la membrana vestibular, mientras que hacia abajo se continúan en el epitelio de la prominencid espiral. Desde el punto de vista ultraestructural, las células marginales se caracterizan por su' elevado contenido de mitocondrias. También se encuentran numerosos pliegues del plasmalema BOOK'S COPICENTRO USAC / 2016 Rampa vestibular Ligamento espiral Estría vascular Capilares intraepiteliales Conducto coclear Células limitantes externas Cresta basilar Órgano de Corti Prominencia espiral +++ Rampa timpánica $ Fig. 25-10, a Imagen con microscopio óptico de un corte trans- versal del conducto coclear. Corte incluido en plástico epon, teñido con azul de metileno. x150. Barra: 100 pm. b Imagen con micros- copio electrónico de barrido del conducto coclear. La membrana tectoria se ha levantado como consecuencia del proceso de des- hidratación durante la preparación de la muestra. x220. Barra: 100 tm. (Cedido por Michael Larsen y Klaus Qvortrup). Membrana vestibular Membrana tectoria-É Órgano de Corti en la superficie basal y cortas microvellosidades hacia la luz. Por su aspecto, las células margina- les se asemejan a las células que intervienen en el transporte de sales y agua (p. ej.. las células de los túbulos proximales renales). Es muy pro- bable que contribuyan con la producción de la endolinfa que, como se mencionó, tiene elevada concentración de ¡ones potasio. En los pliegues basales de las células margina altas concentraciones de ATPasa de Na'-K*, y además aquí se encuentra una bomba de potasio que interviene en el mantenimiento del 1 endococlear positivo. es se encuentran - FADAS Membrana vestibular Células interdentales Limbo espiral Membrana tectoria Células limitantes internas Lámina espiral ósea Pe Pay Ganglio coclear Las células intermedias y basales sólo se di- ferencian por su ubicación y no por su citología. El citoplasma es pálido, con contenido aislado de mitocondrias, muchas vesículas y gránulos de melanina dispersos. Envían largas prolonga- ciones citoplasmáticas hacia la porción basal de las células marginales. Las células intermedias al parecer tienen importancia para el funciona- miento normal de la estría vascular. En relación con ciertos tipos de hipopigmentación (p. ej., el síndrome de Waardenburg o el síndrome de Tiet- ze), a menudo se observa pérdida de la audición o hipoacusia. El oído BOOKS COPICENTRO USAC / 2016 Fig. 25-11. Imagen con microscopio óptico de un corte transversal del conducto coclear. Corte incluido en plás- tico epon, teñido con azul de metileno. 400. Barra: 10 ym. Membrana tectoria Célula ciliada interna Célula del pilar interno Túnel interno (de Corti) Célula falángica interna las fibras va seguido de un estrechamiento de la lámina espiral ósea y un aplanamiento de la cresta basilar. Mientras que la longitud disminuye desde el vértice hacia la base, el diámetro de las fibras aumenta de 0,5 um en el vértice a 1,5 um en la base; desde el punto de vista biomecánico, esto incrementa la rigidez de cada fibra unas 100 veces. Aunque los filamentos de las cuerdas acústicas son de colágeno, desde el punto de vista biomecá- nico las estructuras son elásticas. Los movimientos vibratorios en la base del estribo sobre la ventana oval desencadenan una onda vibratoria en la pe- rilinfa de la rampa vestibular que es transmitida a la membrana basilar y los consiguientes movi- mientos del órgano de Corti estimulan las células ciliadas receptoras del sonido. De acuerdo con la frecuencia del sonido, una zona determinada de la membrana basilar alcanza una oscilación máxima correspondiente a la distinta longitud y rigidez de las cuerdas acústicas. Órgano de Corti. Sobre la cara superior de la membrana basilar, el epitelio del conducto coclear sufre un engrosamiento y forma el verdadero órgano receptor de la audición, denominado órgano de Corti (Fig. 25-9) u órgano espiral, que se extiende en todo el recorrido del conducto coclear, por lo que en tres dimensiones presenta características de una cresta epitelial en espiral ). El órgano de Corti es muy complejo, ¡plificar, las células pueden dividirse : las células ciliadas y las células de 5-11 y 25-12). Las células ciliadas son los receptores senso- riales y se observa una hilera de células ciliadas internas y 3-5 hileras de células ciliadas externas. Las células de sostén se diferencian en 6 tipos. denominadas de adentro hacia afuera: células limitantes internas, células falángicas internas, Habénula perforada Vaso espiral anar células columnares internas o del pilar interno, células columnares externas o del pilar externo, células falángicas externas (células de Deiters) y células limitantes externas (células de Hensen). Todas las células de sostén tienen una membra- na basal que las separa del tejido conectivo de la membrana basilar. La porción apical llega hasta la superficie libre del epitelio, donde en especial las células ciliadas, las células de los pilares y las células falángicas están relacionadas mediante complejos de unión muy estrechos. En su citoplas- ma apical poseen condensaciones de microtúbulos y microfilamentos que, junto con los complejos de unión y con microscopia óptica, se visualizan como una delgada estructura continua similar a una membrana, la membrana reticular, a través de la cual pasan los estereocilios de la superficie de las células ciliadas (véase la pág. 714). A diferencia de las células de sostén, las célu- las ciliadas no llegan hasta la membrana basilar, dado que las células ciliadas internas están sepa- radas de ella por las células falángicas internas, mientras que la parte basal de las células ciliadas externas está rodeada por las células falángicas externas (Figs. 25-12 y 25-14). Se encuentran varios espacios entre las células del órgano de Conti (Figs. 25-11, 25-12 y 25- 13). Las células de los pilares interno y externo están separadas por un espacio triangular al corte transversal, el cunículo o túnel interno (túnel de: Corti), que se comunica a través de las hendiduras entre las células del pilar externo con el cunículo 0 túnel medio (espacio de Nuel), que se encuentra entre las células del pilar externo y las células ciliadas externas. A su vez, el túnel medio se comunica con el cunículo o túnel externo, que se encuentra entre la hilera más externa de las células ciliadas y la hilera más interna de células Célula ciliada externa Célula del pilar externo 7 Célula falángica externa g Célula de Hensen Célula de Claudius El oído BOOK'S COPICENTRO USAC / 2016 Estereocilios Red terminal Contacto oclusivo- Célula ciliada interna Célula falángica interna Terminaciones nerviosas Célula limitante Células ciliadas externas interna , + Membrana Célula del Célula del basilar — pilarintemo pilar externo limitantes externas. Se cree que los túneles se comunican con la perilinfa de la rampa timpánica a través de conductillos en la membrana basal. El líquido de los túneles se denomina cortilinfa, pero es (casi) idéntica a la perilinfa. Los túneles se cierran mediante zonulae occludentes en la membrana reticular respecto de la endolinfa. Así, todos los túneles se comunican entre sí y con la rampa timpánica, pero no con la endolinfa del conducto coclear. Capítulo 25 NÑ Células limitantes externas Nx” Células falángicas externas Células ciliadas cocleares. Se encuentran unas 3500 células ciliadas internas dispuestas en una ca hilera entre las células limitantes internas por la parte medial y las células falángicas y del pilar interno por la parte lateral (Figs, 25-11, 25-12 y 25- 15) La región basal de las células ciliadas internas establece contacto con numerosas terminaciones de fibras nerviosas provenientes del nervio coclear 5-12) incluidas en una concavidad de la la falángica interna. Fig. 25-12. Dibujo esquemático de la ultraestructura del ór- gano de Corti. (Según Ham y Cormack). SAC / 2016 Fig. 25-13. Imagen corl microscopía electr: nica de barrido del E órgano de Corti. Se z. distingue con nitidez la] conformación tridime: sional de los tres tún: les y las células que li limitan (compárese cop, las Figs. 25-11 y 25-1: y las denominaciones) correspondientes). (08) dido por B. EngstrámSA H. Engstróm). 714 fuera de las regiones sinápticas, en las que se comprueban condensaciones pre y postsinápticas de la membrana. Estas terminaciones sólo rara vez establecen contacto con las células ciliadas, pero: sí lo hacen con las terminaciones de las fibras nerviosas aferentes, y su función es modular la actividad en las terminaciones aferentes.Las casi 12.000 células ciliadas externas se ubican en la periferia de las células del pilar externo, donde forman 3-5 hileras regulares (Figs. 25-11 y 25- 15). Presentan una forma cilíndrica más alargada que la de las células ciliadas internas y sólo están revestidas en su tercio basal por las células falángicas externas (Fig. 25-14), que también se disponen en 3-5 hileras. El núcleo se ubica en la región basal de la célula ciliada, y al igual que en las células ciliadas internas, en la porción apical se distingue una condensación que forma una placa cuticular. En la zona más basal, las células ciliadas externas están en contacto con numerosas terminaciones ensanchadas provenientes de las fibras nerviosas del nervio coclear, y aquí la célula ciliada está separada de la correspondiente célula falángica externa por un espacio en el que se en- Cuentran las terminaciones nerviosas cocleares. UNAUCHO plasma apical del los complejos de y de sostén. Los fila enlaces cruzados entre los filamentos de al tiene propiedades elásticas, y tropomiosina q se fija a la actina y confiere efecto de ri; Las células falángicas y de los pilares tay Mediante microscopía electrónica, 4 se dispersan en di citoplasma cond Capítulo 25 a cuticular y membrana reticular superficie libre está cubierta por estereocilios, entras que, al igual que en las células ciliadas internas, no hay cinocilios. En la superficie libre de cada célula ciliada -neuentran unos 100 estereocilios dispuestoas e forma de V, con los extremos orientados haci; uera, en dirección al centríolo (Fig. 25-15). diferencia de las células ciliadas internas, los stereocilios más largos están incluidos en 1. ¡embrana tectoria suprayacente (Fig. 25-14). Anivel de la porción basal de las células cilia- las externas. se distinguen numerosas siluetas de lerminaciones nerviosas ensanchadas, que en esti :aso también pueden clasificarse en aferentes 'ferentes sobre la base del contenido de vesículas 'n el citoplasma presináptico y postsináptico. Si 'mbargo, aquí tanto las terminaciones aferente: omo las eferentes establecen contacto sináp- ico directo con las células ciliadas externas. n consecuencia, las fibras nerviosas eferentes »jercerían un posible efecto modulador sobre l: 'élula receptora sensorial. Células de sostén. El epitelio bajo del surcí 'spiral interno se continúa en dirección lateral :n las células limitantes internas del órgan: las falángicas ex- úír externo y las células fída por las placas cuticulares aciones de contacto entre los ¡lares antes mencionados (además 'spondientes porciones luminales de llas falángicas internas. las células del ár interno y las células ciliadas internas). 'n algunos casos, los sonidos intensos causar daño del oído interno. En co- :ncia con la frecuencia de estos soni- en aves, y los experimentos suB podría ocurrir neoformación de ct in vivo en mamíferos. anar BOOK'S COPICENTRO USAC / 2016 de Corti (Figs. 25-11 y 25-12). Se trata de varias hileras de células delgadas que en la transición con el epitelio del surco espiral interno son cilín- dricas bajas pero que crecen gradualmente hasta convertirse en cilíndricas muy altas en dirección hacia afuera. Las células falángicas internas se distribuyen en una sola hilera de células de sostén cilíndri- cas altas que están interpuestas entre las células ciliadas internas y las células del pilar interno (Fig. 25-11 y 25-12). El núcleo se ubica en la porción basal de la célula. La región apical de las células falángicas internas es muy angosta y se extiende hacia la superficie libre del epitelio, donde la célula presenta contactos oclusivos con las células ciliadas internas y las células de sostén vecinas (células falángicas internas, células del pilar interno y células limitantes internas). Cada célula ciliada interna está incluida en un surco de la célula falángica interna. Las células del pilar interno descansan so- bre la membrana basal con una base ancha, casi triangular, que limita hacia adentro con las células falángicas internas, mientras que hacia afuera está en contacto con la base de las células del pilar externo (Figs. 25-11 y 25 12). Desde la porción más interna de la base parte un cuerpo celular delgado con forma de columna o pilar, cuya su- perficie interna está totalmente en contacto directo con las células falángicas internas, mientras que la superficie externa se orienta hacia el túnel de Corti. El cuerpo celular se ensancha en dirección apical y adquiere un aspecto similar al del extremo proximal del cúbito (hueso del antebrazo), con una profunda concavidad en la que se ubica la porción apical ensanchada de la célula del pilar externo opuesta. El cuerpo celular con forma de columna crea un ángulo lateral abierto hacia afuera de unos 60* respecto de la base plana y. junto con el cuerpo celular de la célula del pilar externo (que forma con la base un ángulo menos abierto hacia adentro), conforma el túnel de Corti (túnel interno), de forma triangular. Fibras nervio- sas amielínicas para la inervación de las células ciliadas externas pasan a través del túnel de Corti. El núcleo se ubica en la porción externa de la base, mientras que en la porción interna se extien- de un denso haz de fibrillas que atraviesa todo el cuerpo celular hacia arriba y termina en una placa cuticular que se extiende sobre la porción apical de la célula del pilar externo. En rasgos generales, las células del pilar externo tienen la misma configuración que las células del pilar interno, con una base que descansa sobre la membrana basal. En dirección medial, limitan con la base de las células del pilar interno, mientras que en dirección lateral están en contacto con la base de las células falángicas anar externas (véanse las Figs. 25-11 y 25-12). La porción con forma de columna del cuerpo celular termina en dirección apical en un ensanchamiento que forma una prominencia convexa en dirección interna, que coincide con la concavidad en la porción apical de la célula del pilar interno. Una prolongación falángica luminal, de ubicación lateral, con forma de placa y similar a una falange, contiene una placa cuticular condensada y forma contactos oclusivos con células del pilar externo y falángicas externas vecinas. Los delgados cuerpos celulares de las células del pilar externo están separados por grandes hendiduras intercelulares. Las células falángicas externas (células de Deiters) son células de sostén de las células ciliadas externas y se disponen en un número equi- valente de hileras (Figs. 25-11 y 25-12). La parte basal cilíndrica de las células falángicas externas descansa sobre la membrana basal, donde las células se ubican muy juntas y limitan en direc- ción interna con las bases de las células del pilar externo y en dirección externa con las bases de las células limitantes externas. La porción cilíndrica de las células falángicas contiene el núcleo y en la parte apical posee una profunda concavidad que rodea el tercio basal de la célula ciliada externa, aunque separada de la porción más basal por un espacio intercelular en el que se encuentran las terminaciones del nervio coclear. En consecuen- cia, la porción cilíndrica de la célula falángica externa no llega hasta la superficie, pero la parte más externa emite una prolongación falángica larga y delgada que termina en un ensanchamiento aplanado que rodea las células ciliadas externas hacia la luz (Fig. 25-14). El citoplasma de la prolongación contiene numerosas fibrillas que, según se observa con microscopio electrónico, están compuestas por haces paralelos de micro- túbulos. Estas fibrillas parten de condensaciones en la porción basal de la célula y se extienden hacia arriba, a través de la porción cilíndrica, donde continúan en la prolongación falángica y se aplanan en una placa cuticular. Como ya se mencionó, los dos tercios apicales de las células ciliadas externas están separados de las células falángicas externas por los túneles -espacios intercelulares comunicantes-, por lo que estas porciones celulares se encuentran a consi- derable distancia entre sí (véase la Fig. 25-14). Las células limitantes externas (células de Hensen) son varias hileras de células cilíndricas que cierran el órgano de Corti en su parte externa (véanse las Figs. 25-11 y 25-12). Todas descan- san sobre la membrana basal y están muy juntas entre sí, aunque separadas de las células ciliadas externas y las células falángicas externas por el túnel externo. Las células limitantes externas El oído BOOK'S COPICENTRO USAC / 2016 anismo generador de variaciones del potencial de membrana en células ciliadas cobpres La incli de los cilios (véase la pág. 716) genera va- — y la consecuente inclinación hacia afuera de 1g riaciones d 'encial de membrana de las células ciliadas. — dirección del cuerpo basal) genera una despg El poten psitivo de la endolinfa se mantiene por la potencial de receptor actúa sobre la transn secreción conti le iones potasio, combinada con la reab- — las terminaciones aferentes en la base cel lo cual varía la sorción de ¡ones por las células marginales de la estría — frecuencia de los potenciales de acción :orrespondientes. vascular, La endolíí caracteriza por tener un potencial fibras nerviosas aferentes. Con la desp zación de la célula positivo respecto de rilinfa. El potencial es de unos — ciliada, aumenta la frecuencia de i ps en las fibras nervio- +80 mV en el conducidéBglear. Las células ciliadas tienen — sas, mientras que la hiperpolariz la disminuye. un potencial intracelular ivo de -70 mV respecto de la Dentro de cada haz, los e: lios están unidos por va- perilinfa. En conjunto, el pOBgjal transmembrana entre el — rios tipos de “enlaces” fibr xtracelulares (Fig. 25-16). potencial intracelular de las cél iliadas respecto de laen- Los enlaces laterales une fronco de un estereocilioa con dolinfa (en que están bañadas) es € s 150 mV. Es el mayor — el vecinoa. Los enlaces ales pasan desde la punta de un potencial de reposo del organismo, potencial denominado estereocilio al tronco estereocilio vecino más alto. La potencial generador contribuye sin d a la sensibilidad de i áni fos canales iónicos relacionada con la las células ciliadas a la deformación ¡ca y su función — inclinación de los $s está mediada por enlaces apicales que como transductoras. La señal mecánica ionada con los se fijan alas pgs de los canales iónicos mecanosensibles movimientos de los cilios se transforma en eñal eléctri- enlamembi Clular de los estereocilios. La inclinación de ca en la forma de difere: de potencial gel as por los — loscilios 'cción lateral aumenta la tensión en los enlaces canales iónicos mecanosensibles, cuyo período Bgrtura es apicales Úíal afecta la conformación de la proteína de canal afectado por la inclinación de los cilios (del mism: que en dipgión a la apertura y la consecuente despolarización, se describió para las células ciliadas vestibulares; véa: pág. mig que la inclinación de los cilios en dirección medial 706). En el movimiento hacia abajo de la membrana bi * los filamentos de los enlaces ena e inhibe la la correspondiente inclinación de los cilios hacia adentró ión en la proteína de canal, que decir alejándose del cuerpo basal del citoplasma apical AA. SU Emina una hiperpolarización del p MIPCOMO una unidad. No se conoce la composición os enlaces apicalesg y ni de las proteínas de canal. Cinocilio N Estereocilio h Enlace apical = “Conector apical” | ¡enlaces laterales | Ñ basales Y a cuticular ——— y Fig. 25-16. Enlaces de estereocilios (véase el texto para más detalles). (Según Forge). El oído anar BOOK'S COPICENTRO USAC / 2016 718 hacia abajo de la membrana basilar, la membrana reticular se desplaza hacia abajo y afuera, por lo que los cilios se inclinan hacia adentro porque las células ciliadas están fijadas a la membrana basilar, mientras que los estereocilios de las célu- las ciliadas externas están fijados a la membrana tectoria. Ante el consecuente movimiento hacia arriba de la membrana basilar, la membrana re- ticular se desplaza hacia arriba y adentro, por lo que los cilios se inclinan hacia afuera. Como se describió antes, las células ciliadas internas no están en contacto directo con la membrana tec- toria, y se cree que la inclinación de sus cilios es causada por las vibraciones de la propia endolinfa. En el nervio coclear, alrededor del 90% de las fibras son aferentes y el 10%, eferentes. La gran mayoría (90-95%) de las fibras aferentes forman si- napsis con las células ciliadas internas, mientras que alrededor del 90% de las fibras eferentes establecen sinapsis con las células ciliadas externas. Por lo tanto, se considera que las células ciliadas internas son los receptores de sonido primarios. En cambio, en estudios in vitro se demostró que las células ciliadas externas tienen propiedades contráctiles y pueden acortarse en un 10% de su longitud cuan- do se despolarizan, debido a la estimulación por medios mecánicos con la inclinación de los cilios 0 por estimulación eléctrica. La estimulación con sonidos de baja frecuencia causa la contracción y el acortamiento de células ciliadas externas ubicadas cerca del helicotrema, mientras que los sonidos de alta frecuencia causan la contracción de las células ubicadas cerca de la base del caracol, y las frecuencias intermedias causan la contracción de los niveles intermedios correspondientes. Se cree que la contracción de las células ciliadas externas tiene un efecto modulador de la audición, en parte debido a que refuerza los sonidos como consecuencia de la disminución natural de la atenuación existente en la membrana basilar, y en parte a que ocurre para determinadas frecuencias, lo cual implica un aumento de la sensibilidad de las células ciliadas Irrigación sanguínea del oído interno Nicol znagnay Irrigación arterial. El oído interno es irrigado por la arteria laberíntica, una rama de la arteria cerebelosa anteroinferior o de la arteria basilar. Parte hacia el conducto auditivo interno, donde se divide en tres ramas: la arteria vestibular, la arteria vestibulococlear y la arteria coclear. El saco endolinfático es irrigado por la arteria meníngea posterior: La arteria vestibular irriga el nervio ves- tibular y partes del utrículo, el sáculo y los conductos semicirculares. El resto del aparato vestibular recibe irrigación arterial de la arteria vestibulococlear que también nutre el giro basal del conducto coclear. La arteria coclear se divide en varias ramas menores que transcurren por los conductillos longitudinales del modiolo y emiten ramas sucesivas hacia la lámina espiral ósea y el ganglio espiral de los dos giros apicales de la cóclea y parte del basal. Desde la lámina espiral ósea, continúan las ramas hasta la parte interna de la membrana basilar. En el laberinto membranoso, las ramas ar- teriales forman redes capilares subepiteliales, aunque como ya se mencionó, en la estría vas- cular se encuentra una red capilar intraepitelial (véase la pág. 709). En la porción interna de la membrana basilar, se observa en forma cons- tante un capilar característico denominado vaso espiral (Fig. 25-11). Las venas presentan un recorrido diferente al de las arterias, sobre todo en la cóclea. En la có- Capítulo 25 clea, las ramas arteriales terminales transcurren sobre todo por la rampa vestibular, mientras que las venas menores aparecen en la rampa timpá- nica. Drenan desde el giro apical de la cóclea a la vena espiral anterior, mientras que los dos giros basales drenan a la vena espiral posterior. Al unirse, las dos venas espirales dan origen a la vena del acueducto coclear, que se vacía en la vena yugular. Los capilares del modiolo y el ganglio espiral forman la vena espiral del modiolo, que se abre a las venas laberínticas que terminan en el seno sigmoideo o en el seno petroso inferior. La sangre venosa del aparato vestibular drena en parte en las venas vestibulares que se vacían en las venas espirales, y en parte en la vena del acueducto vestibular que desemboca en el seno sigmoideo. Durante el recorrido por los conductillos lon- gitudinales del modiolo, las ramas de la arteria coclear describen giros muy pronunciados que pueden tener aspecto de ovillos y se denominan glomérulos arteriales cocleares. Los giros impiden que las pulsaciones se transmitan a la perilinfa y la endolinfa, lo que podría interferir la función receptora de sonidos del órgano de Corti. No se encuentran vasos linfáticos en el oído interno, dado que los espacios perilinfático y endolinfático no se comunican con el sistema de vías linfáticas. anar BOOK'S COPICENTRO USAC / 2016