Scarica LA COMPARTIMENTAZIONE CELLULARE: MATERIALE INTERO ESAME e più Dispense in PDF di Citologia solo su Docsity!
“la compartimentazione cellulare” La differenza fondamentale che caratterizza le cellule eucariotiche da quelle procariotiche è il fatto di avere dei compartimenti interni ; ciò è dovuto a una questione di complessità funzionale della cellula eucariote, che si traduce nella possibilità di far svolgere alla cellula diverse funzioni contemporaneamente , in sedi separate. Per quanto riguarda le membrane , si sa che sono tutte costituite da un doppio strato lipidico in cui sono inserite proteine etc e il motivo sarebbe il fatto che nelle cellule pre-eucariotiche si sia verificata, in relazione al fatto che le membrane hanno forte capacità dinamica, l’introflessione della membrana plasmatica , che si è separata dal resto della membrana, assumendo autonomia e andando a formare dei compartimenti. Il primo che si pensa si sia formato è il RER e tale fenomeno ha avuto dei riflessi sulla formazione di un nucleo isolato, poichè delle particolari membrane simili a quelle del RER hanno circondato i cromosomi , confinandoli in un compartimento che poi sarebbe diventato il nucleo. “reticolo endoplasmatico” IL RE è un compartimento costituito da più compartimenti apparentemente separati fra loro, ma che in realtà sono uniti da un unico lume e che è suddiviso in 2 domini : uno liscio e uno rugoso , per via dei ribosomi adesi alla superficie delle sue cisterne. Mentre il RER è costituito da cisterne appiattite impilate l’una sull’altra, il REL è costituito da tubuli tutti collegati tra loro che sezionati trasversalmente appaiono come dei circoletti. “ RER ” E’ così chiamato per via dei ribosomi adesi sulla sua superficie; essi sono delle strutture sovra molecolari , ovvero costituite dall’unione di più macromolecole: proteine e acidi nucleici (rRNA). I ribosomi sono caratterizzati da una sub unità maggiore e una minore e valgono circa 80s ; questa misura non è riferita soltanto alla massa poiché se prendiamo le due sub unità separate una vale 60s e l’altra 40s, ma anche ad un altro parametro. Infatti, “ s ” sta per “ Unità di sedimentazione ” o “ Unità di Svedberg ” ed è un valore che non si acquisisce solo in riferimento alla massa ma anche della velocità di sedimentazione , cioè quella a cui i ribosomi messi in una centrifuga che li sottopone a una forza che è centinaia di migliaia di volte maggiore della forza di gravità, essi scendono lungo la provetta , quindi sedimentano. Inoltre questo parametro dipende dalla loro massa, forma e dal liquido in cui scendono; da ciò deriva che le due sub unità, essendo una più grande e l’altra meno e con forme diverse, abbiano valori diversi. Sintesi di una parte delle proteine Trasferimento di gruppi glucidici e modificazione chimica delle proteine introducendo dei legami che servono a far acquisire ad ognuna la propria conformazione in modo da renderle funzionali. Smistamento delle proteine “ REL” Spesso quando si parla del REL si fa riferimento a quello degli epatociti , le cellule del fegato , il quale ha tra le principali funzioni quella di detossificare le sostanze che sono nocive per il nostro organismo, immagazzinare glicogeno , nonché contribuire a produrre le sostanze di cui è composta la bile. Ciò è possibile grazie alla presenza di un abbondante reticolo liscio che ha tra le sue funzioni: Degradazione del glicogeno : quando c’è un input ormonale legato un equilibrio tra insulina e glucagone : se c’è troppo glucosio nel sangue, viene sottratto per formare glicogeno o al contrario in mancanza di glucosio viene degradato. Detossificazione cellulare : se sottoponiamo in maniera esagerata il nostro organismo all’assunzione di sostanze tossiche, come l’alcool ad esempio, esso viene compromesso; per questo motivo il fegato lavora per detossificare tali sostanze. Molto spesso queste sostanze sono idrofobiche , perciò per affinità vengono confinate in parti della cellula come le membrane , nei lipidi di accumulo come trigliceridi ed esteri del colesterolo ; a questo punto, poichè il nostro corpo elimina soltanto sostanze idrofiliche, si pensi all’urina o al sudore, gli enzimi del REL catalizzeranno reazioni di aggiunta di gruppi idrofilici , ad esempio
OH , in modo da renderli smaltibili dal nostro organismo. Produzione di componenti lipidici delle lipoproteine : è logico associare al REL questa funzione in quanto della componente proteica se ne occupa il RER che possiede i ribosomi, però siccome è implicato nel metabolismo lipidico, ha questa funzione. Accumulo e rilascio di ioni Ca2+ quando richiesto: ad esempio, durante la contrazione muscolare. “ FASI DELLA SINTESI PROTEICA E SMISTAMENTO DELLE PROTEINE” La differenza fondamentale che caratterizza le cellule eucariotiche da quelle procariotiche è il fatto di avere dei compartimenti interni. Il primo di questi, che si pensa si sia formato come conseguenza di fenomeni di invaginazione della membrana plasmatica, è il reticolo endoplasmatico; esso è suddiviso in domini: uno liscio, dove i ribosomi sono assenti ed è caratterizzato da una morfologia cellulare; uno rugoso, a cui sono adesi i ribosomi. La molecola con il più alto contenuto informazionale è il DNA costituito dai cromosomi; tale informazione viene trascritta sotto forma di RNA , nel nucleo , e tale processo di trascrizione porta alla sintesi di molecole di RNA : transfert messaggero ribosomiale Gli rRNA insieme alle proteine ribosomiali formeranno i ribosomi; tRNA insieme agli amminoacidi formano amminoacil tRNA e con l’ mRNA si arriva alla formazione di polipetid i e ciò avviene nel citoplasma ; infine è previsto l’ assemblaggio della proteina vera e propria. “ TRASCRIZIONE” La fase di trascrizione consiste nel sintetizzare tra di loro nucleotidi e come ogni reazione biochimica a livello cellulare richiede la presenza di enzimi , quali RNA polimerasi. In un dato punto la molecola di DNA si svolge affinchè possano essere lette le sequenze nucleotidiche ; su questa base vengono presi nucleotidi trifosfato che entrano nel nucleo e grazie all’intervento dell’rna polimerasi verrà catalizzata una reazione di polimerizzazione e diversi nucleotidi trufosfato verranno messi insieme e si formerà la molecola di RNA. Negli eucarioti abbiamo 3 tipi di RNA polimerasi I II e III , ognuna delle quali si occupa della polimerizzazione di RNA diversi : mRNA : polimerasi II rRNA : polimerasi I (eccetto l’rna 5s piccolo) tRNA : polimerasi III “ TRADUZIONE” Nel processo di traduzione il linguaggio degli acidi nucleici viene trasformato in quello delle proteine che utilizzano come unità l’amminoacido. Il tRNA è capace di mediare tra questi due linguaggi, in quanto in corrispondenza della coda è presente il sito di legame dell’amminoacido mentre nella porzione di testa vi è l’ansa dell’anticodone , che risulterà complementare alla tripletta di nucleotidi sulla molecola di mRNA. In realtà sono tanti i ribosomi che si associan o alla stessa molecola di mRNA ( polisoma/poliribosoma ), al fine di produrre più catene polipeptidiche uguali, quindi proteine uguali. Quando la catena polipeptidica inizia a prendere contatto con il citosol , già si manifesta un certo ripiegamento degli amminoacidi idrofilici e idrofobici con la controparte citosolica, ma sono necessari dei legami che tengono insieme le varie anse della proteina ; ciò è agevolato dalle proteine Chaperon o chaperonine , presenti nel citosol , ma anche all’interno dei mitocondri. “SMISTAMENTO DELLE PROTEINE” Grazie a degli studi fatti sui ribosomi si è visto che i ribosomi liberi nel citosol sono gli stessi che vi sono adesi al reticolo , quindi gli uni sono in grado di svolgere la funzione degli altri; inoltre si vedeva che la sintesi proteica avveniva nel citosol e sicuramente anche nel reticolo endoplasmatico rugoso. La sintesi di qualsiasi proteina del citoplasma inizia nei ribosomi liberi del citosol e se una proteina è destinata al reticolo rugoso vorrà dire che i ribosomi che hanno iniziato la sintesi proteica nel citosol
Le proteine che lasciano il RER vanno a finire nell’apparato di Golgi. Prima si pensava che esso fosse solo una zona di passaggio per le proteine, invece poi si è scoperto che non è così in quanto svolge diverse funzioni biochimiche che hanno la finalità di modificare le varie glicoproteine. L’apparato di Golgi consiste in un sistema di cisterne separate tra loro per ragioni legate alla presenza di enzimi presenti nelle diverse cisterne, che catalizzano, con esigenze diverse, reazioni per modificare la struttura delle proteine, in modo che sia il più vicino possibile a quella definitiva, quindi aggiunta o rimozione di gruppi glucidici, fosfato ecc e la rimozione di una porzione della catena polipeptidica che rende inattivo l’enzima finchè non è pronto per la sua utilizzazione. Il Golgi inoltre è sede di un traffico bidirezionale , cioè non solo dal RER alla periferia cellulare , ma anche un traffico retrogrado che va dalla periferia cellulare verso il RER. La parte del sistema che è rivolta verso il RER è detta CTN , cioè Cis Golgi Network, e si tratta di una cisterna con diversi fori perché li arrivano tante vescicole in quanto soggetta al traffico, così come quella Trans che invece è sede di uscita delle vescicole ( TGN ) Una vescicola di transizione gemma dagli elementi di transizione del RER e arriva al Golgi, dove si fonde con la zona cis e li riversano parte della loro membrana, in quanto va ad integrarsi con quella del Golgi, ma soprattutto il loro contenuto, che sono glicoproteine. Da qui il contenuto può anche rimanere nel Golgi , se per esempio si tratta di un enzima che opera li, ma la maggior parte sono destinate a lasciarlo. Queste possono quindi essere: vescicole della secrezione attraverso l’ esocitosi , che consiste nella fusione della membrana plasmatica con la vescicola stessa e il rilascio del contenuto all’esterno; vescicole destinate a diventare lisosomi , che si caratterizzano per la presenza di enzimi idrolitici, detti idrolasi , i quali siccome idrolizzano in pH acido, sono dette idrolasi acide Infine vi è la via retrograda che spesso parte da fenomeni di endocitosi , meccanismo per cui si forma una vescicola che gemma verso l’interno della cellula, avvalendosi del contributo della membrana plasmatica, che possono racchiudere dentro di se delle proteine utili. Queste vescicole possono seguire 2 vie:
- una è quella che porta il contenuto della vescicola ai lisosomi , perché hanno capacità degradativa; ad esempio i nutrienti che noi assumiamo devono essere degradati affinchè possano essere assorbiti dal nostro organismo. Il processo avviene in questo modo: RER gemma una vescicola, dentro la quale è contenuta una glicoproteina, un enzima in particolare; Attraversa le diverse cisterne del Golgi , dove subisce trasformazioni , come magari il rimodellamento della componente oligosaccaridica; esso è anche in grado di fosforilare le glicoproteine sia nella componente proteica sia in quella oligosaccaridica , ad esempio viene aggiunto un fosfato al mannosio. Quando arriva alla zona trans avviene il riconoscimento del mannosio 6 fosfato da un recettore presente sottoforma di proteina transmembrana. A questo punto la vescicola lascia il trans Golgi (insieme alle tante altre) e vanno a fondersi con una vescicola più grande in prossimità della zona trans, detta endosoma tardivo , formando il lisosoma maturo e recettore e glicoproteina si staccano in quanto avviene un cambiamento conformazionale da parte del recettore a causa del pH basso dell’endosoma tardivo.
- esistono anche delle vescicole che gemmano dalla zona Cis del Golgi per andare a fondersi con il RER. Quando abbiamo una proteina che deve stare nel RER perché li deve svolgere delle funzioni, però transitoriamente lo lascia , va a finire nell’apparato di Golgi nelle zone Cis, dopo di che il recettore di cui questa proteina è fornita, che ha lasciato anch’esso la membrana, riconosce la proteina a cui era agganciato e la riporta indietro ; la sequenza che permette di riconoscere la proteina è detta KDEL. recettore e glicoproteina si staccano in quanto avvieme un cambiamento conformazionale da parte del recettorea causa del pH basso dell’endosoma tardivo. “FORMAZIONE DELL’ENDOSOMA TARDIVO” Dopo che l’idrolasi si è unita all’endosoma, da esso si stacca una vescicola che torna indietro e va a fondersi con la faccia trans, perché dopo che il recettore si è staccato dall’idrolasi, in quato il pH si è abbassato, viene recuperato il recettore per essere riutilizzato. Si pensa che l’endosoma tardivo origini da invaginazioni della membrana plasmatica che formano vescicole le quali si uniscono e danno origine all’endosoma precoce. Esso all’interno ha un pH acido che è dovuto alle pompe V , presenti su una vescicola originata dal trans, che prendono H+ dal citosol e li mandano dentro l’endosoma precoce grazie alla loro fusione e formando l’endosoma tardivo.
“LISOSOMI”
Sono frutto della fusione di una vescicola idrolasica e un endosoma tardivo. Al suo interno un lisosoma ha: pompe V H+ ATPasiche che lavorano idrolizzando ATP e rendono il pH del fluido interno al lisosoma acido idrolasi acide , cioè enzimi idrolitici che catalizzano a pH acido reazioni di degradazione di tutte le macromolecole biologiche. Nel versante interno della membrana del lisosoma è presente uno strato saccaridico analogo a quello del glicocalice per evitare che gli enzimi si possano avvicinare e degradare la membrana lisosomiale. “ESOCITOSI” L’esocitosi è il fenomeno di membrana alla base del processo secretorio il quale porta all’espulsione di sostanze al di fuori della cellula e all’integrazione delle proteine della vescicola con la membrana plasmatica. Le vescicole secretorie possono andare incontro a due comportamenti: una secrezione costitutiva , la cellula la mette in atto continuamente indipendentemente dalla presenza di segnali una secrezione regolata , in cui le vescicole di secrezione che gemmano dalla zona trans del Golgi non si fondono subito con la membrana plasmatica, ma tendono ad accumularsi in prossimità spesso fondendosi e formando vescicole più grandi, chiamati granuli di secrezione , dopo di che queste sono destinate a fondersi con la membrana solo dopo l’arrivo di un qualche segnale alla cellula “ENDOCITOSI” Nell’endocitosi, materiale che sta esterno alla cellula vieni portato dentro , si forma la vescicola che si stacca e porterà il materiale o ai lisosomi o al Golgi. In questo caso ciò che stava sulla membrana plasmatica , come lipidi e proteine, si staccano e impoveriscono la membrana plasmatica quando si forma la vescicola. Fagocitosi : durante la quale vengono fagocitate cellule e batteri, ma anche aggregati macro molecolari. Dopo la loro ingestione, molto probabilmente andrà ai lisosomi per essere degradato Pinocitosi vorrebbe dire cellula che beve, ma ad oggi sappiamo che l’acqua può entrare nella cellula con un fenomeno abbastanza semplice che è l’osmosi, senza determinare un’invaginazione. Il vero significato si pensa sia legato ad un equilibrio della membrana plasmatica , perchè quando la cellula fa esocitosi la membrana di ogni vescicola va ad aggiungersi alla membrana plasmatica che diventerà sempre più grande e studi hanno dimostrato che una cellula deve mantenere determinati rapporti tra superficie e volume. Quindi, siccome questi fenomeni avvengono soprattutto nelle cellule che secernono , come quella del pancreas esocrino, più secerne più aumenterà la superficie. La pinocitosi serve proprio ad equilibrare questo problema , perché una endocitosi di qualsiasi tipo equilibra il fenomeno nel modo opposto, però se ad una vescicola di esocitosi , che porta membrana, si alterna una di endocitosi che la sottrae, il fenomeno si equilibra. Tra l’altro è stato dimostrato che la vescicola di endocitosi che si forma sottrae componenti vecchi della membrana. Siccome la pinocitosi porta all’interno della cellula solo materiale liquido, ci si è resi conto che la esercitano cellule che fanno una intensa secrezione , la membrana delle quali diventerebbe sproporzionata senza questo fenomeno. Endocitosi mediata da recettori clatrino-dipendente : i recettori riconoscono materiale di interesse per la cellula, come avviene nella fagocitosi, in questo caso però la particolarità è data da proteine che si uniscono in coincidenza con l’invaginazione della membrana plasmatica , cioè in corrispondenza della fossetta sono presenti delle proteine periferiche (adaptine) che si associano ai recettori e consentono il legame dei recettori con la clatrina. Le molecole di clatrina hanno una conformazione particolare: più catene polipeptidiche che si uniscono a formare una struttura molto geometrica con angoli di 120° e formano le triskelion , i quali quando si mettono insieme formano diverse molecole di clatrina e viene a formarsi una
La presenza di due membrane è legata all’evoluzione cellulare che ha visto un passaggio dalla condizione di cellula procariotica a pre-eucariotica fino ad eucariotica, con acquisizione di una compartimentazione interna; parallelamente le cell. pre-eucariotiche , che erano tendenzialmente più grandi , attraverso un’invaginazione si sono portate dentro una cellula procariote e ciò sta alla base della “ Teoria endo- simbiontica”. Questa teoria spiega che questi processi sono avvenuti in relazione a ciò che stava accadendo nell’ambiente, cioè l’ atmosfera stava progressivamente arricchendosi di O2 e la cellula procariote aveva sviluppato un metabolismo adatto all’ossigeno , mentre quella pre-eucariotica era più grande e aveva sviluppato delle compartimentazioni interne; perciò è avvenuta l’endocitosi della cellula procariote a vantaggio della cellula pre-eucariote, ma anche per l’altra cellula che può stare in un ambiente chimicamente definito , usufruire della capacità sintetica della cell pre-eucariote. I mitocondri hanno una capacità di moltiplicazione che è simile a quella dei procarioti , che si dividono per scissione: duplicano prima il DNA e poi vanno incontro a scissione , non solo sono anche in grado di sintetizzare molte proteine. Essi hanno funzioni, quali: Metabolismo energetico Seconda sede di accumulo degli ioni Ca Coinvolti nel metabolismo degli acidi grassi (non superiori a 16C) (in collaborazione con i perossisomi) “Metabolismo energetico” I mitocondri sono impegnati nel metabolismo ossidativo per cui producono prodotti di rifiuto, ovvero i radicali liberi, dannosi per la cellula. Per la motilità, la biosintesi di sostanza cellulari, trasporto attivo e la trasmissione degli impulsi è necessaria energia sotto forma di ATP, prodotto soprattutto dai mitocondri e dalla glicolisi anaerobia. Per produrlo hanno bisogno di combustibili, quindi carboidrati e acidi grassi + O2+ADP+PiCO2+H2O. “Anaerobiosi” Ricavare ATP senza la presenza di O2: Glicolisi. Consiste nel rompere i legami per ricavare gli elettroni (H+) dall’acido piruvato, ottenendo acido piruvico; gli H+ vengono acquisiti da NAD+ e FAD che si riducono in NADH e FADH2. “Aerobiosi”
- Respirazione cellulare Entrano piruvato e acidi grassi e vanno nella matrice mitocondriale Dall’ossidazione del piruvato e degli acidi grassi si ricava l’Acetil-CoA (3C) Esso entra nel Ciclo di Krebs, il cui fine è quello di ricavare e- e prodotti di scarto, cioè CO Gli e- vanno NAD+ e FAD che diventano NADH e FADH Successivamente il processo si trasferisce nelle creste mitocondriali (membrana interna), dove NADH e FADH2 si ossidano per darli a complessi proteici accettori di e- e saranno pronti a riiniziare il ciclo. “Membrana esterna” La membrana esterna è abbastanza permeabile perché sono presenti porine (proteine) che fanno passare componenti idrofili. “Membrana interna” Nella membrana interna abbiamo gli eccettori degli e- che vengono forniti da NADH e FADH2. L’elettrone viene acquisito dal primo complesso respiratorio, che è un insieme di proteine, passandolo agli altri, creando una catena di ossido-riduzione, che è un flusso di elettroni, cioè energia. Questo meccanismo è detto catena di trasporto degli elettroni/catena respiratoria. Dopo i complessi respiratori troviamo i complessi Fo (oligomicina, che è un antibiotica)-F1. Ognuno è scomponibile in 2 parti: Proteina canale che fa passare lo ione H+ Parte ATPasica che ricava ATP da ADP+Pi “FOSFORILAZIONE OSSIDATIVA” Gli e- del ciclo di Krebs vanno a FAD e NAD+ che diventano NADH e FADH2.
Gli e- vengono ceduti sulla membrana interna alla catena di trasporto degli elettroni per prenderli alla fine l’O2, accumulandoli nello spazio intermembrana, ottenendo un gradiente ionico, cioè energia disponibile, che prende il nome di Potenziale intermembrana dei mitocondri . I complessi Fo-F1 funzionano come pompe V che funzionano al contrario in quanto, sfruttando il gradiente ionico, riportano gli ioni H+ secondo gradiente nella matrice, liberando energia, cioè ATP. “MAP2” Le MAP2 contribuiscono a stabilizzare la struttura del microtubulo, tenendo insieme i singoli eterodimeri, mettendosi parallele al microtubulo e alla fine formano una specie di piega, che serve a far prendere contatto i microtubuli con altri microtubuli o altri componenti del citoscheletro. “MAPs MOTRICI” Sono proteine motrici che hanno un ruolo nella motilità cellulare, cioè i movimenti che avvengono all’interno delle cellule, i quali coinvolgono i microtubuli, microvescicole e vescicole. Queste proteine hanno 2 teste che usano per camminare sul microtubulo e appartengono a 2 famiglie principali: Dineine si muovono verso l’estremità – Chinesine si muovono vero l’estremità + I microtubuli sono rivolti con il – verso il centrosoma e il + verso la membrana plasmatica. Il movimento è possibile grazie alla loro capacità ATPasica. Tali proteine sono formate da catene polipeptidiche che si avvolgono in maniera più globulare in corrispondenza delle teste e un sistema di intermediazione utile, dove è presente un dominio di carico, per far agganciare la proteina motrice alla membrana cargo. Gli studi hanno dimostrato che le vescicole, sia del traffico anterogrado sia retrogrado, si muovono grazie alla chinesine e alle dineine che camminano sui microtubuli; inoltre, si sa che l’apparato di Golgi tende a localizzarsi in corrispondenza del centrosoma. “MECCANISMO DI TRASPORTO ASSONICO” I neuroni sono state le cellule più studiate per avere tutte le informazioni che ad oggi abbiamo sul citoscheletro, in quanto deve sostenere i vari dendriti e l’assone. Il traffico vescicolare è ancora più importante in un assone, che permette la comunicazione fra neuroni e fra neurone ed altre cellule, poiché le vescicole trasportano i neurotrasmettitori fino alla terminazione sinaptica; in particolare il traffico delle vescicole sinaptiche è possibile grazie alle chinesine che si muovono verso la periferia. “MICROTUBULI ASSONEMALI” Questa tipologia non è presente in tutte le cellule, ma solo quelle che hanno l’assonema, cioè quelle che possiedono ciglia o flagelli. Esse si trovano nell’apparato respiratorio, genitale femminile e negli spermatozoi. Se seziono un ciglio o un flagello vedo la stessa cosa, cioè un assonema che è un insieme di microtubuli, detto Sistema 9+2, ovvero 9 coppie+1coppia centrale. I microtubuli sono di 2 tipologie: A: che hanno parete completa, formata da 13 protofilamenti B: addossa i suoi protofilamenti al microtubulo A e ha una parete incompleta, formata da 9/ filamenti. I microtubuli centrali hanno entrambi la parete completa. La coppia centrale è tenuta insieme da una guaina proteica; un’altra proteina dispone le coppie periferiche secondo una disposizione circolare e si lega soprattutto al tubulo A formando ponti di Nexina. Altre proteine partono dal microtubulo A fino alla guaina per formare raggi e sono presenti anche bracci di dineina che vanno dal microtubulo A al B della coppia vicina. La dineina flagellare ha 3 teste con capacità ATPasica, le quali sono capaci di caricare un microtubulo e camminare su un altro; tale fenomeno è detto “Sliding del microtubulo”. Attraverso degli esperimenti, si è visto che le coppie periferiche scorrevano l’una sull’altra, ma non si muovevano e alla fine si è scoperto che il movimento del ciglio è il frutto di una risultante di forze contrarie, che non permette lo scorrimento ma che porta ad un piegamento.
Protrusioni : es. microvilli, filopodio se molto lunghi, sostenuti da un fascetto di filamenti di actina; Estrusioni a foglietto : lamellipodi Invaginazioni : durante i fenomeni di endocitosi o nella citodieresi. “STRUTTURA DEI MICROFILAMENTI ” I microfilamenti del citoscheletro sono costituiti da actina G , cioè globulare che polimerizza e si dispone come se ogni microfilamento fosse formato da 2 catene e presenta un’incavatura che lega non il GTP, ma l’ ATP. Anche nei microfilamenti è stato dimostrato il treadmilling, in base alla quantità di actina libera nel citosol e si pensa serva per il rinnovamento e per il trasporto di vescicole nel mondo vegetale. Anche nei microfilamenti di actina è presente una certa polari tà, abbiamo infatti una Pointed end che depolimerizza più facilmente e una Barbed end che polimerizza più facilmente e velocemente ; è possibile distinguerle tagliando l’actina con la miosina, la quale si lega sempre alla Pointed end. Abbiamo una < polimerizzazione di Actina G a formare Actina F dove il citoplasma è più denso , come nella corteccia cellulare. La cellula può avere diverse ragioni per avere un citoplasma più o meno denso, ad esempio nel caso della diapedesi in cui il citoscheletro viene disorganizzato per permettere alla cellula di extravasare; la proteina che lo permette è la Gelsolina , che è Ca-dipendente , il quale quando viene rilasciato provoca la sua attivazione, cioè taglia i microfilamenti ( Severing protein ). Inoltre avere un citoscheletro più o meno denso più aiutare la cellula a sopportare stress meccanici. “ABP” Così come le MAPs dei microtubuli, le ABP sono proteine che legano l’actina : Miosina I : serve ad agganciare un microfilamento ad una membrana biologica , quindi non solo a quella citoplasmatica; Miosina II o sarcomerica: funziona come le dinenine e le chinesine che possono anche far scorrere 2 microfilamenti tra di loro; Troponina “ORGANIZZAZIONE DEI FILAMENTI” A fasci , come nei sarcomeri e possono essere più o meno distanti:
- + distanti : - Tinina , affinchè si possano interporre i filamenti di Miosina II;
- distanti : Villina (è stata scoperta nei microvilli, i quali hanno capacità contrattile che facilita l’assorbimento, quindi actina e miosina oltre a tenerlo dritto, lo fanno anche contrarre) o Fimbrina ; In reti , come nella corteccia cellulare, che è molto solida e resistente grazie all’actina e per dare al citoplasma una capacità contrattile, utile nel movimento. “STEADY-STATE E TRADMILLING” La quantità di monomero non polimerizzato che è in equilibrio con il polimero è definito “concentrazione critica” e rappresenta la minima concentrazione di proteina necessaria per la polimerizzazione. Allo Steady-state le concentrazioni sia del polimero sia del monomero non cambiano , ma ciò non significa che il sistema sia fermo; infatti, una certa quantità di monomeri continua ad essere polimerizzata a livello della barbed end, mentre la pointed end rilascia monomeri e il flusso è unidirezionale B P. Questo fenomeno è detto Treadmilling. “AVANZAMENTO DI UNA CELLULA SU UN SUBSTRATO SOLIDO ” Una cellula che vuole muoversi manda avanti una protrusione , sfruttando la capacità dei microfilamenti di polimerizzare tutti insieme , spostando la membrana. Questo lamellipodio non ha citosol , che viene spinto dalla cellula all’interno di esso; a questo segue la cellula che stacca i contatti focali che la tenevano ancorata al substrato. “FILAMENTI INTERMEDI” I filamenti intermedi sono strutture piene che tendono a concentrarsi intorno al nucleo , al fine di metterlo nella posizione giusta e ancorarlo. Essi vengono considerati come i connettori del citoscheletro e anche in questo caso sono presenti Proteine Associate ai Filamenti Intermedi ( IFAP ).
Possono essere di 2 tipologie : Citoplasmatici , nei quali il costituente proteico dipende dal tessuto istologico da cui deriva:
- Cheratine , cell. epiteliali
- Vimentina e Vimentino-simili , cell. Del connettivo, muscolari ( Desmina ) e della neuroglia; Nucleari :
- Lamine nucleari , in tutte le cellule nucleate. I filamenti sono come una fune, cioè sono costituiti da unità più piccole , cioè proteine filamentose , che aiutano a distribuire le forze. Fil+Fil= Dimero Dim+Dim= Protofilament o Protofilamenti in più piani= Filamento intermedio Per questo motivo è difficile la loro depolimerizzazione , che viene fatta con la fosforilazione. “IL NUCLEO CELLULARE” La struttura del nucleo è mantenuta da 2 componenti fondamentali : la lamina nucleare , che è una maglia costituita da filamenti intermedi ( lamìne ), che sostiene l’involucro nucleare ; essa si continua
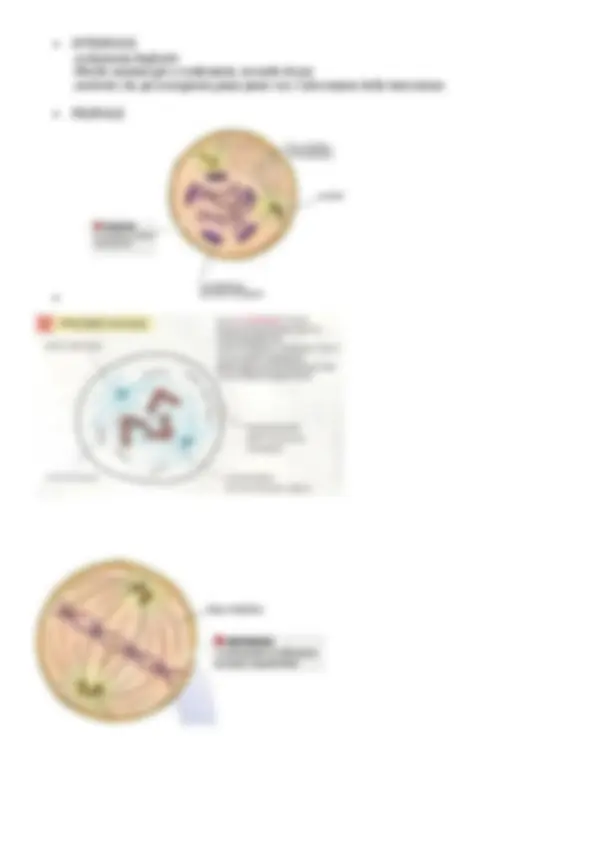
In nero : l’eterocromatina, con DNA e proteine In bianco : l’ eucromatina , “ “ “ Grigio chiaro : il nucleolo , con RNA e proteine. L’eterocromatina è solitamente associata all’involucro nucleare oppure al nucleolo e prendono il nome di “ eterocr.costitutiva ”; essa è inattiva trascrizionalmente e ha funzione strutturale ; mentre l’eterocromatica “ facoltativa” che è trascrizionalmente inattiva, ma se richiesto può decompattarsi e diventare attiva. L’ eucromatina è definita come attiva trascrizionalmente. “Composizione chimica della cromatina” Se esamino una singola fibrilla , è costituita da DNA associato a proteine , dette istoni , e altre proteine che non sono associate dette “ non istoniche ”. “ISTONI” Le proteine istoniche sono caratterizzate da catene polipeptidiche con amminoacidi prevalentemente basici perché nel DNA i gruppi fosfato avranno perso gli ioni H+ avranno carica negativa. Gli istoni principali sono H1, H2A H2B H3 H “NON ISTONICHE” Hanno una minore interazione con il DNA poiché possiedono cariche negative oppure sono neutre. Esse servono a far funzionare il Dna, quindi ad attivare la lettura di certi geni piuttosto che altri e in grado di catalizzare la trascrizione e la duplicazione come la DNA-polimerasi. “GLI ORDINI DI COMPATTAMENTO DELLA CROMATINA” Collana di perle ( 10nm ): è costituita da nucleosomi caratterizzati da 8 proteine istoniche tranne H1 , che formano un ottamero istonico o core del nucleosoma , il quale è avvolto dal DNA ( coppie di nucleotidi) e ognuno di essi è unito da DNA linker ; Fibra formata da nucleosomi compattati/solenoide ( 30 nm): da questo livello in poi interviene l’istone H 1, che servirebbe a tenere bloccate le spire del solenoide , formando una sorta di guaina. Domini ad ansa ( 300 nm) Eterocromatina ( 700 nm) Cromosoma mitotico ( 1400 nm) Ciò è stato visto che è possibile perché c’è un’impalcatura di natura proteica che gli dà la forma , sulla quale si ancorano le anse , la quale è costituita da proteine non istoniche e che prende il nome di “scaffold”. Esso nell’interfase corrisponde al nucleoscheletro. “IL NUCLEOLO” Il nucleolo è costituito da proteine e RNA ribosomiale. Esso è molto importante perché la sua presenza risulta associata ad una cellula che ha un’alta attività trascrizionale , in quanto essendoci tanto RNA ribosomiale significa vi è un’alta attività trascrizionale e il nucleo sarà eucromatico. L’elevata sintesi proteica si può dedurre quindi da:
Nucleolo evidente Nucleo eucromatico Citoplasma ricco di ribosomi Abbondante RER e Golgi. Questo organulo non è facilmente individuabile poiché non è definito da membrana , ma appare come una nuvola molto densa formata da unità granulari ; in esso è riconoscibile una: Porzione interna : pars fibrillaris Porzione esterna : pars granularis. Le granulazioni sono le subunità dei ribosomi appena assemblate e che stanno per lasciare il nucleolo attraverso i pori nucleari; esso è sede della trascrizione di tutti gli rRNA , fatta eccezione per l’RNA 5s , il quale viene prodotto in sede extra-nucleolare, e che in particolare avviene nella parte fibrillare grazie alla presenza di geni per gli RNA , provenienti da 10 cromosomi interfasici ; tali porzioni di cromosomi sono detti NOR-Organizzatori nucleolari. Nella cellula possono esserci più nucleoli o solo uno a secondo della distanza dal ciclo cellulare , in particolare la mitosi. Ad esempio, nel neurone ne vedo sempre 1 perché non va mai in mitosi ; nelle cellule che si dividono , può capitare che i cromosomi che contengono gli organizz. nucleolari siano lontani , quindi in corrispondenza di essi si formano piccoli nucleoli che poi si uniranno in un unico nucleolo. “DIVISIONE DELLE CELLULE” Lo zigote è la prima cellula che si forma dall’unione di 2 gameti , che ha da subito l’ obiettivo di diventare un organismo pluricellulare. In un organismo pluricellulare adulto , la proliferazione è finalizzata al mantenimento dell’organizzazione dei tessuti ; mentre l’organismo unicellulare prolifera al fine di generare cellule figlie. Tuttavia non tutte le cellule hanno capacità di divisione , come i neuroni o le cellule muscolari cardiache ; infatti esse si distinguono in: Cellule perenni : non si dividono mai, come i neuroni; Cellule stabili : si rinnovano una volta all’anno , come gli epatociti; Cellule labili : si rinnovano una volta al giorno , come gli enterociti dell’epitelio intestinale, i precursori dei globuli bianchi e rossi. Tale classificazione è stata fatta da uno scienziato italiano, Bizzozzero. Alla base di ogni divisione cellulare vi è sempre la duplicazione del DNA , in quanto le cellule figlie devono avere lo stesso corredo cromosomico della cellula madre, e che prende il nome di “ Sintesi ”. La fase M , corrisponde a Mitosi (divisione del nucleo) e citocinesi (divisione del citoplasma), ma non è detto che avvengano entrambe. Il periodo che sta tra la fase M e la S e tra la S e la M è la fase G1 e G2 , detta intervallo : la G1 è una fase di accrescimento durante la quale la cellula inizia ad esplicare la propria attività funzionale , mentre la G2 è un periodo di preparazione alla divisione. Le fasi S, M e G 2 hanno una durata che sta nell’ordine delle ore , quindi la fase varia è la G1 , a seconda della durata del ciclo cellulare. “MODELLO SEMICONSERVATIVO DELLA REPLICAZIONE DEL DNA” Il DNA è un acido nucleico a due filamenti , i quali si avvolgono a doppia elica e sono uniti da legami idrogeno. La duplicazione del DNA avviene tramite il modello semiconservativo che prevede che la doppia elica si apra e che vengano sintetizzati , sulla base di un principio di complementarietà tra le basi , 2 nuovi filamenti di DNA. “IL CICLO CELLULARE” Il controllo sul ciclo cellulare è un fattore importantissimo, considerando che le cellule neoplastiche si caratterizzano per avere dei parametri che saltano, tra i quali una capacità proliferativa incontrollata, e alcuni problemi legati a questo aspetto risiedono in un mancato controllo del ciclo cellulare.
La cellula prima di entrare in mitosi inizia a produrre tante cicline che troveranno pronte le chinasi, si uniranno e diventeranno un complesso Cdk attivo pronto a fosforilare altre proteine. Queste altre proteine dipendono dalla fase del ciclo in cui mi trovo, cioè complessi Cdk diversi per ogni fase perché altrimenti fosforilerebbero tutte le stesse proteine. Nella fase G1 abbiamo una chinasi arancione e una ciclina in azzurro e succede che la cellula da l’avvio al passaggio alla fase S successiva nel punto di controllo solo se riesce a formare un complesso CdK solo se c’è abbastanza ciclina. Il complesso Cdk attivo può fosforilre nella fase S proteine come DNA polimerasi; una volta completata la fase S la ciclina di questa fase viene degradata e la Chinasi non può più essere attivata e la cellula va avanti nel suo ciclo. Il Cdk per la mitosi sarà formato da un’altra chinasi e un’altra ciclina e fosforilerà in questo caso proteine come quelle che hanno a che fare con la compattazione dei cromosomi, quindi l’istone H1, in modo che i cromosomi passino dalla forma interfasica a quella mitotica; poi proteine (lamine) dell’involucro nucleare affinchè i cromosomi possano prendere contatto con i microtubuli del fuso mitotico.
“LA MITOSI”
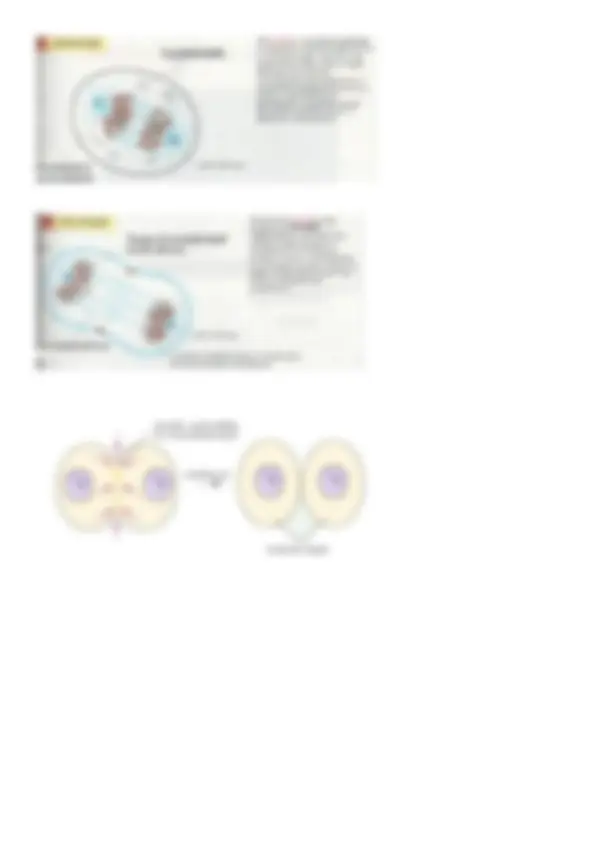
“Ciclo dei centrioli” Il centrosoma è il centro di organizzazione dei microtubuli che da qui si dipartono in tutta la cellula e al cui centro vi sono i centrioli. Il fuso mitotico è una struttura simmetrica che posso dividere in 2 parti perché è costituito da 2 centrosomi ognuno andrà a una delle cellule figlie. La cellula acquisisce i 2 centrosomi nell’ultima parte della fase S, in particolare i centrioli, che prendono il materiale pericentriolare dal citosol per formarlo. G1: I 2 centrioli da cui partono i microtubuli S: I centrioli vengono duplicati e il materiale pericentriolare li circonda tutti e 4 M: - profase: i 2 centrosomi iniziano a distanziarsi l’uno dall’altro e avremo i due poli del fuso e i 2 potenziali centrosomi che andranno nelle cellule figlie; “Mitosi”