Scarica Microbiologia: Fondamenti e Applicazioni - Prof. Turin e più Appunti in PDF di Microbiologia solo su Docsity!
Microorganismi
- esseri viventi invisibili a occhio nudo
- esistono come singole cellule oppure piccoli aggregaI cellulari, oppure non hanno nemmeno la struttura cellulare (virus)
- Esempi: Adenovirus: forma regolare Coronavirus: forma irregolare (ha un envelope) ➔ Le cellule microbiche sono diverse dalle cellule eucariote. È in grado di svolgere una vita autonoma / indipendente, svolgere tutte le funzioni vitali. La cellula
- Unità fondamentale di tutta la materia vivente (eccetto i virus a struttura acellulare)
- Entità isolata dall’ambiente esterno (membrana e parete)
- ConIene una varietà di sostanze, organuli e strutture cellulari
- Tutta la materia vivente è fatta da 4 componenti (ecc. i virus che a volte hanno solo alcuni dei seguenti componenti):
- Proteine
- Acidi nucleici
- Lipidi
- Polisaccaridi 6 gruppi principali di microorganismi :
- Batteri (struttura cellulare, unicellulari)
- Protozoi (struttura cellulare, unicellulari)
- Funghi (struttura cellulare, pluricellulari o unicellulari)
- Alghe (struttura cellulare, pluricellulari)
- Cianobatteri (struttura cellulare, unicellulari)
- Virus (acellulari) Procarioti vs Eucarioti Dai procarioti agli eucarioti: Tutti gli organismi presenti ad oggi sulla Terra sono derivati da una cellula procariota primordiale (3,5 miliardi di anni fa) che si nutriva di sostanze organiche. Per circa 1,5 miliardi di anni erano presenti solamente procarioti sulla Terra, fino a quando non è iniziata l’evoluzione a causa della mancanza di sostanze organiche. Sono comparse cellule eucariote che erano in grado di sintetizzare le sostanze organiche. Così la Terra si è popolata prima di piante poi di animali. Si sono sempre adattati ai cambiamenti delle condizioni ambientali. Nonostante ciò, ancora oggi i procarioti sono le forme dominanti nella biosfera! (La loro massa biologica
supera almeno di 10 volte quella di tutti gli eucarioti messi insieme, quindi sono preponderanti nella biosfera.). Ad oggi non scoperti archeobatteri patogeni. Mesosomi: Sono dei pseudoorganelli presenti nei procarioti (invaginazioni della membrana plasmatica dei batteri), supportano delle strutture ad esempio i ribosomi. Nel citoplasma dei procarioti sono presenti i ribosomi. Il citoplasma delle cellule eucariote conIene gli organelli, separati dalla membrana:
- Reticolo endoplasmatico
- Complesso del Golgi
- Lisosomi / perossissomi
- Citoscheletro
- Mitocondri (in cellule vegetali oltre ai mitocondri ci sono i cloroplasti)
- Vacuoli (cellule vegetali) **Caratteristiche del materiale ereditario comune a procarioti ed eucarioti = molto importante
- Replicazione + Trasmissione alla progenie dell’informazione genetica
- Trascrizione e traduzione dell’informazione genetica —>** il materiale genetico è come un libro chiuso, per svolgere le funzioni servono le proteine che trascrivono (in RNA messaggero) e traducono le informazioni
- Mutazione e ricombinazione per introdurre variabilità genetica (+ Selezione Naturale) Questi tre step vengono chiamati “Dogma Centrale della Biologia” Replicazione: DNA—>DNA Trascrizione: DNA—>RNA Traduzione: mRNA—>proteine La sintesi proteica È una delle funzioni primarie della cellula, perché solo grazie alle proteine può funzionare tutto. L’informazione genetica necessaria per sintetizzare le proteine è costituita da DNA, contenuta nei geni (unità ereditarie della cellula). La sintesi proteica è importantissima. La sintesi proteica ha luogo sui ribosomi, a livello dei ribosomi l’RNA messaggero serve per sintetizzare le proteine (catena di amminoacidi). Reticolo endoplasmatico RER: ribosomi sulla membrana, sintetizza le proteine
Le cellule animali sono connesse tramite 3 tipi di giunzioni:
- Giunzione di ancoraggio (desmosomi)
- Giunzioni occludenti (serrate)
- Giunzioni comunicanti (gap) D’ora in poi la cellula eucariote non viene più trattata nel corso, solamente per fare paragoni durante il ciclo riproduttivo dei virus. I microorganismi impattano sulla nostra vita sia in senso positivo che in senso negativo. Esempi: Non dobbiamo imparare tutti i nomi delle malattie a memoria, molti li conosciamo già
Distribuzione dei microorganismi in natura: Sono presenti dappertutto:
- Suolo
- Acqua
- Aria
- Cibi
- Materiali e utensili
- Organismi viventi (piante e animali) Gli organismi patogeni preferiscono vivere sull’ospite. In generale i microorganismi sono più abbondanti dove trovano nutrimento / umidità adatti ad accrescimento e moltiplicazione. Tipologie:
- Mutualismo (microorganismi uIli, ci proteggono perché competono con nuovi batteri potenzialmente patogeni ed aiutano ad eliminarli). Le due specie traggono beneficio reciproco, l’animale fornisce sostanze nutritizie e fattori di crescita per il microorganismo, condizioni di pH, T, pressione osmotica costanti, i batteri aiutano la digestione degli alimenti.
- Commensali : non danneggiano ne beneficiano l’ospite
- Parassiti : sono patogeni Il termine “simbiosi” è più generale e include sia mutualismo sia commensalismo sia parassiIsmo. Può essere esocellulare (superfici cutanee, mucose, cavità) o intracellulare/endosimbiosi (i batteri che colonizzano gli organismi unicellulari come i protozoi). Il simbionte solitamente produce metaboliti o tossine importanti per l’ospite. Ruolo dei microorganismi:
- Ruolo nocivo dei microorganismi
- Agenti di malattie
- Contaminanti alimentari (lungo periodo di contaminazione)
- Ruolo benefico dei microorganismi
- Decompositori
- Fissatori di Azoto (es. rhibozium)
- Celluloso – Digestori (microorganismi del rumine)
- Alimentari (lieviI e muffe)
- Produttori di Farmaci (addiIvi)
- Decontaminatori e produttori di energia (degradano prodotti di rifiuto)
- Ingegneria genetica e applicazioni biotecnologiche (Es.: DNA ricombinante) Morfologia, struttura e funzione della cellula batterica La morfologia della cellula batterica si riferisce, oltre alla forma e alle dimensioni della singola cellula, anche al modo in cui le cellule si dispongono tra loro. La maggior parte dei batteri presenta forma ben definita, a cocco (sferico), a bacillo (allungato) o a spirillo (a spirale). Alcuni microrganismi sono invece filamentosi (più sottili, sembrano fili che si attorcigliano); altri invece sono pleomorfi , cioè possono assumere diverse forme, non hanno una forma propria. Siccome i batteri allungati si dividono sul piano trasversale, si vedono ancora parzialmente legati tra loro tramite un polo. Cocchi Sono microrganismi sferici di dimensione 0,5- 1 mm (diametro); si osservano spesso a gruppi a causa dell’incompleta separazione delle cellule singole durante la riproduzione. La divisione, sempre secondo lo stesso piano,
Nei patogeni la capsula serve a:
- Favorire l’adesione e la colonizzazione a livello , ad esempio, delle mucose (dà il via al processo infettivo)
- Proteggere il batterio dagli agenti antibatterici naturali (batteriofagi, colicine, complemento —> costituito da noi, è un meccanismo di difesa contro i batteri soprattutto capsulati, lisozima —> enzima che viene secreto e contenuto nelle lacrime ed è antibatterico)
- Proteggere il batterio dall’ingestione e distruzione da parte delle cellule fagocitarie dell’ospite I batteri legati con la capsula sono patogeni, gli altri no. L’unica differenza tra ceppi patogeni e ceppi innocui (es Streptococcus pneumoniae ) è l’assenza o la presenza della capsula. Esempi di noti patogeni che possiedono la capsula sono il Clostridium perfrigens (causa gangrena gassosa), il Bacillus anthracis (causa del carbonchio ematico), la Klebsiella pneumoniae (agente della polmonite), Haemophilus influentiae (che può causare la meningite). Anche lo sviluppo della carie dentaria ad opera di Streptococcus mutans dipende dalla sua adesività: aderiscono ai denti e trattengono altri batteri che digeriscono gli zuccheri che mangiamo aiutano a far degenerare lo smalto dei denti generando in seguito le carie. Questo microrganismo normale residente del cavo orale metabolizzando il saccarosio produce il glicocalice , che ingloba e fissa ai denti altri batteri, i quali producono metaboliti responsabili della carie. La composizione chimica della capsula dipende dalla specie: nella maggior parte è di natura polisaccaridico o polisaccaridico-proteica; in alcuni casi (es nel Bacillus anthracis) è di natura polipeptidica, in particolare costituita da un polimero dell’acido D-glutammico, che tra l’altro è responsabile della metacromasia del Bacillus anthracis al blu di Loffer (colora bene il batterio perché è dotato di capsula polipeptidica). Parete dei batteri È la struttura che circonda la membrana citoplasmatica e che conferisce ai batteri la forma (infatti se si rimuove il batterio assume forma sferica detto protoplasto o sferoplasto). Ha funzioni di:
- Protezione fisica della cellula (nei confronti dell’osmosi): protegge dalla lisi osmotica, cioè la morte per rottura (osmosi fenomeno per cui l’acqua passa per la cellula). La lisi osmotica è quando l’acqua entra nella cellula con dei soluti in modo da portare la cellula ad avere troppi richiami d’acqua causando lo scoppio della cellula
- Conferisce rigidità alla cellula Particolarità:
- È costituita da sostanze presenti solo nel mondo microbico
- In alcuni batteri è responsabile della patogenicità
- È il bersaglio di alcuni antibiotici
- La sua diversa composizione è responsabile del comportamento alla colorazione di Gram : nell’800 si riuscì a distinguere i batteri in due grandi categorie in base alla composizione della parete, gram positivi e gram negativi È costituita dal ripetersi di molecole di natura polisaccaridica-amminoacidica dette peptidoglicano , che è costituito da due amminozuccheri e una catena di quattro o cinque amminoacidi. Gli amminozuccheri sono l’acido N-acetilmuramico e l’N-acetil-glucosamina, che si alternano a formare un polimero ad elevato peso molecolare, consentendo anche l’aggancio delle catene peptidiche laterali di quattro o cinque amminoacidi a formare il muramilpeptide. Nei gram positivi poi le catene peptidiche sono ulteriormente legate dell’interpolazione di altre catene peptidiche (5-glicine): transpeptidazione a costituire il peptidoglicano. I diversi batteri hanno strati più o meno numerosi sovrapposti di peptidoglicano, legati tra loro in diverso modo a seconda che si tratti di gram positivi o negativi. In particolare, in quelli positivi i legami sono più forti e numerosi. Gram-positivi Più strati di peptidoglicano: hanno parete costituita da circa 20 strati sovrapposti di peptidoglicano tenuti insieme da ponti amminoacidici, si tratta in sostanza di un intreccio di fibrille. Il peptidoglicano costituisce circa il 95% della parete cellulare. Nelle maglie viene trattenuta una piccola percentuale di materiale di natura non mucopeptidica tra cui gli acidi teicoici (polimeri del ribitolo-fosfato o del glicerolo-fosfato) che
costituiscono le componenti antigeniche (risposta immunitaria) della parete, responsabili della risposta anticorpale Gram-negativi (colorazione diversa) Hanno ancora la parete costituita da peptidoglicano, però molto più sottile rispetto ai positivi (rappresenta solo il 10% della composizione della parete): risulta quindi meno resistente da un punto di vista meccanico. Per il resto c’è un rivestimento esterno o pseudocapsula trilaminare con un doppio strato fosfolipidico più proteine, lipoproteine e lipopolisaccaridi (costituiscono circa il 40% della parete dei gram-negativi), che comprendono gli antigeni somatici o antigeni 0 (catene polisaccaridiche di una 40ina di monomeri, diversi nei vari batteri, responsabili della specificità antigenica), un core polisaccaridico (molto simile nei diversi gram- negativi) e più all’interno il lipide A (un glicofosfolipide costituito da acidi grassi uniti a un disaccaride) che si identifica con i LPS responsabili della tossicità dei gram-negativi (endotossine). I diversi antigeni 0 possono essere utilizzati per l’identificazione (sierologica) di diverse specie in quanto sono diversi da batterio a batterio. CI possono essere delle proteine coinvolte nella formazione di pori ( porine ) utili alla permeabilità di molecole di piccola dimensione. Nella parete di tutti i batteri ci sono sempre enzimi autolitici in grado di degradare i costituenti muco- peptidici, i quali consentono ad una struttura rifida quale è la parete cellulare di espandersi ad esempio per generare cellule figlie. La tendenza di alcune colonie batteriche di perdere la gram-positività con l’invecchiamento può essere ricondotta alla disordinata attività di questi enzimi idrolitici: la cellula si è divisa molte volte e produce enzimi idrolitici; alcuni vengono persi e quindi con l’invecchiamento il peptidoglicano viene perso. Batteri privi di parete cellulare È possibile distruggere la parete cellulare trattando il batterio con il lisozima , enzima che spezza il legame glicosidico del peptidoglicano. I gram positivi privati della parete prendono il nome di protoplasti , mentre i gram negativi mantengono il manicotto di LPS, per cui sono più resistenti alla lisi e prendono il nome di sferoblasti. Es i microplasmi , che sono appunto pleomorfi, hanno però una membrana plasmatica ricca di steroli, che li proteggono dalla lisi osmotica, e la membrana sarà più resistente. Altro esempio sono i microrganismi alofili , che vivendo ad alta concentrazione salina non sono limitati da una vera e propria parete (non ne necessitano a causa del loro habitat). Tutti i batteri privi di parete sono relativamente fragili, non hanno una forma definita, si moltiplicano più lentamente degli altri e ovviamente sono resistenti agli antibiotici che agiscono sulla parete. Membrana cellulare o Citoplasma Permette alla cellula di mantenere la sua identità, cioè la delimita dall’ambiente esterno. Nei batteri ha uno spessore di circa 8 nm e ha struttura simile a quella degli eucarioti, cioè ha struttura bilaminare costituita da fosfolipidi (acidi fosfatidici, fosfatidilglicerolo, fosfatil-serina) e proteine (transmembrana o di superficie). A differenza degli eucarioti invece non contiene steroli. D’altra parte, nei procarioti può contenere gli opanoidi , molecole complesse che rendono più solida la membrana. Com’è fatta? (Chiesta all’esame) I fosfolipidi sono molecole anfifiliche, con testa idrofilica polare carica e code idrofobiche apolari non cariche. La testa si orienta nell’ambiente acquoso e verso il lume. Nella membrana ci sono delle proteine Funzioni di membrana
- Il trasporto di molecole all’interno/esterno della cellula: permeabilità selettiva, trasporto attivo recettore-mediate e diffusione passiva per osmosi
- Le secrezioni di enzimi extracellulari, ad esempio per demolire molecole complesse esterne e portare dento le subunità tramite permeasi
- La respirazione e la fotosintesi: a livello dei mesosomi , che sono invaginazioni della membrana plasmatica con estensioni a forma di vescicole, tubuli o lamelle recanti gli enzimi della fosforilazione ossidativa
- La regolazione della riproduzione: proteine della membrana si legano al DNA in fase replicativa e favoriscono la separazione dei cromosomi neoformati; anche il setto trasverso si forma dalla membrana o dai mesosomi
A differenza degli eucarioti, i flagelli dei batteri sono abbastanza rigidi, si muovono con movimento rotatorio invece che per sbattimento, spostando la cellula ( chemiotassi = spostamento verso le sostanze nutritizie e allontanamento da sostanze antibatteriche). Si muovono in senso orario o antiorario per avvicinarsi o allontanarsi dal target. Raramente i batteri a forma sferica hanno flagelli. Filamenti assiali Sono presenti in un gruppo particolare di microrganismi mobili, le spirochete ; sono costituiti da due gruppi di fibre che originano da due estremità opposte della cellula e sembrano sovrapporsi nel mezzo. Dal punto di vista chimico e strutturale sono simili ai flagelli, però sono ricoperti da uno strato di materiale della parete cellulare. I batteri dotati di filamenti assiali si muovono mediante rotazione attorno all’asse longitudinale e mediante ondulazione e flessione. Questi filamenti avvolgono intorno la spirocheta, sono fibre che sembrano appunto sovrapporsi verso il centro, non sono realmente delle sporgenze, infatti sono normalmente ricoperti da uno strato di materiale della parete cellulare. Non conferiscono un movimento come quello dei flagelli, ma un movimento di rotazione intorno all’asse longitudinale. Le fibrille originano dalle due estremità del microrganismo (es spirocheta ) sovrapponendosi al centro e sono rivestite dalla parete cellulare. Pili o fimbrie Sono tipici dei batteri gram-negativi , non hanno un ruolo nella mobilità, sono strutture tubulari cave che emergono dalle cellule, più dritte, corte e sottili dei flagelli (visibili solo al microscopio elettronico). Sono di natura proteica, per cui hanno proprietà antigeniche. Si distinguono almeno in due tipi:
- Pili sessuali : distinguono batteri che li hanno da batteri che non li hanno (sex pili o pili F della fertilità) che intervengono nel fenomeno della coniugazione, che avviene tra batteri che hanno i pili sessuali e batteri che invece hanno un altro genotipo, e sono privi di questi pili (trasferimento di materiale genetico tra batteri)
- Pili comuni : molto più numerosi e corti, che conferiscono al batterio proprietà di adesione ( adesine ) ai tessuti dell’ospite (es ceppi enterotossigeni di E. Coli hanno pili che li ancorano all’apparato gastroenterico da cui altrimenti verrebbero eliminati per il flusso di liquidi organici e per i movimenti peristaltici) Le fimbrie sono osservabili solo al microscopio elettronico, sono di natura proteica e instaurano legami molto specifici con recettori presenti sulle cellule dell’ospite. STRUTTURE INTRACITOPLASMATICHE Tutti i batteri possiedono una regione nucleoide in cui è addensato il materiale genetico; il restante citoplasma contiene un elevato numero di ribosomi (fondamentali per la sintesi proteica) e mitocondri. In più alcuni batteri possiedono inclusioni citoplasmatiche. Il resto del citoplasma è costituito da acqua, enzimi e piccole molecole. Nucleoide È la regione in cui si trova l’informazione genetica del batterio, che è un singolo cromosoma circolare a doppio filamento non circondato da membrana (la cellula quindi dal punto di vista genetico è aploide ). Il cromosoma è una singola lunga molecola di DNA “nuda” (non è compensato con delle proteine, anche se in alcuni casi sono state scoperte delle proteine a stabilizzare il DNA; diverso dal nostro che è complessato con delle proteine chiamate istoni ) che se distesa raggiunge 1 mm (cioè 1000 volte più lunga del batterio). È circolare chiusa covalentemente e superspiralizzata (“impacchettamento”). La singola molecola di DNA contiene tutte le informazioni genetiche del batterio. Il nucleoide, che si trova normalmente addossato ai mesosomi, va incontro a divisione quando la cellula si divide. Talvolta si possono osservare temporaneamente due nucleoidi, quando la cellula si sta dividendo e il setto di separazione tra le due cellule figlie si forma in ritardo rispetto alla replicazione del DNA; il genoma si è replicato, ma siamo in fase di divisione cellulare, quindi i due nucleoidi saranno disposti uno in una cellula figlia e uno nell’altra.
Il cromosoma varia in dimensioni a seconda del tipo di batterio, può contenere da 1000 (micoplasmi) a 10. geni diversi (cianobatteri). L’E. Coli codifica per circa 3000 prodotti genici (4x10^6 paia di basi). Informazioni genetiche addizionali (utili ma non necessarie) possono essere presenti sui plasmidi. Plasmidi Si tratta di DNA circolare a doppio filamento che contiene:
- Un’origine di replicazione autonoma (ori, sito di origine di replicazione del plasmide, che assicura replicazione e segregazione nelle due cellule figlie)
- Da 50 a 100 geni con informazioni per la sopravvivenza del batterio ma non necessarie (es resistenza ad antibiotici o fattore R; produzione di tossine, portate da geni che stanno sul plasmide, vantaggio per il batterio che lo rende patogeno e lo aiuta a prendere il sopravvento sull’ospite; produzione di appendici adesive; produzione di pili sessuali o fattore F, fattore della fertilità, quello che codifica i geni dei pili sessuali per il meccanismo della coniugazione) Alcuni plasmidi possono perdere la vita autonoma e reintegrarsi nel cromosoma (episomi) e replicarsi in sintonia con il cromosoma stesso; un episoma può risepararsi dal cromosoma e ricominciare la replicazione autonoma come plasmide; ad un certo punto, excisione, il plasmide torna a vivere la sua vita autonoma ed esce dal cromosoma. La maggior parte dei batteri contiene più plasmidi. Il plasmid fingerprinting (isolamento, purificazione e restrizione del plasmide) può essere utilizzato per l’identificazione batterica; ceppi diversi di batteri della stessa specie possono portare dei plasmidi diversi; questo fa sì che i tipi di plasmidi presenti in un determinato batterio possano essere utili per la classificazione (molto più dettagliate rispetto alla specie). Ribosomi Nel citoplasma di un batterio ci sono fino a 15.000 ribosomi che funzionano nella sintesi delle proteine; hanno diametro di 20 nm, più piccoli e più leggersi di quelli eucarioti (coefficiente di sedimentazione, legato alla dimensione, 70S e sono costituiti da due subunità, più piccole rispetto a quelle degli eucarioti, una maggiore e una minore, di 50S e 30S). Ciascuna delle due subunità è costituita a sua volta da rRNA e proteine. L’mRNA arriva a livello delle subunità del ribosoma e il tRNA riconosce la sequenza portata dal messaggero e attacca gli amminoacidi gli uni agli altri a formare una proteina. La diversa composizione chimica rispetto ai ribosomi degli eucarioti ne condiziona la sensibilità selettiva ad alcuni antibiotici (es streptomicina ). I ribosomi dei procarioti sono simili a quelli dei mitocondri, il che potrebbe giustificare la teoria dell’origine endosimbiontica. Materiali di riserva Molti batteri formano granuli di riserva nel loro citoplasma, osservabili al microscopio ottico dopo la colorazione (talvolta fenomeno della metacromasia , ovvero l’acquisizione di un colore diverso da quello del colorante utilizzato): es globuli di glicogeno o di amido (es nei clostridi ) costituiscono riserva di carbonio ed energia. I granuli di polifosfati (verosimilmente prodotti intermedi del metabolismo cellulare e utili nella sintesi degli acidi nucleici) si osservano in corinebatteri e yersinia. Granuli di lipidi si osservano in micobatteri (probabilmente correlati con la virulenza e la patogenicità). La sintesi di inclusioni granulari varia in funzione del tipo di terreno colturale e della fase di crescita del microrganismo. Spore Due generi di batteri, i bacillus e i clostridium , hanno la capacità per cui nel momento in cui le condizioni ambientali diventano sfavorevoli per loro, formano delle strutture che gli permettono di mantenersi in forma quiescente, senza metabolismo attivo, per tornare “in vita” quando le condizioni ambientali migliorano. In determinate condizioni la cellula batterica di alcuni bacilli gram-positivi presenta al suo interno una spora ( endospora ) con caratteristica di resistenza verso agenti chimici, fisici e sterilizzanti e verso l’invecchiamento (le spore sopravvivono anche per migliaia di anni, sopravvive anche a trattamenti batterici abbastanza forti). La scoperta delle endospore dei batteri è stata fondamentale nella microbiologia in quanto ha consentito lo sviluppo di adeguati metodi di sterilizzazione. I batteri che producono spore si dicono sporigeni : possono trasformarsi in queste strutture quiescenti e mantenersi senza crescere né riprodursi per lungo tempo. La spora viene prodotta a partire da una cellula batterica attraverso il processo di sporulazione quando le condizioni ambientali diventano sub-ottimali (es assenza di un nutriente). Prima fase: preparazione della spora, tutto il materiale genetico, tutto il ribosoma e
di nuove particelle virali complete o virioni. I virus portano quello che sanno di non trovare all’interno della cellula ospite (che andranno a infettare); devono quindi portare i siti di assorbimento, necessari per entrare nella cellula, con altri enzimi o molecole, oltre a enzimi per avviare la replicazione. Sono assenti dal virus gli enzimi per il metabolismo e la produzione di energia, e tanti altri enzimi per la loro replicazione che vengono forniti dalla cellula ospite; si capisce il parassitismo endocellulare del virus. Portano invece gli enzimi che la cellula non può fornire (es enzimi per iniziare l’infezione, per avviare la replicazione). I virus sono entità completamente diverse dagli altri agenti patogeni:
- Non hanno un’organizzazione strutturale/funzionale di tipo cellulare
- Il DNA non è sempre il depositario dell’informazione genetica, ma nei virus a RNA è l’RNA, nei virus non possono coesistere i due acidi nucleici
- Persistenza nell’ospite di alcuni virus attraverso l’integrazione del DNA (o cDNA) nel genoma della cellula ospite, il fatto di poter rimanere indefinitamente nella cellula ospite finché l’animale muore. Particolarità di integrare il proprio genoma in quello dell’ospite, integrazione che non si scioglie mai; quando vengono meno le nostre difese immunitarie, il virus prende il sopravvento e inizia a moltiplicarsi. Portano due enzimi, una trascrittasi inversa e un enzima che degrada il DNA, quindi sono in grado di sintetizzare un doppio filamento di DNA e poi integrato nel nostro genoma (prima molecola ibrida di DNA e RNA, poi l’RNA viene sostituita con un altro filamento e suo essere trascritta nel nostro DNA)
- Non sono sensibili ad antibiotici ma a specifici chemioterapici antivirali
- Non hanno una curva di crescita come quella batterica, ma immediatamente dopo l’infezione “scompaiono” come particelle infettanti perché entrano nelle cellule target, dove si moltiplicano e poi “ricompaiono” improvvisamente dopo qualche ora moltiplicati di un fattore di 100-150 e poi non c’è più un ulteriore incremento dopo questa rapida amplificazione In generale i virus vengono divisi in tre principali categorie secondo la natura dei loro ospiti:
- Virus animali : infettano insetti, molluschi, pesci, rettili, anfibi, uccelli e mammiferi
- Virus vegetali : secondi dopo i funghi nel provocare malattie di importanza economica nelle piante
- Virus batterici : o batteriofagi, che infettano limitatamente i batteri e sono in genere molto specifici per un certo genere batterico Struttura dei virus (o virioni) I virioni sono particelle virali mature infettive. I virus più semplici sono costituiti da acido nucleico + capside. L’acido nucleico è in genere strettamente associato a proteine con funzione stabilizzante e prende il nome di core ; si trova in genere all’interno di un rivestimento chiamato capside , formata da molte subunità di natura proteica. Il capside può essere icosaedrico o a simmetria elicoidale filamentoso o a bastoncino rigido con o senza ulteriori rivestimenti. Vengono definiti nudi i virus con capside sia elicoidale (virus non molto regolare, aspetto più amorfo, struttura non rigida, si ripiega e aggomitola nello spazio a creare un aspetto meno regolare) che icosaedrico non ulteriormente rivestiti dall’envelope. Altra particolarità dei virus animali con struttura elicoidale è la presenza dell’envelope (membrana esterna di natura lipoproteica a doppio strato fosfolipidico) con duplice funzione:
- “Mascherare” le proteine antigeniche del capside in modo che non vengano riconosciute e attaccate dal sistema immunitario dell’ospite
- Iniziare l’infezione tramite strutture definite siti di assorbimento o di adesione che consentono il legame con uno specifico recettore presente sulle membrane cellulari Il capside è una struttura composta da un numero definito di unità morfologiche chiamate capsomeri collegate tra loro da legami non covalenti, sono i singoli mattoncini necessari a costituire i capsidi, sono a tutti gli effetti delle proteine virali sintetizzate e prodotte dal macchinario cellulare, il virus infetta la cellula, blocca la sintesi delle proteine della cellula e fa produrre alla cellula le proprie proteine virali. Esso viene sintetizzato all’interno della cellula infetta secondo una simmetria ben definita. Le uniche due simmetrie osservato per il capside sono:
- Simmetria elicoidale (struttura a spirale, cava all’interno): il prototipo di questo genere di virus è rappresentato dal virus del mosaico del tabacco (TMV) che non è un virus animale, costituito da bastoncini rigidi lunghi circa 300 nm e del diametro di circa 15-17 nm. Osservato nella direzione dell’asse maggiore, appare esagonale con un foro centrale di 4 nm di diametro. Struttura di un’elica
sinistrorsa costituita da subunità proteiche o capsomeri connessi tra loro a formare dei dischi sovrapposti. Tutti i capsiomeri sono identici tra loro e costituiti da una singola molecola proteica di 17kD. La parte più interna della catena polipeptidica è flessibile e prende contatto con l’RNA genomico virale, che viene così alloggiato in un avvallamento tra i dischi e ne segue l’andamento a spirale. Il nucleocapside dei virus elicoidale può essere rigido, come nel TMV, con aspetto a bastoncello, o flessibile, come nel caso dei virus animali. Può arrotolarsi su sé stesso e assumere forme diverse (es a “gomitolo di lana” tipico dei paramyxovirus ). Questa struttura, nei virus animali, è avvolta nell’envelope
- Simmetria icosaedrica : molti virus hanno un capside che sembra sferico, però ad un più attento esame si verifica che è di forma poliedrica, più preceisamente icosaedrica (poliedro con 12 vertici e 20 facce ciascuna a triangolo equilatero). La forma icosaedrica rappresenta la soluzione per creare una struttura stabile che racchiuda il massimo volume possibile (per l’acido nucleico). I capsomeri a loro volta sono costituiti da un numero variabile (3, 4, 5, 6) di subunità strutturali (monomeri detti protomeri ), pertanto possono essere: o Trimeri : adenovirus, iridovirus o Tetramero : picoronavirus o Pentameri : papovavirus o Esameri : herpesvirus Possono essere a un unico peptide (omo oligomeri) o a due o più peptidi diversi (etero-oligomeri). I capsomeri dei virus degli animali sono in genere costituiti da almeno tre o quattro proteine strutturali diverse (protomeri) che si auto-assemblano spontaneamente a costituire il capsomero. I capsomeri poi sono le unità morfologiche che costituiscono il capside. Ci sono minimo 60 capsomeri, che costituiscono il capside; nei virus più complessi invece sono di più (es gli adenovirus hanno 252 capsomeri; i polyomavirus 72 capsomeri) Il virus della peste suina africana presenta una membrana lipidica aggiuntiva tra il capside e le strutture più interne, è una particolarità unica. Nei batteriofagi più complessi a DNA, sono presenta una testa a simmetria simil-icosaedrica, una coda a struttura elicoidale e un colletto contrattile. COMPOSIZIONE CHIMICA DEI VIRUS
- Proteine : sono componenti necessarie, presenti in tutti i virus e sintetizzate secondo l’informazione contenuta nel virus. Si distinguono in due grandi categorie, le proteine strutturali , che costituiscono il capside e l’envelope (in alcuni virus più complessi ci sono proteine associate agli acidi nucleici, come negli adenovirus, herpesvirus, polyomavirus ) e quelle non strutturali (o regolatorie), che hanno diverse funzioni: sono enzimi coinvolti nella replicazione virale (RNA polimerasi, DNA polimerasi, enzimi per modificazioni post-traduzionali), ma anche proteine “interferenti” con la sintesi proteica dell’ospite deregolando geni cellulari e/o transattivando geni. Ci sono proteine che influenzano la virulenza o il tropismo del virus (es inibizione della risposta immunitaria dell’ospite)
- Acidi nucleici : presenti necessariamente in tutti i virus. Tutti i genomi virali sono aploidi , ovvero contengono una sola copia di ogni gene, tranne i retrovirus , che sono simil-diploidi, due copie identiche di RNA (al momento non ne è noto il significato biologico). Attualmente il genoma di molti virus di impatto medico e veterinario è noto ( sequenziato ). Il genoma può essere a DNA o a RNA , ma in maniera esclusiva, per cui ci può essere un solo acido nucleico. Le dimensioni dei genomi a DNA variano da 1.7kb (kilobase) del single strend DNA dei circovirus a più di 200kb dei double strend DNA degli herpesvirus e dei poxvirus. ➢ Genoma a DNA costituito da: o Una singola molecola a doppio filamento (ds) lineare continuo (es adenovirus ) oppure frammentato (es herpesvirus ) o Una singola molecola a doppio filamento (ds) circolare continuo (es paopovavirus ) oppure frammentato (es hepadnavirus ) o Una singola molecola a singolo filamento (ss) lineare (es parvovirus ) o Una singola molecola a singolo filamento (ss) circolare (es circovirus )
dà luogo ad una viremia secondaria , in particolare il virus si replica nelle cellule target specifiche (es i virus neurotropici, le cellule neuronali ecc). La viremia , ovvero la presenza di particelle virali nel sangue, può essere quantificata, mantenersi alta oppure può essere controllata dal sistema immunitario dell’ospite, o ancora può andare incontro a cicli che dipendono dall’equilibrio tra la replicazione del virus e la risposta immunitaria dell’ospite, in base alla diversa gravità della malattia. L’evento finale è la disseminazione all’esterno del soggetto infettato. Morte o guarigione sono il risultato dell’equilibrio dinamico e dell’interazione del virus con l’animale infettato. TIPI DI INFEZIONI VIRALI Le infezioni virali possono essere acute o persistenti:
- Infezioni acute : sono caratterizzate dalla comparsa di sintomatologia evidente, che compare in tempo breve dal contagio; l’esito dipende dal virus ed è la morte o la guarigione (in genere accompagnata da immunità duratura) del soggetto. I virus possono causare: ▪ Lesioni cellulari : possono essere più o meno gravi, e includono la destabilizzazione delle membrane lisosomiali (con conseguente rilascio degli enzimi autolitici), l’inibizione della sintesi dell’RNA cellulare (può riguardare mRNA, rRNA), l’inibizione della sintesi del DNA cellulare (come conseguenza delle precedenti due inibizioni). In tutti questi casi si hanno alterazioni delle cellule per cui le cellule non possono più replicarsi ▪ Alterazioni delle funzioni cellulari : possono avvenire attraverso la proliferazione delle membrane endocellulari (nucleari oppure degli organelli intracitoplasmatici; le membrane possono essere degradate ad opera della fosfolipasi) oppure l’induzione di enzimi per la sintesi del DNA virale (che possono indurre la cellula ad andare in mitosi o a proliferazione neoplastica) ▪ Predisposizione alle infezioni secondarie : ad opera di microrganismi opportunisti, generalmente batteri, tra cui è noto il caso di associazione tra i Rotavirus e i Coronavirus bovini e Pasteurella (polmoniti) o E. Coli (enteriti) ▪ Danni al sistema immunitario : tra cui deplezione dei linfociti B (con conseguente immunodepressione), alterazione delle funzioni dei linfociti B (con conseguente carenza di anticorpi), deplezione dei linfociti B e T (come nel caso di FelV ), deplezione dei linfociti T (con conseguente immunodepressione) ▪ Lesioni immuno-mediate : possono essere ADCC (citotossicità cellulo-mediata anticorpo- dipendente, si verifica perché le cellule infette sulla cui superficie sono legati gli anticorpi specifici attivano le cellule che possiedono il recettore per le Fc delle immunoglobuline), lesioni da immunocomplessi (quando gli antigeni virali sono in forte eccesso rispetto agli anticorpi specifici, quindi si formano immunoprecipitati), ipersensibilità ritardata (che comporta l’accumulo di linfociti T citotossici in diversi tessuti) e infine malattie autoimmuni
- Infezioni persistenti : sono caratterizzate dalla permanenza del virus nell’organismo per periodi molto lunghi (anche l’intera vita dell’animale). Gli animali infetti costituiscono un’importante fonte di contagio, tali infezioni possono riattivarsi anche più volte nello stesso individuo dando luogo ad episodi acuti (es Herpesvirus ); possono causare disordini immunopatologici che possono accompagnarsi a infezioni secondarie (es Retrovirus ) o causare neoplasie (es Papillomavirus ). Le infezioni persistenti a loro volta possono distinguersi in: ▪ Infezioni latenti : in cui le particelle virali complete non sono mai dimostrabili se non durante i periodi di acutizzazione, mentre il genoma virale è dimostrabile (es virus herpetici nei gangli nervosi ) ▪ Infezioni croniche : in cui le particelle virali sono sempre dimostrabili e il virus viene costantemente o saltuariamente eliminato dall’animale infetto nell’ambiente ▪ Infezioni lente : caratterizzate da lenta progressione della malattia e dall’aumento progressivo delle particelle virali presenti nell’organismo ed eliminate nell’ambiente Le cause della persistenza possono essere ricercate o nell’agente patogeno o nel sistema immunitario dell’ospite. Le cause dovute all’agente patogeno sono: possibile assenza di immunogenicitià (come nel caso della scrapie o dei virus adeno-associato ); l’integrazione dell’acido nucleico nel genoma
cellulare (con replicazione contemporanea alla mitosi); moltiplicazione in siti protetti (es cellule nervose); variazioni antigeniche del virus. Le cause dovute al sistema immunitario dell’ospite sono: difettosa risposta anticorpale; difettosa risposta cellulo-mediata; replicazione all’interno dei macrofagi; alterazione dell’MHC (sistema maggiore di istocompatibilità) OSSERVAZIONE E STUDIO DEI MICRORGANISMI I microrganismi sono esseri viventi invisibili ad occhio nudo … (definizione). I batteri hanno dimensioni che vanno da 0,5 a 10 mm, i virus da 10 a 300 nm, pertanto per visualizzarli sono necessari strumenti e accorgimenti con un adeguato limite di risoluzione (= distanza minima tra due punti tale per cui risultino distinti). Per visualizzare i batteri si utilizza la microscopia ottica. Per visualizzare i virus si utilizza la microscopia elettronica. Microscopia ottica I moderni strumenti sono composti, con almeno due lenti, una obiettivo e una oculare (es combinando un obiettivo a 40 ingrandimenti con un oculare a 10 ingrandimenti si ottiene un’immagine ingrandita 400 volte). In genere i microscopi in uso possiedono più obiettivi intercambiabili mediante rotazione del blocco portante. Gli obiettivi a immersione in olio sono quelli più usati portandoli a contatto con una goccia di olio di legno di cedro deposta sul preparato da osservare. Può essere:
- In campo illuminato (o campo chiaro) : è la classica microscopia usata in microbiologia. Il limite di risoluzione dipende dalla lunghezza d’onda della luce impiegata e dal suo angolo di entrata negli obiettivi. Con lo spettro del visibile (l = 400-700 nm) il limite inferiore di risoluzione è 0.2 mm, pertanto ben pochi dettagli possono essere messi in evidenza direttamente con questo tipo di microscopio. La risoluzione aumenta con la colorazione, ma ovviamente ciò comporta che le cellule muoiano o si modifichino, quindi non sempre è adeguato allo studio di determinate caratteristiche. Si può arrivare a 0,1 micrometri usando lenti a immersione in olio. Il campione è visualizzato grazie alla luce che passa direttamente dal condensatore al vetrino. Si può optare per diversi ingrandimenti: 10x (per visualizzare l’intero campione), 40x (utile per osservare microrganismi di grosse dimensioni), 100x (per osservare batteri, miceti e alcuni dettagli morfologici)
- In campo oscuro : microscopia ottica nella quale il sistema di lenti è modificato; c’è un condensatore che fa in modo che la luce non raggiunga direttamente il campione, ma questo è raggiunto solo dalla luce diffusa e obliqua, pertanto questo appare come un’immagine luminosa su sfondo scuro. Il potere di risoluzione è 10 volte maggiore di quello della microscopia a campo chiaro (arriva a distinguere dettagli di 0,2 micron) e consente di identificare i batteri più sottili. Vantaggio è una maggiore risoluzione , svantaggio che poiché la luce non attraversa il campione, consente di visualizzare solo i contorni e non le strutture interne di un batterio
- A contrasto di fase : ovvia all’inconveniente della microscopia in campo oscuro, infatti consente di esaminare i dettagli interni del campione. Si basa sul principio per cui quando i raggi luminosi attraversano un corpo trasparente alcuni vengono ritardati a causa della diversa densità delle zone dello stesso. Questo ritardo fa sì che i raggi si trovino “fuori fase” rispetto al resto dei raggi luminosi. Tali differenze (non apprezzabili con altri microscopi) vengono trasformate da questo particolare microscopio in differenze di intensità luminosa. In questo modo senza colorazioni è possibile vedere strutture tridimensionali anche minime all’interno di cellule viventi
- A fluorescenza : viene impiegata la luce UV come fonte luminosa (poiché I è circa la metà di quella della luce visibile, la risoluzione è molto maggiore). Non essendo la luce UV percepibile dall’occhio umano, questo microscopio consente di visualizzare solo i campioni fluorescenti o per loro caratteristica naturale (es microrganismi fluorescenti) oppure perché sono stati trattati con coloranti fluorescenti o con anticorpi marcati fluorescenti. Il colorante fluorescente o fluoroforo/fluorocromo è un composto che assorbe la luce ad una determinata I (eccitazione) ed emette a I maggiore. In particolare, assorbe nello spettro dell’ultravioletto (I = 10-400 nm) ed emette nello spettro del visibile. Il microscopio usa speciali lampade a vapori di mercurio o di xenon che generano una lunghezza d’onda inferiore rispetto alla luce emessa dai microscopi ottici tradizionali (potere di risoluzione maggiore). Il microscopio invertito ha la sorgente di luce ed il condensatore in alto (sopra il tavolino
poi mediante speciali ultramicrotomi si eseguono sezioni ultrasottili del preparato per l’esame al microscopio. Poiché generalmente i componenti dei campioni biologici possiedono una massa ridotta, il contrasto prodotto dal passaggio del fascio di elettroni è modesto. Pertanto per ottenere sufficiente contrasto in genere le preparazioni vengono trattate con sali di diversi metalli pesanti (piombo, tungsteno, uranio), che avendo elevato peso atomico, disperdono marcatamente il fascio di elettroni (colorazione positiva, colorazione negativa, ombreggiatura) Fissazione, disidratazione, inclusione e colorazione possono danneggiare le strutture cellulari, pertanto bisogna stare attenti all’interpretazione dei risultati. Per prevenire la formazione di artefatti è stata messa a punto la tecnica del “ freeze-etching ”, in cui il campione viene congelato senza alcun trattamento chimico e il blocco congelato viene fratturato in modo da esporre le porzioni interne o esterne delle cellule. Dopo sublimazione del ghiaccio si ottiene un “bassorilievo” o “replica” della superficie di frattura, che viene poi coperta di metalli pesanti. Il campione biologico poi viene distrutto e rimane la replica che riproduce i dettagli della superficie esaminata. Fasi della preparazione del campione
- Prelievo
- Fissazione : per TEM e SEM con protocolli simili, in genere con tetrossido di osmio
- Disidratazione : per TEM e SEM con EtOH, acetone, diossido di carbonio al punto critico liquido
- Inclusione : in resina per TEM
- Taglio : sezioni ultrasottili con microtomo per TEM
- Montaggio : su retino per TEM, su una matrice per SEM
- Staining con metalli pesanti per contrasto d’immagine : TEM e SEM RIPRODUZIONE DEI BATTERI La scissione binaria è il metodo più comune tra i batteri: una cellula madre si scinde in due cellule figlie dopo che ha replicato il genoma; c’è la possibilità di frammentazione o gemmazione (tipica dei lieviti, i quali sono eucarioti unicellulari). Si tratta di un processo asessuato. 20- 30 minuti è la tempistica di replicazione di molti batteri in laboratorio in condizioni ottimali: il tutto dipende dalla concentrazione di ossigeno, umidità, pH ecc (in natura i tempi possono essere molto più lunghi, per esempio l’E.Coli impiega anche 12 ore nell’intestino). Non tutti sono così veloci, i microbatteri arrivano minimo a 18 ore anche in condizioni ottimali. Occorre il microscopio ottico per visualizzare la cellula batterica, ma se si fa crescere in laboratorio i batteri crescono in modo raggruppato creando una colonia e perciò questa sarà visibile ad occhio nudo. Le colonie di batteri (su terreno solido) che hanno un rapido periodo di replicazione, si sviluppano da un giorno all’altro (18-24 ore); mentre i microbatteri hanno anche bisogno di 30 - 40 giorni. La scissione binaria si realizza attraverso fasi successive: prima avviene la duplicazione del cromosoma, poi la cellula si allunga e la membrana citoplasmatica si estende in senso centripeto, in genere a livello di un mesosoma, formando un setto trasverso, che separa in due cellule identiche (1 cromosoma per cellula); infine la parete cellulare nella regione mediana della cellula forma un restringimento anulare e si sviluppa una parete trasversa, che completa la separazione delle due cellule figlie. La gemmazione (nei lieviti) si crea una protuberanza sulla superficie della cellula madre che aumenta gradualmente di volume, può essere terminale o laterale. Colonia o clone : i batteri possono moltiplicarsi in terreno liquido (da due a cinque batteri visibili come aggregati/colonie) e solido (non sono mobili, il batterio rimane stabile in un determinato punto e sarà visibile finché non ci sarà un aggregato di 10^6 - 107 batteri, ovvero 1 milione-10 milioni).
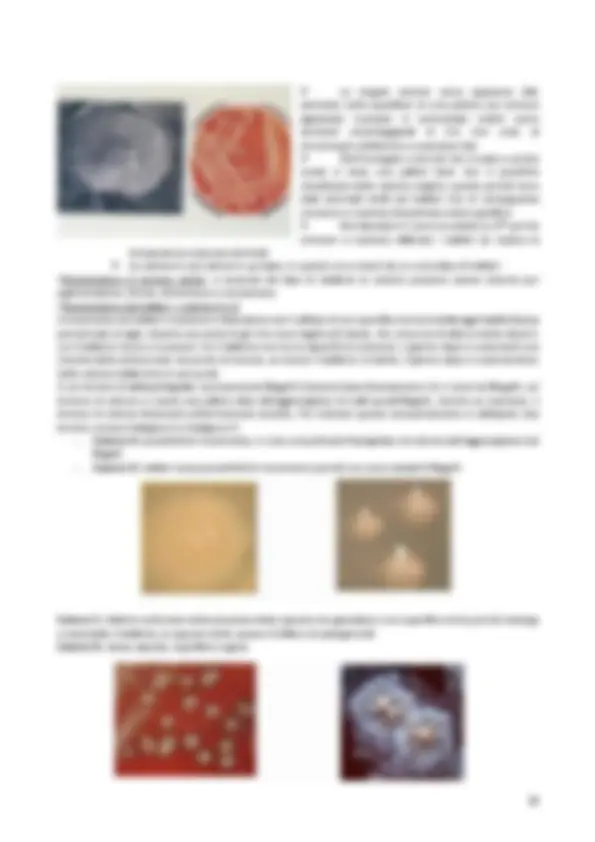
➢ Le singole colonie come appaiono (dx) seminate sulla superficie di una piastra con terreno agarizzato incubate in termostato visibili come ammassi rotondeggianti di 2 - 6 mm viste al microscopio elettronico a scansione (sx) ➢ Nell’immagine a dx (ciò che si vede a occhio nudo) si vede una patina dove non è possibile visualizzare delle colonie singole: questo perché sono stati seminati molti più batteri che di conseguenza crescono in maniera disordinata sulla superficie ➢ Nei laboratori ci sono incubatori a 37° per far crescere a maniera ottimale i batteri (si replica la temperatura corporea animale) ➢ La colonia è una coltura in purezza, in quanto va a crearsi da un unico tipo di batteri
- Riproduzione in terreno solido: a seconda del tipo di batterio le colonie possono essere diverse per pigmentazione, forma, dimensione e consistenza.
- Riproduzione dei batteri – colonie H e O Il movimento dei batteri si osserva in laboratorio con l’utilizzo di uno specifico terreno detto agar molle (bassa percentuale di agar, diventa una sorta di gel che crea maglie più lasse), che crea una struttura molto lassa in cui il batterio riesce a muoversi. Se il batterio non ha la capacità di muoversi, il giorno dopo si osserverà una crescita della colonia solo nel punto di inoculo, se invece il batterio è mobile, il giorno dopo si osserveranno delle colonie batteriche in vari punti. In un terreno di coltura liquido , la presenza dei flagelli è determinata diversamente. Se ci sono dei flagelli, sul terreno di coltura si creerà una patina data dall’aggrovigliarsi di tutti questi flagelli, mentre se mancano, il terreno di coltura diventerà uniformemente torbido. Per indicare questo comportamento si utilizzano due termini, ovvero Antigene H e Antigene O:
- Colonie H : possibilità di movimento, si crea una pellicola frastagliata che deriva dall’aggrovigliarsi dei flagelli
- Colonie O : batteri senza possibilità di movimento perché non sono dotati di flagelli Colonie S : distinte sulla base della presenza della capsula che garantisce una superficie liscia perché avvolge a manicotto il batterio; la capsula molto spesso è fattore di patogenicità Colonie R : senza capsula, superficie rugosa