Scarica Microbiologia sbobina e più Sintesi del corso in PDF di Microbiologia solo su Docsity!
Microbiologia generale
Capitolo 1
1.1 Il mondo dei microrganismi
Vivente = un sistema in disequilibrio termodinamico con l’ambiente esterno, capace di mantenersi, riprodursi e di evolvere. Tutti gli organismi viventi sono costituiti da cellule. Alcuni organismi sono costituiti da una sola cellula ( unicellulari ), altri da più cellule ( pluricellulari ). La maggior parte degli organismi viventi sono unicellulari, questi insieme ad alcuni organismi pluricellulari rappresentano i microrganismi. Definizione di microrganismi = organismi viventi totipotenti permanenti. Le cellule hanno innanzitutto una loro “identità” garantita da un rivestimento, la membrana plasmatica , che le delimita e le isola e allo stesso tempo le mette in comunicazione con l’ambiente esterno. Grazie ad essa la cellula è anche in grado di mettere in atto processi di adattamento che le consentono di mantenere la propria integrità ( omeostasi ). Secondo l’assioma di Rudolph Virchow , ogni cellula esistente deriva dalla divisione di un’altra cellula. Il mantenimento e la riproduzione cellulare richiedono quindi un continuo approvvigionamento di materia , che dall’ambiente è importata e trasformata attraverso una complessa rete di reazioni, e di energia. È evidente che un sistema che preleva energia e materia dall’ambiente e lo trasforma in cellule non può mantenersi per un tempo indefinito, poiché le risorse utilizzate presto si esauriranno. La contraddizione tra l’imperativo posto alla cellula di “crescere e moltiplicarsi” e la disponibilità non infinita delle risorse è risolta, con la morte e il riciclo delle molecole e degli atomi di cui un organismo è composto. La mancanza di fonti inesauribili di materie prime per la costruzione di nuove cellule porta a una competizione tra organismi viventi per le risorse disponibili. Gli organismi viventi hanno caratteristiche in comune; innanzitutto l’ apparato informatico è costituito dallo stesso tipo di macromolecole (DNA, RNA e proteine). I sistemi di approvvigionamento energetico sembrano più diversificati, consentendo così l’uso sia di energia luminosa sia di energia prodotta da reazioni chimiche, convergendo però la produzione degli stessi composti (ATP e NAD (P) ridotto).
Che cos’è il metabolismo? E’ quel meccanismo per cui c'è un' identità (costituita dal patrimonio genetico). Questa identità evolve e tendenzialmente si ripete in un altro organismo tramite la riproduzione , salvo mutazioni, che portano l’identità a perdersi anche di un minimo. Ad esempio, in una colonia di 100 milioni di cellule, se il tasso di mutazione è di 10-6, significa che su ogni locus ci sono potenzialmente 100 mutazioni. Dopo la riproduzione, le varie forme ottenute per imitazione vengono selezionate e questo porta all’ evoluzione. Tutto questo processo modifica il metabolismo. I suoi processi fondamentali rimangono gli stessi, ma ci sono piccole differenze per cui una specie fa delle cose, e un’altra ne fa altre.
Storia della classificazione dei microrganismi
Intorno alla metà dell’800’ gli esseri viventi si divisero in regni di piante e animali. Nel 1886 Ernst Haeckel propose il nuovo regno dei protisti (protozoi, funghi, alghe e batteri), dove si poteva inserire tutto ciò che non rientrava nei due regni tradizionali. Inoltre egli seguiva una tradizione che affermava che le piante fossero immobili, fotosintetizzanti e assumessero il cibo per assorbimento; mentre gli animali fossero mobili, non fotosintetizzanti e assumessero il cibo per fagocitosi o digestione. I protisti però avevano alcune strutture intracellulari non dissimili da quelle di piante e animali (nucleo delimitato da una membrana nucleare, complesso sistema di membrane interne, organelli racchiusi in membrane).
Per questi ultimi fu coniato il termine di procarioti (E. Chatton), distinti da tutti gli altri organismi detti eucarioti. I procarioti dichiararono così l’indipendenza dai protisti, fondando il nuovo regno Monera.
Un ulteriore scissione venne effettuata da R.H. Whittakker, che propose che i funghi venissero raggruppati nel regno indipendente dei funghi. Woese infine propose di riclassificare il mondo vivente in tre “domini”: Eukarya (piante, animali, funghi e protisti), Bacteria e Archea.
Parete e assorbimento: funghi e alghe si; animali no; Mobilità: funghi e alghe no; animali si; Multicellularità: funghi no; alghe e protozoi si;
LUCA. L’ultimo progenitore ancestrale: LUCA (3,5 miliardi di anni fa). E l’acronimo di last common universal ancestor. Esso è quella cellula che non è un batterio dalla quale è partita l’evoluzione. Da LUCA si distinguono i batteri e gli archaea e da essi comincia la formazione di organelli dentro le cellule (endosimbiosi).
Organismi modello molto utilizzati per lo studio delle strutture e dei meccanismi cellulari, fisiologici, genetici e molecolari in microrganismi eucarioti sono, ad esempio, Saccharomyces cerevisiae (o lievito di birra), che anzi è divenuto il modello il sistema modello per eccellenza di cellula eucariota, o Neurospora crassa, adottata inizialmente per l’analisi genetica del metabolismo e per gli studi sulla meiosi e la ricombinazione. Il batterio più studiato è senza dubbio Escherichia coli. La diversità biologica delle specie microbiche risponde a specifici meccanismi di adattamento che consentono ai microrganismi di vivere negli habitat più disparati e utilizzare le più diverse fonti nutritive.
1.2 Come si costruisce una cellula
Caratteristica di ogni organismo vivente è la capacità di riprodursi, nei batteri e negli Archea, organismi unicellulari e riproduzione non sessuale, ciò coincide con la duplicazione cellulare. Una cellula deve procacciarsi materia ed energia dall’ambiente, mentre l’informazione è specificamente contenuta nella cellula stessa ( informazione genetica ). Le forti similarità nell’impianto molecolare di tutte le cellule fa sì che la composizione chimica elementare sia assolutamente comparabile in tutti gli organismi viventi. H , O , C , N , P , S sono gli elementi presenti strutturalmente nelle macromolecole biologiche. Mentre altri elementi ( Mg, Fe, Ca, K, Na ) sono presenti in forma ionica, liberi o associati alle molecole organiche. Una tipica cellula batterica è costituita da diverse strutture sopramolecolari complesse: la parte più esterna forma una specie di involucro da cui possono protrudere appendici (pili e flagelli) e delimita uno spazio interno costituito da un “citosol”. Queste strutture sopramolecolari sono costituite dall’assemblaggio di pochi tipi di macromolecole: le proteine , presenti in quasi tutte le strutture sopramolecolari, dove svolgono funzioni strutturali o enzimatiche, l’ RNA , il DNA , i fosfolipidi , i lipopolisaccaridi, la mureina e il glicogeno. Amminoacidi, nucleotidi, acidi grassi e monosaccaridi sono fabbricati attraverso complesse reazioni metaboliche che consumano energia, a partire da una dozzina di molecole organiche più semplici (dette metaboliti precursori , carbonio, idrogeno, fosforo) e da due molecole inorganiche ( ammonio e solfato ) che apportano l’azoto e lo zolfo. Non tutti gli organismi sono capaci di assimilare zolfo, azoto e fosforo a partire da solfato, ammonio e fosfato. L’involucro esterno alla membrana plasmatica può mancare o essere molto diverso nei diversi organismi.
Alcuni organismi ricavano energia dalla luce: piante, alghe, molti batteri e archei sono fototrofi , capaci cioè di trasformare l’energia luminosa in energia metabolica. Altri sfruttano l’energia libera rilasciata da reazioni
Il dibattito sulla generazione spontanea fu ripreso a metà del settecento tra Spallanzani e Needham. Quest’ultimo forte sostenitore della generazione spontanea, osservava che campioni di brodo conservati in fiasche chiuse con tappi di sughero dopo pochi giorni diventavano torbidi per la crescita di svariati tipi di organismi microscopici. Spallanzani al contrario, bollendo a lungo i campioni di brodo e sigillando ermeticamente dimostrò che il brodo poteva conservarsi indefinitamente senza che si osservasse crescita di microrganismi. Egli giunse pertanto alla conclusione che la bollitura uccidesse i “germi vitali” presenti nel brodo e che la sigillatura impedisse l’ingresso di nuovi germi capaci di moltiplicarsi nel brodo. Si obiettava però che la bollitura prolungata avrebbe potuto distruggere la forza vitale. Ma si pone sempre il problema dei vitalisti: secondo loro ciò non dimostrava nulla perché sigillando l’ampolla dopo la bollitura non poteva entrare la vis vitalis e ciò era il motivo per cui non si creavano microrganismi.
- Allora intervenne Louis Pasteur: fece l’esperimento dei matracci a collo d’oca.
Tyndall dimostrò l’esistenza di spore resistenti alla bollitura, oggi dette endospore , prodotte da alcuni batteri. Louis Pasteur , dimostrò che il pulviscolo atmosferico è ricco di microrganismi: filtrò diversi campioni di aria attraverso batuffoli di fulmicotone (nitrocellulosa) che trattenevano le particelle solide; la nitrocellulosa veniva poi sciolta con una miscela di etere ed alcol. L’analisi al microscopio rivelava la presenza di numeri microrganismi che assomigliavano a quelli spesso trovati in materiali in decomposizione. Pasteur sviluppò altri esperimenti i cui risultati demolirono definitivamente la teoria della generazione spontanea. L’esperimento più famoso è quello del pallone a collo di cigno. In questo caso un brodo di coltura è posto in un matraccio la cui imboccatura viene modellata a collo di cigno per permettere il passaggio dell’aria ma non quello delle “particelle” microscopiche presenti nell’aria stessa. Il brodo viene quindi portato ad ebollizione in modo da uccidere tutte le forme viventi ivi presenti. Anche dopo un lungo periodo, il brodo non diventa torbido. Solo se il collo del pallone viene rotto permettendo alle particelle sospese nell’aria di penetrarvi, si osserva intorbidimento del brodo dovuto alla crescita di microrganismi presenti nell’aria.
Robert Koch si dedicò allo studio delle cause di molte malattie e diede origine alla microbiologia medica. Un primo contributo importante fu la messa a punto di terreni di coltura solidi su cui far crescere i microrganismi in “colonie” distinte. Inizialmente per solidificare i terreni liquidi fu utilizzata la gelatina, la quale però fonde a 35 gradi. Hesse però utilizzò l’ agar , un polisaccaride complesso. L’uso dell’agar presentava un duplice vantaggio rispetto alla gelatina, solidifica intorno ai 40 gradi ma liquefa alla temperatura di 85 gradi, offrendo così una buona stabilità termica; inoltre non viene degradato da microrganismi, salvo poche eccezioni.
1.4 Distribuzione dei microrganismi nell’ambiente
Martinus Beijerinck studiò i microrganismi del suolo e sviluppò la tecnica delle colture di arricchimento basata sull’impiego di terreni colturali di tipo selettivo. Egli scoprì che aggiungendo o togliendo certi componenti da un terreno di coltura tradizionale o incubando la coltura in diverse condizioni ambientali era possibile favorire la crescita di determinati microrganismi escludendo quella di altri. Sergej Vinogradskij fu uno dei primi ad isolare in coltura pura batteri capaci di ossidare l’ammonio a nitrato; studiò inoltre il consumo d’idrogeno solforato da parte dei batteri zolfo-ossidanti direttamente nel loro habitat naturale. Introdusse inoltre il concetto di chemiolitotrofia per descrivere i batteri capaci di crescere usando esclusivamente composti inorganici come fonte di carbonio e di energia. La prima dimostrazione diretta del ruolo dei microrganismi come agenti di malattia deriva dagli studi condotti sul carbonchio da Robert Koch , isolò infatti l’agente responsabile della tubercolosi, della colera. Sulla base dei suoi esperimenti formulò dei criteri generali, noti come postulati di Koch , per provare che un determinato microrganismo è responsabile di una specifica malattia. Man mano che aumentavano le conoscenze sulle malattie di origine microbica, si svilupparono anche farmaci per la cura di queste malattie dando inizio all’era degli antibiotici. Per colpa di un impiego sconsiderato di questi farmaci, si sono diffusi nelle popolazioni batteriche meccanismi di resistenza agli antibiotici.
1.5 Microrganismi e trasformazione della sostanza organica
Pasteur , convinse il mondo scientifico che i processi fermentativi erano il risultato dell’attività di microrganismi, e in particolare che la fermentazione alcolica era opera del lievito. Egli scoprì inoltre che la produzione indesiderata nelle bevande alcoliche di composti quali l’acido lattico e l’acido acetico era dovuta alla presenza di microrganismi di altra natura.
Altri batteri hanno forme molte più complesse: i batteri filamentosi, come gli streptomiceti, formano ife costituite da lunghe catene di cellule, che danno origine ad un micelio simile a quello dei funghi; i batteri peduncolati presentano un peduncolo che consente loro di aderire al substrato o aumentare la superficie di scambio con l’ambiente. La morfologia batterica è determinata dalla parete mureinica o sacculo, costituita da un’unica macromolecola gigante di un polisaccaride complesso (il peptidoglicano ) che avvolge la cellula.
Un elemento del citoscheletro batterico è FtsZ , una proteina essenziale per la divisione cellulare, simile alla tubulina, che al momento della divisione si aggrega formando un anello (setto) nel piano equatoriale ove recluta altre proteine della divisione. Molti batteri di forma bastoncellare contengono anche una o più proteine simili all’actina chiamate MreB , che assumono una struttura elicoidale e sono essenziali per il controllo della forma cellulare. Il batterio a forma di vibrione, contiene un terzo elemento del citoscheletro, la crescentina (CreS) , simile ai filamenti intermedi e necessaria alla curvatura della cellula.
2.2 Membrane e pareti
La cellula procariota è delimitata innanzitutto da una membrana plasmatica. Questa è protetta di norma da una parete. Inoltre molti procarioti in aggiunta alla parete possiedono ulteriori strati di rivestimento ( strato S, capsula o glicocalice ) e appendici di superficie ( pili, flagelli ). Nella parete dei batteri Gram negativi si osserva una vera e propria membrana con doppietto di lipidi e glicolipidi e con proteine associate detta membrana esterna (per cui la membrana plasmatica è anche detta membrana interna ). Nello spazio delimitato dalle due membrane, detto periplasma , si trova un sottile strato di peptidoglicano. La membrana esterna conferisce alla cellula una particolare resistenza a sostanze tossiche e la capacità di colonizzare svariati ambienti ostili. I batteri Gram positivi sono invece privi di membrana esterna e la loro parete è composta da uno spesso strato di peptidoglicano.
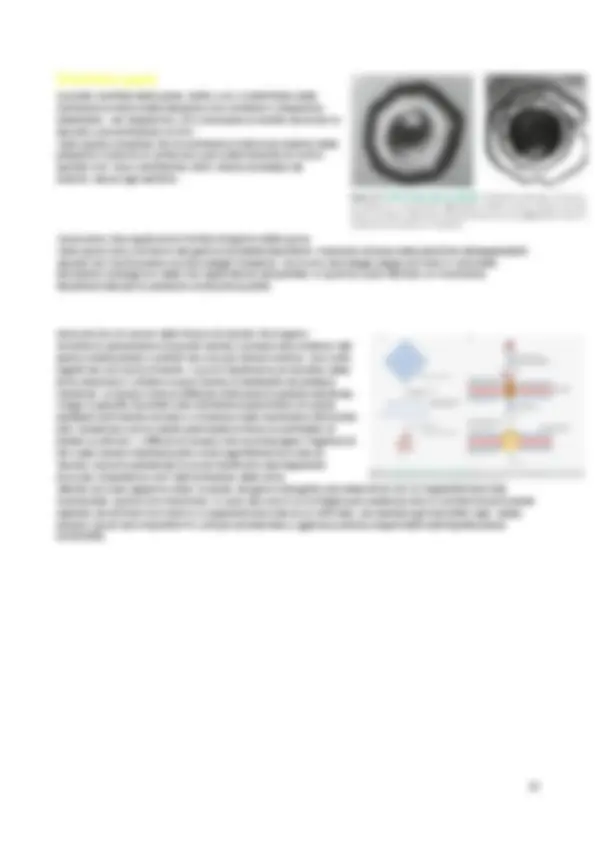
Colorazione di Gram
La colorazione di Gram , permette di distinguere i batteri in due gruppi; in base alla diversa risposta al procedimento che consiste in una colorazione con cristalvioletto e mordenzatura con ioduro di potassio, seguite da decolorazione con etanolo. La fissazione al calore, può essere fatta in vari modi= 1-Sospensione al vapore; 2-Lasciare la sospensione all’aria, aspettando che si asciughi; 3-Sospensione con una pinzetta su una fiamma (in questo caso non ci interessa la forma, ma la colorazione); Il cristalvioletto interagisce con lo ioduro in soluzione acquosa formando un complesso insolubile che precipita nel citoplasma della cellula. I batteri che dopo il lavaggio con alcol trattengono il complesso cristalvioletto-ioduro appaiono blu-viola al microscopio e vengono detti Gram positivi. In altri batteri, invece, detti Gram negativi, i ripetuti lavaggi con etanolo rimuovono il precipitato dalle cellule che così si decolorano. Per visualizzare al microscopio i batteri Gram negativi dopo la decolorazione con etanolo si ricorre a una successiva colorazione con safranina , colora di rosso-rosa i Gram negativi. È stato chiarito che la risposta alla colorazione di Gram dipende dallo spessore della parete di mureina. La spessa parete dei positivi riesce a trattenere il complesso cristalvioletto-ioduro precipitato nel citoplasma. Nei negativi, al contrario, la membrana esterna è distrutta dall’etanolo e il sottilo strato di mureina non è
sufficiente per trattenere il precipitato colorato. I procarioti appartenenti al dominio degli Archea mancano del petidoglicano. Oggi si preferisce indicare i batteri Gram negativi e Gram positivi rispettivamente con didermi (dotati di due membrane e più evoluti) e monodermi (dotati di una sola membrana e più antichi).
Membrana plasmatica
La membrana plasmatica delimita il contenuto cellulare in uno spazio definito e separa la cellula dal “resto del mondo”. La membrana plasmatica è spessa circa 7nm. Le cellule sono in grado non soltanto di acquisire nutrienti, eliminare sostanze di rifiuto o secernere molecole specifiche per le più diverse funzioni, ma anche di mantenere costante e in uno stato altamente organizzato il loro contenuto indipendentemente da cambiamenti delle condizioni esterne (omeostasi). La membrana plasmatica oltre che fungere da barriera di permeabilità selettiva, è la sede di importanti processi metabolici. Inoltre grazie alla presenza di recettori specifici, la membrana plasmatica, è una struttura fondamentale per la trasduzione di segnali. Le membrane di eucarioti e procarioti sono costituite da un doppio foglietto fosfolipidico , ma differiscono per composizione lipidica e tipologia di proteine. La membrana plasmatica di Bacteria ed Eukarya è costituita tipicamente da fosfolipidi in cui la testa polare è rappresentata da una molecola di D-glicerolo 3-fosfato e la coda da due molecole di acidi grassi non ramificati esterificati al C1 e C2 dello scheletro del D-glicerolo. Al fosfato esterificato in C3 possono essere legati gruppi funzionali. La struttura generale della membrana plasmatica è stabilizzata da interazioni idrofobe e da cationi divalenti quali Mg2+ e Ca2+, che stabiliscono interazioni ioniche con le cariche negative dei fosfolipidi. Gli acidi grassi presenti nei fosfolipidi delle membrane batteriche sono in maggioranza saturi, la presenza di catene lipidiche con doppio legame ne diminuisce la viscosità. Nelle membrane citoplasmatiche eucariote si ritrovano anche steroli , lipidi caratterizzati da un nucleo a 4 anelli aromatici, come il colesterolo, che sono generalmente assenti nei batteri. Gli steroli sono molecole planari rigide e la loro presenza nella membrana costituisce a stabilizzarne la struttura e a renderla meno flessibile. Nella maggior parte dei batteri sono invece presenti gli opanoidi , molecole simili agli steroli. Anche la presenza di proteine integrali di membrana (insolubili in soluzioni acquose), conferisce alla membrana numerose proprietà funzionali.
La membrana plasmatica negli Archea : le differenze con le altre membrane riguardano la chiralità del glicerolo (L-glicerolo anziché D-glicerolo), il tipo di legame tra glicerolo e catena alifatica (etere anziché estere) e il tipo di catena alifatica (satura ramificata anziché lineare insatura). Infatti negli Archea il fosfato è esterificato con il glicerolo in C1; le catene laterali alifatiche non sono costituite da acidi grassi lineari insaturi esterificati al glicerolo, bensì da catene isoprenoidi. Inoltre i lipidi degli Archea possono essere dieteri di glicerolo (costituiti da due catene alifatiche tipicamente a 20 atomi di C) o tetraeteri di glicerolo (costituiti da due catene alifatiche tipicamente a 40 atomi di C). Questo tipo di membrana si ritrova infatti frequentemente negli Archea ipertermofili, che crescono a temperature estremamente elevate. I lipidi dei Bacteria ad alte temperature divengono più permeabili all’acqua. I lipidi degli Archea formano un’impalcatura rigida che mantiene un’elevata impermeabilità anche alle alte temperature grazie alla minor facilità di movimento, si trovano infatti in ambienti estremi e quindi serve una membrana più stabile. La presenza di anelli ciclopentanici, a causa del loro ingombro sterico, contribuisce a ridurre la mobilità delle catene isoprenoidi.
cellule bersaglio; qui possono formare pori che danneggiano la membrana plasmatica, oppure essere traslocate nel citoplasma. Le batteriocine hanno una azione simile a quella degli antibiotici ma a differenza di questi ultimi agiscono in modo “selettivo”, tollerano lo stress, le oscillazioni di temperatura e di pH, sono quindi di facile impiego e innocue al consumo umano. Tipologia di batteriocine: 1- Colicine : proteine di peso molecolare > 40 kDa; prodotte da escherichia coli. 2- Microcine: prodotte da batteri Gram negativi, peso molecolare< 10 kDa. 3- Batteriocini : prodotte da batteri Gram positivi, peso molecolare da <5 a >30 kDa.
Polimixine: le polimixine costituiscono una famiglia di antibiotici lipopeptidici a sintesi non
ribosomiale. Queste molecole sono formate da un decapeptide cationico e da una coda lipofila. Per la loro struttura le polimixine agiscono come detergenti cationici, interagendo con i fosfolipidi della membrana plasmatica e, nei batteri Gram negativi, con il lipopolisaccaride della membrana esterna. In tal modo distruggono l’integrità e la permeabilità selettiva di queste membrane, alterando così l’equilibrio elettro-osmotico della cellula e causando di conseguenza il blocco delle sue funzioni fino anche alla lisi cellulare. La polimixina B e la polimixina E sono i due antibiotici impiegati nella pratica clinica per il trattamento di infezioni causate da batteri Gram negativi multiresistenti. Un meccanismo di resistenza consiste nel ridurre la carica negativa del lipopolisaccaride, il principale bersaglio nei batteri Gram negativi, riducendo così l’affinità per questo antibiotico. In alcuni casi particolari la resistenza è dovuta addirittura all’inibizione della sintesi del lipopolisaccaride (e quindi all’assenza della cellula bersaglio). La polimixina B viene dal polymixa, mentre la polimixina E (colistina) proviene dal bacillus colistinus.
Daptomicina: la doptimicina è attiva nei confronti dei batteri Gram positivi.
La daptomicina, in presenza di ioni calcio, si lega alla membrana plasmatica ove inserisce la sua coda lipofila; quindi oligomerizza alterando così la conformazione della membrana, creando pori e causando una rapida depolarizzazione con perdita di ioni potassio e conseguente rapida morte cellulare. Meccanismi di resistenza: alterazione dell’espressione e a un accumulo di mutazioni in diversi geni che portano a un cambiamento delle cariche elettriche di superficie, impedendo per repulsione elettrostatica il legame tra l’antibiotico e il bersaglio.
Polieni: i polieni sono costituiti da una lunga catena carboniosa ciclizzata tramite legame estere
contenente da una parte una serie di doppi legami coniugati e dall’altra numerosi gruppi ossidirilici. Queste molecole formano complessi con gli steroli di membrana, e sono pertanto attive solo nei confronti di cellule eucariote. Vengono usate per il trattamento di infezioni fungine ed esplicano azione fungistatica o fungicida. I batteri non sono sensibili ai polieni poiché la loro membrana cellulare è sprovvista di steroli.
2.3 Parete batterica
La maggior parte dei batteri, esternamente alla membrana plasmatica possiede una parete , un sacculo di mureina che avvolge completamente la cellula formando un esoscheletro cellulare. Il sacculo conferisce alla cellula rigidità e forma, ne protegge l’integrità contrastando la pressione osmotica interna ed è capace di restringere ed espandere il suo volume fino a tre volte. Questa struttura cellulare è essenzialmente costituita da peptidoglicano. Gram positivi = unico spesso strato di peptidoglicano; Gram negativi = struttura pluristratificata e complessa; Il peptidoglicano ha una struttura reticolare formata da catene glicaniche costituite da due aminozuccheri alternati, l’N-acetil-glucosamina ( NAG ) e l’ acido N-acetil-muramico ( NAM ). Al NAM è legato un tetrapeptide tramite il quale le catene glicaniche si legano tra loro covalentemente formando un’enorme molecola che avvolge la cellula. La parete mureinica può essere indebolita o rimossa mediante trattamento con antibiotici che inibiscono la sintesi del peptidoglicano (es. lisozima). Questi trattamenti possono portare alla lisi osmotica delle cellule se si trovano in ambiente ipotonico ; tuttavia se si trovano in ambiente isotonico si possono ottenere cellule sferoidali prive di parete dette protoplasti. Se i protoplasti sono trasferiti in ambiente ipotonico, questi si rigonfiano per ingresso del solvente fino a lisare. In ambiente ipertonico, l’ambiente richiama acqua dalla cellula, quindi la membrana non è più tesa e tende a collassare provocando la plasmolisi. Grazie alla parete però ciò non avviene, le cellule tendono ad opporsi in ogni modo alla plasmolisi; quello più fisico consiste nell’avere delle molecole, come gli acidi lipoteicoici , che ancorano la membrana alla parete. Quindi nelle soluzioni ipotoniche è garantito il turgore cellulare e nelle soluzioni ipertoniche invece l’integrità contro la plasmolisi.
Funzioni parete:
1. Turgore cellulare: la parete favorisce un equilibrio tra le pressione dentro la cellula e quella fuori (pressione osmotica-pressione idrostatica) 2. Mantiene e dà la forma ; 3. Contribuisce all’isolamento dall’esterno con la membrana ; 4. Favorisce la difesa da altri microrganismi ; 5. Fa parte del sistema di attacco ad altre cellule.
I filamenti di glicano adiacenti (che costituiscono lo scheletro glicosidico della mureina) sono interconnessi da legami tra i tetrapeptidi. Questi legami transpeptidici si formano, nella maggior parte dei casi, tra il gruppo carbossilico della D-alanina in posizione 4 di un tetrapeptide e il gruppo aminico del diaminoacido in posizione 3 di un tetrapeptide di un filamento glicanico. Tale legame è definito legame crociato 3-4 diretto , si trova principalmente nei Gram negativi. Nei Gram positivi in genere il legame è mediato da un ponte interpeptidico. La terza dimensione è formata da 5 glicine nei gram positivi. Mentre nei gram negativi, manca la terza dimensione, perché il legame amminico va direttamente a legarsi con il carbossile della D-alanina.
Biosintesi
La biosintesi del peptidoglicano è un processo complesso che possiamo suddividere in 3 stadi. Nel citoplasma avviene la sintesi dei precursori ; a livello della membrana plasmatica si ha la sintesi dell’unità monomerica del filamento peptidoglicanico ; infine nello spazio esterno alla membrana avvengono le reazioni di polimerizzazione e transpeptidazione (formazione legami crociati).
1 stadio
I precursori sintetizzati nel citoplasma sono i due aminozuccheri NAM e NAG legati all’UDP a formare UDP-NAG e UDP-NAM. La biosintesi del peptidoglicano comincia nel citoplasma, dove l'amminoacido glutammina dona un gruppo amminico allo zucchero fruttosio 6-fosfato , trasformandolo in glucosamina-6-fosfato. L'acetil-Coenzima A trasferisce invece un gruppo acetile consentendo la formazione della N-acetilglucosamina-6-fosfato , che a sua volta diviene N-acetilglucosamina-1- fosfato grazie a una isomerizzazione.
La N-acetilglucosamina-1-fosfato (NAG) si unisce all'uridintrifosfato (UTP) con perdita di un pirofosfato e conseguente formazione di NAG-UDP. Il complesso NAG-UDP viene mutato dal fosfoenolpiruvato (PEP) e dall'NADPH nel cosiddetto Nucleotide di Park, composto da UDP e acido N-acetilmuramico (NAM).
Al gruppo carbossilico del NAM vengono aggiunti gli amminoacidi L-alanina, acido D-glutammico e L-lisina, che nei Gram-negativi è sostituita dall'acido mesodiaminopimelico (DAP), al terzo amminoacido verranno poi collegati amminoacidi caratteristici per ogni specie batterica. Infine vengono attaccate al tripeptide due D- alanine, formate a partire dalla L-alanina tramite la D-alanin-racemasi. Si forma così una molecola di UDP- NAM-pentapeptide. Il monomero precursore del peptidoglicano è quindi sintetizzato come glicano pentapeptide. La D-alanina in posizione 5 sarà successivamente rilasciata durante la formazione del legame transpeptidico. Il glican pentapeptide è il monomero che unisce la prima e la seconda dimensione. Viene montato fuori dalla parete. L’UDP NAM e l’UDP NAG vengono trasformati con il bactoprenolo attraverso la membrana e vengono assemblati sulla membrana insieme al glicanpentapeptide.
Parete dei batteri Gram positivi
Essa è costituita essenzialmente da uno spesso strato di mureina in cui sono inserite altre molecole polimeriche ( acidi teicoici e acidi teicuronici ) cui si aggiungono proteine di superficie. La parete si estende per uno spessore di 50nm intorno alla membrana. Gli acidi teicoici si possono distinguere in acidi teicoici di parete (polimeri polianionici) e acidi lipoteicoici. I primi sono ricchi in fosfato, e quindi responsabili della carica negativa della superficie cellulare e sono ancorati all’involucro mureinico tramite legami covalenti con residui NAM del peptidoglicano. Gli acidi teicoici costituiscono dal 30 al 60% della parete cellulare e sono ritenuti essenziali per il normale funzionamento della cellula. Gli acidi teicuronici sono invece polimeri privi di fosforo contenenti N-acetil-galattosamina e acido D-glucoronico. Questa complessa matrice polianionica, costituita da peptidoglicano, acidi teicoici e teicuronici: Definisce le caratteristiche chimico-fisiche dell’involucro Controlla il movimento degli ioni e di altre molecole, antibiotici inclusi; Regola il movimento e l’attività di proteine periplasmatiche ed extracellulari; Media l’adesione delle proteine di parete, il riconoscimento da parte di batteriofagi e le molteplici interazioni con l’ambiente esterno, inclusa la risposta immunitaria.
Nella foto gli acidi teicoci (in verde) e lipoteicoici (in rosso) sono immersi nel peptidoglicano (nero).
Sulla base delle sequenze dell’rRNA 16S, i batteri Gram positivi sono raggruppati in due phyla: il phylum dei Firmicutes o con basso contenuto G+ C nel loro DNA e il phylum degli Actinobacteria o con alto contenuto G+ C.
Antibiotici inibitori della sintesi del peptidoglicano
Il peptidoglicano è un componente unico ed essenziale della parete di quasi tutti i bacteria, non sorprende quindi che gran parte degli antibiotici interferiscano con questo sistema. Gli antibiotici che inibiscano la sintesi
del peptidoglicano agiscono a livello dei diversi stadi del complesso processo di biogenesi della parete cellulare stessa. INIBITORI DI REAZIONE DEL 1 STADIO: Fosfomicina : è un analogo strutturale del fosfoenolpiruvato e si lega covalentemente all’enzima piruviltransferasi che catalizza la reazione di condensazione del fosfoenolpiruvato con l’UDP-N-acetil- glucosamina; in tal modo inibisce la formazione di enoil-N-acetil-glucosamina, precursore dell’acido N- acetil-muramico. La resistenza verso la fosfomicina è frequente ed è associata a mutazioni in geni cromosomali. Cicloserina : impedisce il completamento della catena pentapeptidica dell’acido N-acetil-muramico, e di conseguenza la formazione di legami interpeptidici tra due filamenti di glicano.
INIBITORI DI REAZIONE DEL 2 STADIO: Bacitracina : questo antibiotico si lega all’unducaprenil pirofosfato e ne inibisce la defosforilazione a undecaprenil fosfato e di conseguenza la formazione del lipide I e la biosintesi del glicano pentapeptide.
INIBITORI DI REAZIONE DEL 3 STADIO: Antibiotici Beta-lattimici : agiscono come inibitori competitivi delle ”PBP” , enzimi che catalizzano alcune reazioni chiave nella biosintesi del peptidoglicano, in particolare: -transglicosilasi, transpeptidasi, D-alanina, peptidoglicano-endopeptidasi. Il legame dell’antibiotico agli enzimi comporta l’inibizione della funzione di quest’ultimi. Penicilline = sono un ampio gruppo di antibiotici costituiti da un nucleo legato a differenti catene laterali che ne condizionano le peculiarità farmacocinetiche e lo spettro d’azione. La penicillina G , agisce principalmente sui Gram positivi, essendo la membrana esterna dei Gram negativi scarsamente permeabile a questa molecola. Le penicilline quindi legano le principali PBP, implicate nella sintesi e nell’allungamento del peptidoglicano, nella formazione del setto di divisione e nel mantenimento della forma del microrganismo. La resistenza agli antibiotici beta-lattamici può essere ricondotta a tre meccanismi principali:
- L’impermeabilità dell’antibiotico;
- La modificazione del bersaglio;
- L’inattivazione enzimatica, a seguito quindi della produzione di enzimi come le beta-lattamasi, che idrolizzano il legame amidico dell’anello beta-lattamico Nei batteri Gram positivi le beta-lattamasi, la cui sintesi è indotta dalla presenza dell’antibiotico, sono esocellulari e sono attive verso le penicilline (penicillinasi).
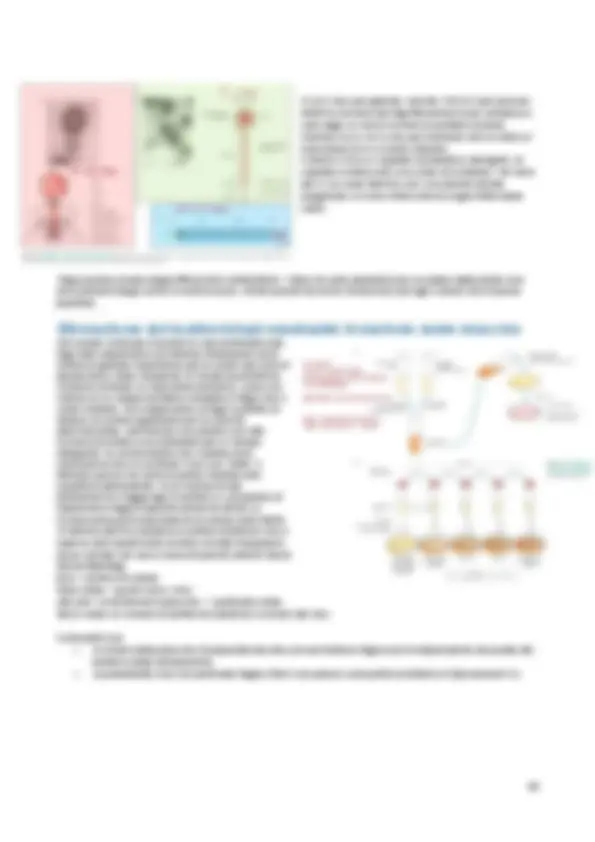
Il lipide A, costituito da un dimero di N- acetil-glucosamina è la parte lipidica che ancora il LPS alla membrana esterna. Al lipide A è legato il core polisaccaridico, che può essere suddiviso ulteriormente in core interno e core esterno: la composizione del core interno è abbastanza conservata nei diversi tipi di LPS e contiene da una a tre molecole di Kdo. Il core esterno è più variabile e contiene zuccheri a 6 o 7 atomi di C. al core saccaridico è legata la catena polisaccaridica (antigene O), costituita da lunghe catene di unità tetra- o pentasaccaridiche ripetute che si estendono verso l’esterno della cellula.
Grazie alla composizione e alla struttura del LPS la membrana esterna rappresenta un’efficace barriera di permeabilità. Nei batteri patogeni il LPS rappresenta un importante fattore di virulenza. Il LPS rappresenta anche, per gli organismi ospiti, un importante segnale della presenza batterica. Infatti la catena polisaccaridica O è in grado di indurre la produzione di anticorpi specifici nell’ospite, mentre il lipide A è in grado di attivare l’immunità innata dell’ospite.
La diffusione nel periplasma di ioni e piccole molecole è possibile grazie alla presenza di proteine canale denominate porine. La dimensione del canale può variare a seconda della porina considerata. Dal momento che la membrana esterna è priva di fonti energetiche, il passaggio di nutrienti attraverso le porine avviene secondo il gradiente di concentrazione.
Attraverso la membrana esterna passano anche molecole di grandi dimensioni, mediante macchinari multiproteici; costituiti da proteine della membrana esterna, proteine periplasmatiche e proteine della membrana interna il cui funzionamento coordinato consente il trasporto delle sostanze dall’esterno al citoplasma delle cellula o in senso opposto.
Un’altra classe di proteine associate alla membrana è rappresentata dalle liporoteine. La modificazione lipidica all’N-terminale ne consente l’ancoraggio alla membrana esterna. Le lipoproteine batteriche partecipano a numerose funzioni cellulari quali stabilizzazione delle strutture di superficie della cellula, trasporto, resistenza ad antibiotici, trasduzione del segnale.
Trasporto proteine integrali, lipoproteine e lipolisaccaride;
Le proteine della membrana esterna sintetizzate nel citoplasma possiedono una sequenza all’N-terminale che le indirizza alla traslocazione attraverso la membrana interna. I sistemi responsabili della traslocazione delle proteine attraverso la membrana plasmatica sono due: il cosiddetto traslocone Sec , che provvede al trasporto di proteine non ancora ripiegate nella struttura terziaria definitiva, e il sistema Tat , che invece trasloca proteine che sono state già ripiegate nel citoplasma. Le lipoproteine sono sintetizzate nel citoplasma come lipoproteine e possiedono una sequenza all’N terminale che le indirizza al traslocone Sec, il quale provveder al loro trasporto attraverso la membrana plasmatica. Il complesso formato dalle proteine LolCDE, un trasportatore ABC localizzato nella membrana interna, prende in carico le lipoproteine che hanno attraversato la membrana interna e, grazie all’energia derivata dall’idrolisi di ATP, le indirizza allo chaperon periplasmatico LolA. Il complesso LolA-lipoproteina interagisce con LolB, un recettore localizzato nella membrana esterna responsabile dell’inserzione della lipoproteina nella membrana esterna. La modificazione lipidica all’N-terminale e la rimozione della sequenza segnale avvengono nel periplasma. Il LPS viene sintetizzato in modo particolare; la porzione lipidica A-core è sintetizzata separatamente dalle unità polisaccaridiche che costituiscono l’antigene O. I micoplasmi sono batteri incapaci di sintetizzare il peptidoglicano e pertanto privi di parete. Le cellule dei micoplasmi, quindi sono racchiuse esclusivamente dalla membrana plasmatica. La loro membrana è lipoproteica e ricca di steroli, caso unico fra le cellule batteriche.
Varianti citologiche (lezione 4)
La maggior parte dei batteri risponde in modo univoco alla colorazione di Gram, ma alcuni gruppi di batteri filogeneticamente appartenenti ai Gram positivi si colorano in modo diverso a seconda della fase di crescita; questi batteri vengono definiti gram variabili. Molte cellule dei batteri Gram variabili di una coltura in fase avanzata di crescita risultano più fragili a livello del setto di divisione e rilasciano il colorante Gram, risultando così fenotipicamente Gram negative.
Micobatteri
La parete dei micobatteri ha una struttura abbastanza complessa che può essere divisa in due strati: Lo strato basale (o core ) costituito da peptidoglicano e arabinogalattano (un polisaccaride ramificato complesso); Una struttura più esterna bistratificata costituita da uno strato di acidi micolici (acidi grassi complessi a catena lunga, contribuiscono alla scarsa fluidità della membrana esterna) e uno di cere ; Questa parete cellulare così complessa, impermeabile e poco fluida costituisce una barriera protettiva formidabile e rende i micobatteri completamente impermeabili a molte sostanze utilizzate nella terapia medica, compresi alcuni degli antibiotici più comuni. Proprio per questo i micobatteri sono così difficili da colorare.
Colorazione di Ziehl-Neelsen
La colorazione adottata è quella di Ziehl-Neelsen. Questa colorazione differenziale prevede, in primo luogo, dopo la fissazione al calore del campione su un vetrino da microscopio, un trattamento a caldo dei batteri con una soluzione di fucsina e fenolo in etanolo ( carbolfucsina ): a temperatura elevata questo colorante è solubile nella parete dei micobatteri e può così penetrare nel citoplasma. Si procede poi con una decolorazione con alcol acido , che decolora tutti gli altri batteri presenti sul vetrino. Infine si effettua una colorazione di contrasto con blu di metilene. Sul vetrino così colorato i micobatteri appaiono come bastoncini rossi (perché non hanno perso la colorazione iniziale della fucsina), mentre altri batteri eventualmente presenti sono colorati in blu.
Colorazioni batteriche
Per facilitarne l'osservazione ed il riconoscimento, i batteri possono essere colorati, ovvero modificati con agenti chimici. Le colorazioni batteriche possono essere semplici o differenziali. Nel primo caso si procede con l'applicazione di una singola soluzione di colorante (blu di metilene, fucsina, eosina...), su un vetrino portaoggetti su cui una piccola quantità di cellule batteriche era stata distesa e fissata. Dopo la colorazione, l'eccesso di colorante viene rimosso mediante un rapido risciacquo. Consente di rilevare la morfologia e l’organizzazione cellulare, le dimensioni e la carica. Nel caso di colorazioni differenziali, vengono fatti agire due coloranti in successione, di cui il primo viene rimosso con attenzione in modo da evidenziare le varie popolazioni batteriche ed il secondo viene usato per colorare le cellule che non si sono colorate con il primo colorante. Consente di distinguere due differenti tipologie di microrganismi o 2 differenti strutture di un microrganismo A questa categoria appartengono la Colorazione di Gram e la Colorazione di Ziehl-Neelsen. Se a noi interessa osservare la morfologia delle cellule, basta utilizzare la colorazione semplice, non differenziale, la quale colora tutte le cellule. Nel caso specifico ad esempio, la safranina colora sia gram positivi, sia gram negativi. Mentre il blu di metilene , entra in tutte le cellule, ma può essere differenziale, infatti se le cellule sono vive lo buttano fuori.
Una colorazione differenziale può essere dovuta a una struttura differenziale o a qualcosa di fisiologico (solo se la cellula è viva).