Preuzmite MALIGNA CELIJA-skripta-predavanje Part1 i više Rezime u PDF od Biologija ćelije samo na Docsity!
MALIGNA CELIJA
UVOD
U medicinskoj profesiji postoji aksiom da je kancer skup od oko dve stotine različitih patoloških stanja a ne jedna bolest. Ovo je razumljivo sa tačke gledišta patologa koji se suočava se mnoštvom ćelijskih abnormalnosti, za epidemiloga koji poznaje veliki spektar heterogenih uzročnika i za onkologa-internistu koji se bori sa različitim terapijskim odgovorima na raznovrsne terapijske modalitete. Za molekularnog biologa situacija je sasvim različita: svi tipovi malignih ćelija poseduju ograničen skup zajedničkih, fundamentalnih abnormalnosti i za to mora da postoji jedinstveno racionalno objašnjenje. Sada postoji dobar razlog da se veruje da konačno postoji način da se objedine naizgled oprečna gledišta. Bez obzira kako se kancer manifestuje, suštinu fenotipa usmeravaju geni a većina, ako ne i svi, uzročnici obolenja nanose oštećenja genima ili direktno ili indirektno.. Istraživanja u oblasti molekularne biologije raka se usmeravaju uverenjem da je shvatanje patogeneze raka na molekularnom nivou preduslov za razvoj novih tipova dijagnostičkih sredstava i efikasnijih terapeutika. Ovo uverenje tek treba da se ostvari. Poslednja decenija je donela prosvetljavajuća otkrića o molekularnim osnovama ovog obolenja ali njihova primena još nije prevedena u konkretnu onkološku praksu. Izvesno je, međutim, da će do toga doći, a kada dođe, zasnivaće se na osnovnim istraživanjima koja će biti opisana. Dva pristupa problemu Ovaj tekst je, između ostalog, i svedočenje o revoluciji koja je prohujala kroz biomedicinske nauke u toku poslednje tri decenije. Revolucija se bazirala na otkrićima koja su omogućila izolovanje i umnožavanje (kloniranje) gena, njihovo sekvenciranje i kreiranje strukturnih izmena koje modifikiju njihove primanrne funkcije. Ova tehnološka unapređenja su u osnovi našeg razumevanja fundamentalnih procesa nastajanja gena odgovornih za maligni fenotip. Istraživanja odgovorna za pomenute naučne prodore se zasnivaju na dva fundamentalna koncepta. Prvo, kancer je u osnovi obolenje pojedinačnih ćelija. Odavde sledi da bi morali da razumemo većinu, ako ne i sve osobine malignih tumora, ako razumemo osobine pojedinačnih ćelijskih komponenti. Ovo uverenje je bilo u osnovi decenajama dugih istraživanja na in vitro medelima tumorigeneze. Drugo, ponašanje ćelija i kao logični nastavak, ponašanje složenih tkiva se može razumeti kao posledica aktivnosti gena. I zaista, pokazalo se da se složenost kancera može shvatiti samo preko analize gena - kancer je perverzija ćelijskog fenotipa a geni su determinate tog fenotipa. Klonalno poreklo Rudolf Virchow je maglovito shvatio naslednu prirodu raka znatno pre kraja 19- og veka. Uočio je sličnost ćelijskih komponenti kod metastaza i kod primarnih tumora. Za taj vremenski period on pravi smelu hipotezu da sve ćelije potiču od drugih ćelija. Zatim sledi još hrabriji stav. Ako su sve ćelije tumora srodne, onda je moguće da sve
potiču od zajedničkog pretka. Ovo je verovatno najranija sugestija za nešto što danas smatramo fundamentalnom osobinom većine, ako ne i svih, tumora: - oni su klonalni. Ova osobina tumora ima dalekosežne implikacije koje Virchow nije bio u situaciji da pojmi. Suština je u tome da neoplastični fenotip mora biti izuzetno stabilan jer perzistira kroz neprebrojive ćelijske deobe koje su neophodne za genezu tumora iz zajedničkog klona - jedne jedine ćelije. Drugim rečima maligni fenotip mora biti nasledan. Nameće se sumnja da abnormalnost malignih ćelija mora imati svoje ishodište u genetskoj abnormalnosti odnosno izmeni u genetskom materijalu. Iznenađuje kako su naučnici 20. veka bili spori u prihvatanju validnosti ove jednostavne dedukcije. Spoljašnji uzročnici Ako rak počinje kao pojedinačna ćelija koja prerasta u razvijeni malignitet, šta inicira ovaj smrtonosni tok događaja? Više od dva veka istraživanja su bila usmerena ka spoljašnjim uzročnicima. U ovom procesu, početkom 20. veka, iskristalisala su se dva suprostavljena koncepta. Jedan se je bio usmeren ka fizičkim i hemijskim agensima a drugi ka virusima kao primarnim uzročnicima kancera. Obe tačke gledišta su se pokazale kao izvor infomacija od vitalnog značaja za razumevanje procesa kancerogeneze. Saznanja o fizičkim i hemijskim agensima kao mogućim uzročnicima raka potiču čak iz 1761. godine kada je John Hill predložio da ušmrkavanje burmuta može da uzrokuje pojavu nazalnog kancera. Nešto kasnije Percival Pott stiče slavu svojim zapažanjima da dimnjičari imaju sklonost ka kanceru skrotuma, verovatno zbog izloženosti čađi koja nastaje pri nepotpunom sagorevanju uglja. Ova važna opažanja su bila, uglavnom, zanemarivana do kraja 19-og veka. Tek tada je nagli industrijski razvoj istakao sumnju u štetne efekte koji se javljaju u toku prerade parafinskih ulja, karakteristične prašine u ugljenokopima, u preradi arsena i anilinskih boja. Istovremeno, postalo je očigledno da novootkriveni X-zraci i UV zračenje mogu biti kancerogeni. Epidemiološka istraživanja kod specifičnih humanih populacija su uskoro potvrđana eksperimentima. Pioniri, u ovoj oblasti istraživanja, su bili Clunet koji je 1908 dokazao da X-zraci mogu indukovati rak kože kod pacova, i Yamagiwa, koji je 1915 indukovao rak kože na ušima zečeva i leđima miševa aplikacijom čađi dobijene nepotpunim sagorevanjem uglja. Ovo je bio početak obimnih istraživanja na širokom frontu usmerenih ka identifikaciji kancerogenih hemijsko-fizičkih agenasa i mehanizma njihovog delovanja. Drugi pristup problemu je bio usmeren ka virusima kao primarnim uzročniku raka. Ova istraživanja su bila pod velikim uticajem uspešniha o infektivnom poreklu bolesti baziranih na radovima Roberta Koch-a u Nemačkoj i Lous Paster-a u Francuskoj krajem 19-og veka. Njihov spektakularni doprinos medicini je podsticao mišljenje da su sva obolenja mikrobiološkog porekla. Otkriće virusa u periodu 1898 do 1900 je dodatno podstaklo onkološka istraživanja ka traženju virusnog uzročnika malignih obolenja. Prvu ozbiljnu vezu između malignih obolenja i virusa je uspostavio Peyton Rous nakon
- On je opisao virus koji može da indukuje sarkome kod pilića i da dalje prenosi tu patologiju na način tipičan za infektivne agense. Prvorazredna je ironija da Rous napušta svoja istraživanja jer ne uspeva da izoluje sličan, ili bilo kakav, infektivan materijal iz eksperimentalnih tumora glodara. Njegov rezon je bio da ne vredi slediti izučavanje fenomena koji nema opšti karakter.
Gensko obolenje somatske ćelije Istraživanja u oblasti onkogenih virusa su pomerila fokus u razmišljanjima o poreklu raka - u prvom planu su geni kao glavni akteri u procesu kancerogeneze. Hemijski kancerogeni nisu predstavljali logički problem u ovakvoj šemi obzirom na njihovu reaktivnost u odnosu na DNK molekul. Ovako iskristalisani problem razmatraju dve, naizged, suprostavljene strategije. Prva od njih sugeriše da je kancerogeni agens usmeren ka izmeni ekspresije gena u smislu selektivne indukcije ili supresije. Za ovaj proces se smatralo da predstavlja aberantnu formu razvića ili diferencijacije. Suprostavljeni stav predlaže da su kancerogeni mutageni koji proizvode oštećenja u genima, direktno ili indirektno, tako da menjaju ili eliminišu njihovu funkciju. U oba slučaja konačan ishod je isti – genetska abnormalnost. Razlika je samo semantičke prirode i zavisi od definicije gena, koja može i ne mora, da pod ovim pojmom da podrazumeva i kodirajuće sekvence i elemente DNK koji kontrilšu ekspresiju ovih sekvenci. Prvi čvršći oslonac teoriji o genetskoj osnovi raka potiče iz 1928. godine, manje od 20 godina nakon otkrića kancerogenih efekata X-zraka, kada je H. J. Muller otkrio da oni mogu biti mutageni na modelu vinske mušice. Iako on nije bio posebno zainteresovan za kancerogenezu, njegova istraživanja su postala osnov svim savremenim modelima procesa onkogeneze. U toku naredne dekade, pokazano da je hemijski kancerogen 3- metilholantren i mutagen kod eksperimentalnih miševa. Ovaj rezultat je bio osnova za formalizovanje hipoteze da svi (spoljašnji) kancerogeni mogu biti mutageni i da ova njihova osobina predstvalja suštinu njihove kancerogenosti. Traganje za potvrdom ove hipoteze se suočilo sa dve komplikacije. Prvo, neki poznati kancerogeni su bili čudno hemijski inertni pa je dovedena u pitanje njihova biološka aktivnost. Prividni paradoks je razrešen definisanjem metaboličkih puteva detoksikacije koji generišu hemijski reaktivne intermedijere koji su direktni kancerogeni. Druga komplikacija je bio problem da se identifikuje ciljni molekul, ili deo molekula, koji se mutagenezom modifikuje u procesu kancerizacije. Razrešenje su obezbedili eksperimenti sa onkogenim virusima. Hipoteza o kancerogenu kao mutgenu je dobila snažnu podršku od strane Bruce Ames-a koji je razvio pogodne testove za određivanje mutagenih osobina hemikalija. Glavno obeležje ovih testova je da se rezultati dobijaju na ćelijama u kulturi i ne zahtevaju kompletne organizme i omogućavaju praktičnu analizu velikog broja jedinjenja. Nakon analize širokog spektra suspektnih supstanci došlo se do očekivanog nalaza – različita hemijska jedinjenja imaju izuzetno različite mutagene potencijale pri čemu faktor razlike može biti i 10 6. Još je važnije da snažni mutageni predstavljaju i potentne kancerogene kada se testiraju na eksperimentalnim životinjama. Utvrđeno je, takođe, da važi i obrnuta proporcija. Iako postoje važni izuzetci, utvrđeno je da sposobnost hemikalija da indukuju rak jasno zavisi od njihove sposobnosti da indukuju mutacije. Sasvim različit eksperimentalni pristup je dodatno podržao hipotezu da genetski aparat ne funkcioniše normalno u procesu kancerogeneze. U pitanju je citogenetika koja jasno dokumentuje da maligne ćelije relativno često nose abnormalne hromozome. Ovu problematiku je otvorio Theodor Boveri još 1914 sa hipotezom da kancer uzrokuje ili višak ili nedostatak hromozoma u normalnom kariotipu. Hipoteza je ostala samo zanimljiva pretpostavka sve do 1960 - te godine i otkrića Filadelfija hromozoma u ćelijama hronične mijeloidne leukemije. Ovo otkriće je bukvalno bilo uvod u "poplavu"
sličnih citogenetskih rezultata. U ovom trenutku, preko 100 različitih hromozoskih translokacija specifično korelira sa malignim poremećajima. Iako je ovo direktan dokaz da genetska oštećenja učestvuju u patogenezi ovog obolenja, citogenetika se pokazala kao suviše grubo oruđe za precizno dešifrovanje prirode oštećenja. Model " mutagen - kancerogen " jasno ukazuje na prisustvo mutiranih gena u svakom ili većini malignih tumora. Ovi geni moraju biti izmenjene verzije postojećih ćelijskih gena koji su oštećeni ili pod dejstvom mutagena ili nastaju kao posledica grešaka koje su inheretno svojstvo genetičkih i biohemijskih procesa u ćeliji. Ovakav koncept je premostio prividne razlike između hemijsko-fizičke i virusne kancerogeneze. Otvorena je moguđnost da su mutantni geni malignih ćelija homolozi funkcionalnih gena onkogenih virusa. Ove mogućnosti su definisale put, ka do sada, neizmenjenoj definiciji raka: stečeno genetičko obolenje somastke ćelije. Sasvim je opravdan akcenat na terminu "somatska ćelija", jer nasledne forme (familijarni kancer sindromi) i danas predstavljaju raritet odgovoran za manje od 5% klinički relevatnih slučajeva. Fizičko-hemijski kancerogeni deluju tako što modifikuju normalne gene dok onkogeni virusi mogu da indukuju mutagenezu, da transdukuju modifikovane gene domaćina ili da nose sopstvene onkogene koji nemaju homologiju sa genima domaćina. Proto-onkogeni Kada je Peyton Rous saopštio otkriće o sarkoma virusu kod eksperimentalnih pilića, napravio je ekstrapolaciju i sugerisao da virusi mogu biti odgovorni za sve maligne poremećaje. Ova ektrapolacija je dočekana sa nevericom i podsmehom i to je dugo trajalo. Raus je morao da čeka 50 godina na satisfakciju (Nobelovu nagradu), a kada je konačno i došla, imala je i očekivanu i neočekivanu formu. Kao što je Raus predvideo neki humani tumori su zaista uzrokavani virusima. Međutim, on nije predvideo dodatne posledice svog istrživačkog rada: “Njegov” virus je obezbedio otvoranje puta ka razumevanju genetske osnove svih maligniteta bez obzira na njihovo poreklo. Prvi korak je bio racionalizacija tumirigeneze indukovane Rous sarkoma virusom
- proces zavisi samo od jednog virusnog gena nazvanog src (asocijacija na vrstu tumora koje indukuje – sarkome). Samo prisustvo ovog gena u genomu virusa je bilo više nego čudno. On niti doprinosi replikaciji virusa niti mu obezbeđuje neku uočljivu selektivnu prednost. Zagonetka je razrešena otkrićem da ovaj gen i nije “zakoniti” virusni gen već “slepi putnik” pri čemu virus transdukuje gen ćelije domaćina u nešto izmenjenoj formi. Vreme je pokazalo da transdukcija src gena predstavlja samo vrh ledenog brega u onkogenetici. Otkriveno je više desetina retrovirusnih onkogena izvedenih iz genoma normalnih ćelija a ishodni geni su osvojili generički ternin proto-onkogeni. Konverzija proto-onkogena u virusni onkogen se može objasniti na nakoliko načina. Na primer, virus može intezivirati gensku ekspresiju pogrešnog gena, u porgešnom tkivu u pogrešno vreme. Takođe, moguće je da se kodirajući domen transdukovanog gena mutira u procesu ilegitimne rekombinacije i transdukcije i da njegova izmenjena struktura bude osnova aberantne funkcije i malignog potencijala. Ako su proto-onkogeni smrtonosni “slepi putnici” u retrovirusnom genomu, moguće je da sličan proces maligne konverzije postoji u ćelijama i bez učešća virusa. Transfekcija ćelija u kolturi dokumentovala je da humani tumori bez virusne etiologije
objašnjava mnoge oblike naslednih predispozicija za rak. Ovo nasleđivanje uključuje i zbunjujuću kontraverzu. Iako su nasledne mutacije recesivne na ćelijskom nivou, njihov obrazac nasleđivanja, u oviru porodice, je jasno dominantan. Skriveni paradoks proizilazi iz specifične kombinacije okolnosti: Somatska ćelija koja već nosi nesleđeni defekt u jednom alelu mora da pretrpi samo još jednu izmenu u drugom nemutiranom alelu da bi došlo do pune ekspresije posledica oštećenja u recesivnom genu. Broj ćelija u većini tkiva je obično dovoljno veliki tako da je verovatnoća da dođe do drugog neophodnog mutagenog događaja sasvim izvesna bar za jednu od ćelija. To je sasvim dovoljno za iniciranje malignog fenotipa. Tako, klonalno poreklo kancera omogućava oblik nasleđivanja koji je praktično jedinstven kod kongenitalnih obolenja. Drugo, ako se na onkogene može gledati kao na faktore ubrzanja ćelijske proliferacije, tomor supresor geni su faktori usporavanja ovog procesa. Ovde leži objašnjenje za razliku između ove dve klase gena. Smrtonosni ishod prebrze vožnje automobilom može biti zaglavljena pedala za gas (dominantnost onkogena) i isti ishod mogu imati dva uzastopna kvara na kočionom sistemu (recesisvnost tumor supresor gena). Model postaje složeniji Otkriće da samo jedan onkogen može da transformiše ćelije u kulturi i da indukuje tumore u eksperimentalnim životinjama je bilo ogromno unapređenje u oblasti eksperimentalne kancerogeneze. Ono što je nekad izgledalo beznadežno složeno je sada redukovano na aktivnost samo jednog gena. Kao što to obično biva, redukcionizam se pokazao kao opasno uopštavanje. Anali istraživanja u oblasti kancerogeneze su obeleženi nizom podataka koji daju razlog da se ovom procesu pripiše višefazni karakter. I, kao što se moglo očekivati, bila je zabluda verovati da samo jedan onkogen ili tumor supresor gen može biti dovoljan za genezu potpuno razvojeng malignog fenotipa u humanim patološkim stanjima. Humane ćelije su veoma dobro opremljene za korekciju grešaka i oštećenja u molekulu DNK. Takođe, organizam ima mehanizme za apoptotsku eliminaciju ćelija u kojima ova korekcija nije bila uspešna. Ergo, pojava mutantnih ćelija u organizmu mora biti izuzetno redak događaj. Bez obzira kako se proceni brzina pojave mutacija u humanom genomu, ćelije ne bi smele da akumuliraju dovoljno genetskih oštećenja za genezu neoplazije u toku prosečne životne dobi. Čini se, da je ova dilema konačno razrešena: u toku kancerogeneze, nastala premaligna ćelija je osetljivija u odnosu na dalju mutagenezu od normalne ćelije. Postoje bar dva objašnjenja za ovo povećanje mutabilnosti. Prvo, moguće je da geni odgovorni za korekciju grešaka pretrpe oštećenje u ranim fazama kancerogeneze i na taj način uvećavaju verovatnoću pojave novih mutacija. Drugo, ćelije su programirane da izvrše samoubistvo ukoliko oštećenje u molekulu DNK ostane nekorigovano, pa je moguće da geni koji nose informaciju za ovaj program, takođe, budu oštećeni u početnim fazama procesa. Na jedan ili drugi način, mehanizmi obezbeđenja bivaju rano onemogućeni i to olakšava preživljavanje mutantnim ćelijama i transmisiju potencijalno malignih lezija na ćelije potomke. Sociološki aspekti problema Istraživači u svakoj oblasti nauke imaju narcisoidnu sklonost da svoju delatnost smatraju posebno značajnom. Svesni te zamke autor ovog teksta će probati upravo
najopasnije – da ubedi čitaoca i slušaoca da molekularna biologija maligne ćelije ima poseban značaj u aktuelnom civilizacijskom trenutku. U tom smislu, pretenzije zaista nisu male i idu preko psiholoških implikacija, političko-ekonomskih efekata, do surove strategije evolucije koje dotiče neke aspekte filozofije i religije. Ozbiljnost problemu, na samom početku razmišljanja, nameće činjenica da, zavisno od matematičke preciznosti obrade epidemioloških podataka, svaki četvrti ili svaki peti čitaoc ovog teksta će prerano završiti svoj život zbog nekog oblika malignog obolenja. Uprkos činjenici da je situacija još gora u analizi statističkih rezultata o posledicama kardiovaskularnih bolesti, psihološki efekat, kada je rak u pitanju, ima nezavidnu vodeću poziciju. Veoma je gorka rasprostranjena polu anegdota “došlo je vreme da je luksuz umreti od infarkta miokarda” verovatno zbog prorode agonije koja prati terminalne faze malignih bolesti. Zbog toga što još uvek značajno veliki deo tehnološki razvijenog sveta umire u toj agoniji, još veći deo živi u strahu od te agonije. Situaciju samo pogoršava činjenica da savremeni amerikanac ima iste šanse da se izleči od malignog poremećaja kao i njegov deda. Kada se uzmu u obzir ogromni intelektualni i ekonomski potencijali koji su angažovani u borbi protiv raka nameće se zaključak da su istraživanja u ovoj oblasti najskuplji, neuspešni, civilni projekat u istoriji. Veoma je rasprostranjena zabluda da je rak bolest uzrokovana tehnološkim razvojem u 20-om veku. Ovaj pogrešan stav se često argumentuje podatkom da je kancer bio četrnaesti po redu uzročnik smrti sredinom 19-og veka, kada se počelo sa ozbiljnim odnosom prema medicinskoj dokumentaciji. Sada je na nezavidno visokom drugom mestu iza kardio- i cerebrovaskularnih poremećaja. Situaciju samo dramatizuje činjenica da ovaj uspon, na skali uzročnika smrti, pokazuje ubedljivu korelaciju sa poboljšanjem opšteg životnog standarda ljudi i krupnim napretkom u medicinskoj nauci i zdravstvenoj zaštiti. Prividni paradoks je lako razrešiti kada se pažljivo izračuna efekat eliminacije infektivnih bolesti i bolesti ranog detinjstva, kao uzročnika smrti. Efekat ima dve važne posledice: (1) Nekada najvažniji uzročnici smrti sada ne predstavljaju ozbiljniji zdravstveni problem pa je normalno pomeranje na skali uzročnika. (2) Prosečni životni vek je sa 37 produžen na 73 godine u toku poslednjih 160 godina. Ovakva situacija je favorizovala bolesti odmakle životne dobi, gde naravno spadaju i maligna obolenja. Uslovno rečeno, najsnažniji kancerogen je starosna dob jer učestalost obolenja eksponencijalno raste kao funkcija ovog “kancerogena”. Navedene čenjenice su definitivno tačne i ne treba da zbunjuje, takođe je tačna, činjenica da neka retka zanimanja i nezdrave navike, koje su posledica tehnološkog razvoja, zaista uvećavaju rizik od pojave ove bolesti. Krajem 1971. godine, predsednik Ričard Nikson je potpisao akt o borbi protiv raka koji je, na prvom mestu, podrazumevao visok nivo finasiranja aktivnosti u ovoj oblasti. Pokrenuta je moćna naučna mašina i obimne kliničke aktivnosti. Usledili su impresivni rezultati. U oblasti relevantnih naučnih aktivnosti dominiraju dostignuća na polju molekularne onkogenetike. U onkologiji, kliničaka praksa beleži upečatljiva smanjenja smrtnosti kod nekih oblika kancera, na primer, Hodgkin-ova bolest, Burkitt-ov limfom, testikularni kancer i različiti oblici pedijatrijskih maligniteta. Svi ovi uspesi su u senci činjenice koja je, u najmanu ruku, depresivna: U periodu 1973 do 1998, poslednja godina za koju postoje potpuni podaci, smrtnost kao posledica kancera je povećana za 6,3%. U
NORMALNE ĆELIJE I MALIGNE ĆELIJE
U ovom poglavlju opisuju se razlike između malignih ćelija i njihovih normalnih prethodnika i diskutuje zanačaj ovih razlika za mehanizam geneze tumora. Saznanja iz ove oblasti potiču, uglavnom, iz eksperimenata na ćelijama u kulturi. To je razlog da se uvodni deo poglavlja posveti toj oblasti. Zatim se razmatraju osobine autonomne proliferativne sposobnosti, genetske nestabilnosti, abnormalnosti u diferencijaciji i kontroli ćelijskog ciklusa. Na kraju se opisuje proces varijabilnosti i selekcije koji su odgovorni za progresiju tumora – proces evolucije malignih ćelija u smeru stalnog uvećanja malignog potencijala. Težište će biti na osobinama i načinu ponašanja malignih ćelija, a genetske izmene u onkogenima i tumor supresor genima, koji su u osnovi ovih pojava, će biti samo površno opisane jer njihov detaljni prikaz sledi u većini narednih poglavlja. Ćelije u kulturi Geneza kancera kod ljudi je složen, višefazni proces. Na ćelijskom nivou, kancer se razvija sporo, jer je svaki pojedinačni korak događaj niske verovatnoće. Čak i kod životinja izloženih velikim dozama kancerogena, latentni period traje više meseci. Sporost progresije kancera je srećna okolnost za pacijente, ali veoma otežava eksperimentalne pristupe izučavanju ove bolesti. Eksperimentalna analiza zahteva da proces maligne transformacije bude proizvoljno čest i reproducibilan događaj tako da se mogu primeniti tehnike molekularne biologije. Dodatna otežavajuća okolnost u izučavanju spontanog kancera je problem identifikacije normalne ćelije od koje potiču maligne ćelije. Maligne ćelije mogu biti izvedene iz male populacije proliferišućih matičnih ćelija. Često, maligne ćelije gube markere diferencijacije tako da je gotovo nemoguće utvrditi iz kog tkiva potiču. Takođe, veoma je teško primeniti tehnike molekularne biologije za izučavanje pojedinačnih malignih ćelija, koje su u okvirima tumora u progresiji u organizmu koji ga nosi. Ovo su razlozi koji ćelije u kulturi, čine nezamenljivom tehnikom u eksperimentalnoj onkologiji. Pod određenim uslovima, normalne ćelije u kulturi mogu biti izmenjene u tumorigene ćelije koje izrastaju u maligni tumor, ako se implantiraju odgovarajućoj eksperimentalnoj životinji. Ovaj proces se naziva maligna transformacija, ili jednostavno transformacija. Kod ćelija u kulturi najvažnije je to što potomstvo jedne jedine ćelije može da da novu populaciju ćelija, odnosno može se klonirati. To znači da ako se jedna ćelija genetski izmeni kancerogenim agensom, njeni potomci se mogu umnožiti do dovoljno velike kulture koja obezbeđuje dovoljno materijala za biohemijske eksperimente. Tumori,
takođe, nastaju iz pojedinačnih ćelija pa geneza ćelijskih kultura iz pojedinačne transformisane ćelije predstavlja adekvatan model tumorigeneze in vivo. Ćelijske kulture su pravljene disocijacijom tkiva u pojedinačne ćelije proteolitičkim tretmanom. Tako dobijene kulture se nazivaju primarnim kulturama. Najlakše se uzgajaju fibroblasti koji brzo počinju sa proliferacijom. U normalnom okruženju, u tkivima, fibroblasti sintetišu i organizuju ekstraćelijski matriks koji predstavlja složenu smesu glikoproteina u ugljenihhidrata koja se deponuje oko ćelije i ima funkciju mehičkog oslonca za tkiva. Kada se fibroblasti odvoje od ovog matriksa, oni se prvo pričvste za staklenu ili plastičnu površinu suda u kome se uzgajaju. Ubrzo, zatim, oni sintetišu ekstraćelijski matriks koji se, takođe, pričvršćuje za površinu suda za kulturu. Prolifaracija fibroblasta formira sloj ćelija koji pokriva površinu suda za kulturu. Ovaj sloj se često naziva monosloj , što nije najsrećniji izraz jer je često u pitanju višećelijski tepih. Ovakav monosloj se može ponovo disosovati proteolitičkim tretmanom i proparigati (pasažirati) u novu kulturu. Karcinomi (maligni tumori epitelnih ćelija ektodermalnog ili endodermalnog porekla) su mnogo češći nego sarkomi (tumori mezodermalnih ili mezenhimalnih ćelija). Zbog toga bi kulture epitelnih ćelija bile veoma korisne. Međutim, nasuprot fibroblastima, primarne epitelne ćelije teško rastu iz pojedinačnih ćelija. Vijabilnost i proliferacija pojedinih tipova epitelnih ćelija se može poboljšati uzgajanjem na sloju kolagena, koji omoguva ćelijama da uspostave okruženje slično in vivo okruženju (2). U ograničenom broju slučajeva, bilo je moguće transformisati epitelne ćelije, na primer, epitelne ćelije mlečne žlezde miša. Hematopoezne ćelije kostne srži se mogu uzgajati i koristiti, kao modeli, u izučavanju leukemija. Hematopoezne ćelije su izvedene iz pluripotentnih matičnih ćelija koje proliferišu u kostnoj srži i predstavljaju prekursore limfoidnih, mijeloidnih, megakariocitnih i eritroidnih linija. Pluripotentne matične ćelije diferentuju u predodređene (committed) progenitorske ćelije, koje se umnožavaju i dalje diferentuju u kostnoj srži ili, u slučaju limfoidnih ćelija, u perifernim limfoidnim organima. Ove ćelije su uspešno kultivisane, ali je većina eksperimenata koristila predodređene pluripotentne ćelije višeg stepena diferencijacije koje se mogu uzgajati u suspenziji u polučvrstom medijumu. U ovakvim kulturama, progenitori prođu kroz nekoliko deoba i zatim diferentuju, dajući kolonije diferenciranih ćelija. Zato se ove predodređene progenitorske ćelije ponekad označavaju kao jedinice formiranja kolonija (CFU, colony forming units). Ćelije dobijene disocijacijom normalnih tkiva se nazivaju primarnim kulturama. Nakon višestrukog pasažiranja, njihov proliferativni kapacitet postepeno opada, i konačno izčezava u procesu koji se naziva ćelijsko starenje ( senecence ). Ograničen životni vek je, izgleda, osobina svih normalnih somatskih ćelija. Jedini očigledan izuzetak su matične ćelije ranog embriona. Ove pluripotentne ćelije mogu da diferentuju u sve somatske i germinalne ćelijske klase. U kulturi, ove ćelije daju postojane ćelijske linije embrionalnih matičnih ćelija (ES) i nikada ne podležu ireverezibilnim genetskim promenama. Izmene obuhvataju nekompletnu replikaciju krajeva hromozoma ili nasumučno, kumulativno, oksidativno oštećenje DNK. Bez obzira koji je mehanizam u pitanju, isključeni (ili nefunkcionalni) mehanizam starenja je opšta karakteistika malignih ćelija. U procesu propagacije ćelija u kulturi mogu se spontano pojaviti potomci koji nemaju ograničen životni vek. Oni predstavljaju imortalizovane ćelijske linije. Ćelije miša skoro
Eksperimentalni onkogeni virusi Onkogeni virusi su imali ključnu ulogu u razvoju molekularne onkologije iz dva razloga. Prvo, ćelije u kulturi se mogu brzo i reproducibilno transformisati infekcijom specifišnim virusima, pa je moguće direktno poređenje transformisanih ćelija i njihovih normalnih prethodnika. Drugo, ovi virusi imaju relativno jednostavne genome pa je olakšana identifikacija i karakterizacija gena odgovornih za kancerizaciju. Sa ove tačke gledišta, dve grupe onkogenih virusa su bile posebno infomativne. Akutno transformišući retrovirusi nose gene odgovorne za malignu transformaciju, koji nisu neophodni za replikaciju virusa. Ovi geni su izvedeni iz ćelijskih homologa – proto-onkogena. Druga grupa, papovavirusi, obuhvata dobro proučene mišje polioma viruse i SV40 majmuna. Ovi virusi nose gene, koji nemaju homologiju sa ćelijskim genima, već nose informaciju neophodnu za replikaciju virusa a mogu da učestvuju i u kancerizaciji specifičnih ćelija. Mnogi onkogeni virusi mogu da indukuju tumore u eksperimentalnim životinjama, a ne mogu da transformišu ćelije u kulturi. Na primer, retrovirusi koji indukuju leukemije ili karcinome nakon dugih latentnih perioda, ne nose posebne gene za transformaciju i ne mogu da transformišu ćelije u kulturi. Papilomavirusi indukuju tumore kod životinja i ljudi, ali ne transformišu ćelije u kulturi ili bar adekvatan in vitro sistem ne postoji. Maligna transformacija i RNK i DNK onkogenim virusima ima jednu zajedničku karakteristiku – transformacija zavisi od integracije virusne DNK u genom ćelije domaćina. Ovo je kritčan korak u ovom tipu kancerizacije, jer obezbeđuje virusnoj DNK replikaciju i propagaciju zajedno sa ćelijskim genomom. Ovakva kontinualna propagacija virusnih gena omogućava i stalnu ekpresiju virusnih proteina neophodnih za održanje malignog fenotipa. Prikaz ćelija u proliferaciji u Petrijevoj posudi. Transformisane ćelije u osustvu kontaktne inhibicije rastu nagomilavajući se u obliku fokusa na monosloju normalnih ćelija Morfologija normalnih i transformisanih fibroblasta (a) Normalni fibroblasti pacova pod fazno-kontrastnim mikroskopom, ćelije su paralelno usmerene i gusto pakovane. (b) Kod embrionalnih fibroblasta pacova koji su transformisani srednjim-T antigenom polioma virusa, ćelije rastu izukršteno i haotično. (c) Kod ovih istih ćelija transformisanih “Abelson-leukemija” virusom miša, ćelije ne pokazuju adheziju i imaju okruglasti oblik. Okrugle ćelije imaju koronu zbog prelamanja svetlosti.
(d) Mikrografija skanirajućeg elektronskog mikroskopa normalnih 3T3 ćelija. Kao i normalni fibroblasti pacova ćelije su usmerene u jednom pravcu i šire se u tanke lamele sa ispupčenjima koje odražavaju položaj jedra. Svaka pljosnata stuktura predstavlja jednu ćeliju. (e) Mikrografija skanirajućeg elektronskog mikroskopa 3T3 ćelija transformisanih Rous sarkoma virusom. Ćelije su postale okruglaste sa procesusima sličnim dlaci i loptastim izraštajima. Ove ćelije rastu jedna preko druge nezavisno od kontakta sa drugim ćelijama. Ćelije embriona pileta, transformisane Rous sarkoma virusom, se nagomilavaju jer rastu bez kontaktne inhibicije. Transformisane ćelije očigledno ne zavise od adhezije za čvrstu podlogu i razvijaju se u kolonije kada se suspenduju u agaroznom ili metilceluloznom polučvrstom medijumu dok se normalne ćelije podele samo jedan ili dva puta nakon suspendovanja u ovim medijumima. Cetiri glavna tipa tumora: klasifikacija je izvrsena na osnovu celijskog tipa koji zahvataju.
- Kanceri - najcesci oblik, transformisane epitelijalne celije koje okruzuju unutrasnje organe i povrsine, pluca, debelo crevo, grudi, prostata...
- Sarkomi - mazenhimska tkiva (fibroblasti i srodne celije), kosti, misici
- Hemopojeticke celije i organi – leukemija, limfomi, mijelom...
- Tumor koji zahvata neuroektodermalne celije – neuroblastomi, glioblastomi, melanomi
- Geneticke mutacije na ovim kriticnim genima vode do ne-regulacije celijskog rasta i deobe
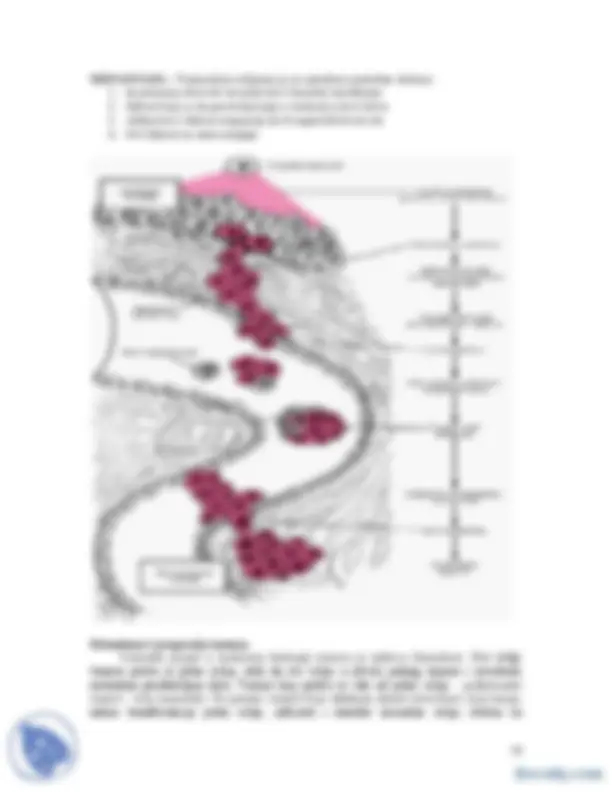
- Kancer se uglavnom desava kao posledica mutacija u somatskim celijama Hromozomske abnormalnosti kod malignih ćelija. Genetska nestabilnost malignih ćelija (a) Filadelfija hromozom c- abl proto-onkogen je translociran sa hromozoma 9 u bcr lokus hromozoma 22. (b) Hromozomske dlecije kod retinoblatoma. Delecije 13q14 se detektuju i kod naslednog i sporadičnog retinoblastoma. (c) Amplifikacija DNK. Zbog reverzibilnosti hromozomskih integracija HSR i DM su izmenljive forme amplifikovane DNK. Posebna specifičnost malignih ćelija je njihova heterogenost. Ćelije u okviru jednog tumora nisu identične iako su potomci jedne ćelije. Ova heterogenost je obično uočljiva ako se analiziraju metafazni hromozomi ili ako se DNK sadržaj pojedinačnih ćelija odredi mikrofluorimetrijom. Metode koje omogućavaju specifično bojenje traka pojedinih hromozoma su omogućile detaljniju analizu ovih promena. Iako je hromozomska heterogenost uobičajena kod malignih ćelija, njeno poreklo je nejasno. Stabilnost normalnog kariotipa zavisi od pouzdanosti enzima koji učestvuju u replikaciji DNK i efikasnosti i preciznosti enzima koji koriguju greške i oštećenja u molekulu DNK. Hromozomska heterogenost, takođe, zavisi od funkcije proteina mitotskog aparata. Ovi proteini obezbeđujuju pravilnu segregaciju hromozoma u ćelije ćerke. Veoma je važan i biohemijski sistem koji reguliše dinamiku ćelijskog ciklusa i koji je pod nadzorom “kontrolnih tačaka” (checkpoint controls) koje, kao složeni enzimski
sistem, proveravaju da li je DNK replikacija kompletirana i sva oštećenja korigovana pre nego što se ostvari ćelijska deoba. Defekti u bilo kom od ovih procesa dovode do hromozomske i genetske nestabilnosti malignih ćelija. Defekti kod enzima koji koriguju oštećenja u DNK i njihova uloga u kancerogenezi su dokumentovani kod nekih naslednih malignih sindroma. Na primer, Xeroderma pigmentosum , je sindrom koji je posledica nefukcionalnosti ekscizionog DNK korekcionog enzimskog sistema, i karakteriše se visokom učestalošću raka kože uzrokovanog UV-komponentom sunčeve svetlosti. Slično, nasledni ne-polipozni karcinom kolona (HNPCC) je posledica defekta kod enzima koji koriguju nekomplementarni bazni par u molekulu DNK. Iako se nedovoljno zna o poreklu hromozomske nestabilnosti ona, nesumnjivo, daje veliki doprinos kancerogenezi. Amplifikacija segmnenata hromozoma povećava broj kopija proto-onkogena u okviru ovih segmenata. Translokacije mogu da povežu prtoto- onkogene sa novim segmentima DNK koji im menjaju funkciju. Delecije mogu da uklone tumor supresor gene koji, normalno, kontrološu i ograničavaju ćelijsku proliferaciju. Hemoterapija citotoksičnim agensima često dovodi do pojave rezistentnih ćelija, kao posledica amplifikacije gena, koji kodiraju enzime koji metabolišu ili transportuju citostatike. Na primer, rezistencija na kolhicin ili deoksirubicin je obično posledica amplifikacije mdr1 gena, koji kodira transportni protein koji eliminiše citotoksični agens iz ćelija. Vaskularizacija je neophodna za konverziju dormantnog karcinoma in situ u invazivni maligni tumor koji može da ubije domaćina. (a) Normalne epitelne ćelije su odvojene od vaskularnog sistema bazalnom membranom. (b) U avaskularnoj fazi, maligne ćelije mogu postojati kao abnormalna izraslina neškodljivog karcinoma in situ mnogo godina. (c) Tumor mora da obezbedi faktor angiogeneze kao što je FGF i VEGF, pre nego što se susedni krvni sud razgrana i pruži kapilare koje prodiru i u masu tumora u ekspanziji. (d-f) Pošto je tumor vaskularizovan, moguć je brzi, karakterističaan rast.
poliklonalni. Postoje i drugi primeri, tumori nastali kao posledica prisustva infektivnog agensa, ili u sličaju naslednih malignih sindroma, kao posledica povećane verovatnoće za inicijacioni događaj. Međutim, ogromna većina spontanih tumora je monoklonalna. Dokaz o monoklonalnosti tumora je dobijen primenom markera koji pokazuju heterogenost normalnih tkiva. Na primer, ženske osobe su heterozigotne u odnosu na markere vezane za pol i pokazuju mozaičnu tkivnu strukturu kao rezultat inaktivacije X hromozoma. Ovaj događaj se dešava rano u procesu razvića i posledica je nasumične inaktivacije jednog od dva X hromozoma u ćeliji. Potomstvo jedne ćelije u kojoj je došlo do ove inaktivacije zadržavaju jedan aktivan i jedan neaktivan X hromozom. Ako je neka žena heterozigotna u odnosu na neki gen na ovom hromozomu, i ako se njen genotip predstavi kao a/A , neke njene ćelije će imati A a druge a fenotip. Tumori koji nastanu kod ovakvih heterozigotnih jedinki će biti ili A ili a fenotip bez karateristične mozaične strukture tkiva. Sličan pristup je iskorišćen za dokaz o monoklonalnosti tumora imunog sistema. Geni koji kodiraju specifične imunoglobuline ili T ćelijske receptore prolaze kroz genske rearanžmane u toku somatskog razvoja, tako da pojedinačne ćelije stvaraju specifične proteine. Tumori limfoidnih ćelija obično nose jedinstven rearanžman u okviru ovih gena. Ako tumorske ćelije vrše sekreciju nekog imunoglobulina, kao kod multipnog mijeloma, sekvenca ovog proteina je specifična samo za taj tumor i svi molekuli imaju istu sekvencu, što dokazuje monoklonalno poreklo tumora. Otkriće da je geneza tumora ekspanzija klona nosi važne teorijske implikacije. Na prvom mestu, jasno je da je maligno stanje suštinska i nasledna osobina somatskih ćelija i da je iniciacioni događaj retka pojava. Češći iniciacioni događaj bi nužno rezultirao u poliklonalnim tumorima. Stabilne promene u ekspresiji gena (epigenetske promene) mogu, takođe, da objasne neke osobine malignih ćelija. Međutim, genetske promene u proto-onkogenima i tumor supresor genima su primarni uzrok inicijacije i progresije tumora. Heterogenost tumora Eksperiment pokazuje klonalno nasledne razlike između ćelija tumora u odnosu na njihov metastatski potencijal. Ćelije izvedene iz jedne maligne ćelije se subkloniraju. Standardni alikvoti se i.v. injeciraju ekperimentalnoj životinji. Subklonovi imaju veoma razlčit metastatski potencijal, meren brojem pigmentisanih metastaza u plućima životinje.
EPIDEMIOLOGIJA KANCERA
Cancer epidemiology is the study of the pattern of cancer appearance in populations and is predominantly statistical, its methods and conclusions being expressed in terms of probabilities; for example, 'Japanese women have less than a quarter the breast cancer risk of US women' or 'women who have a baby before age 20 have only half the chance of getting breast cancer of women without children'. Its primary objective, in the area of public health, is to identify preventable (avoidable) causes of cancer, but it also has an important role in other areas of cancer research; for example, in the evaluation of screening tests to detect cancer at an early stage and in aiding the understanding of the mechanistic basis of cancer. The most basic task of cancer epidemiology is simply to describe the occurrence of human cancer, noting differences between, for example, males and females, people of different ages, different socioeconomic classes, people in different occupations, different time periods, different areas of a country, and different countries. This descriptive epidemiology has been a most fruitful source of ideas as to the possible causes of various cancers. For example, the enormous rise of lung cancer in men, but not in women, between 1920 and 1945 suggested that some recently introduced habit of men, but not of women, must be responsible, and cigarette smoking became the prime candidate. More recently the finding of large differences in the occurrence of colorectal cancer between different countries has led to intense investigation of the possible roles of various aspects of the normal diet (such as fat or fiber content) as factors in the etiology of this cancer. Descriptive epidemiology Incidence rates To describe the differences in occurrence of a particular cancer between different groups the most useful concept is that of an incidence rate—the probability of an individual in a particular group being newly diagnosed as having the particular cancer within a year. In epidemiological studies these probabilities are usually expressed as a rate usually 'per 100000' or 'per million'. For example, the incidence rate of breast cancer in females in England and Wales in 1989 was 107,2 per 100000; this was estimated by noting that there were 25 893 400 females in England and Wales on 30 June 1989 and that 27 768 new breast cancer cases were diagnosed in females in that year: 27 768/ 25 893 4000 = 0.001072 or 107.2 per 100 000. Age-specific incidence rates For most forms of cancer the incidence rate increases rapidly with increasing age. For etiologically meaningful comparisons, incidence rates must be worked out separately for groups of people of a similar age. The collection of such incidence rates over a span of age groups is referred to as an age-specific incidence curve. Table 3.1 and Fig. 3.1 show the age-specific incidence rates for cancer of the colon in females in England and Wales in the period 1983-7. These particular age- specific incidence rates display a more than 100-fold increase between age 25 and age 70. It is most useful to plot both incidence rate and age on logarithmic scales, because the 'log-log' plot thus obtained is, for cancers at many different sites, very close to a straight