Preuzmite MALIGNA CELIJA-skripta-predavanje Part4 i više Rezime u PDF od Biologija ćelije samo na Docsity!
Oncogenes and Tumor Suppressor Genes in Tumors Induced by Chemical Carcinogens In the transgenic mouse models discussed in the preceding section, the inherited oncogenes apparently initiate the process of tumorigenesis by driving the proliferation of a preneoplastic cell population from which neoplastic cells eventually arise. In these cases, the role of the oncogene in initiation of tumor development is clearly defined, because germ line transmission of an activated oncogene has been experimentally used to induce neoplasia. The roles of tumor suppressor genes in inherited cancers resulting from germ line mutations of these genes is similarly evident, as is the role of germ line mutations that activate the ret oncogene in cases of inherited multiple endocrine neoplasia type 2. An independent question concerns the roles of oncogenes and tumor suppressor genes in non- inherited neoplasms. Animal models have again provided important information because tumors can be induced reproducibly by known carcinogenic agents, allowing the analysis of oncogene activation at discrete stages of the neoplastic process. Chemical carcinogenesis in particular has provided an experimental system in which activation of oncogenes, particularly members of the ras gene family, can be directly related to the initiation of neoplastic disease. One of the classic models for multistep carcinogenesis is the induction of tumors in mouse skin by the topical application of chemical carcinogens. Tumorigenesis in this system can be divided experimentally into distinct stages of initiation and promotion. Initiation is an irreversible step resulting from a single carcinogen treatment. Tumors do not develop, however, unless the mice are subsequently treated with a tumor promoter, usually a phorbol ester. Treatment with tumor promoters can take place up to a year after carcinogen treatment, indicating that cells with the initiated phenotype persist in the skin and are irreversibly committed to eventual neoplastic progression. Because the carcinogens active in initiation are mutagens, it is likely that initiation results from direct damage to DNA. In contrast, the tumor promoter phorbol esters are not mutagens but act to stimulate cell proliferation. They are therefore thought to lead to neoplasm development by stimulating the growth of initiated cells. The tumors that develop following initiation and promotion are primarily benign papillomas. Some of these evolve further to malignant carcinomas, illustrating yet another stage in tumor progression.
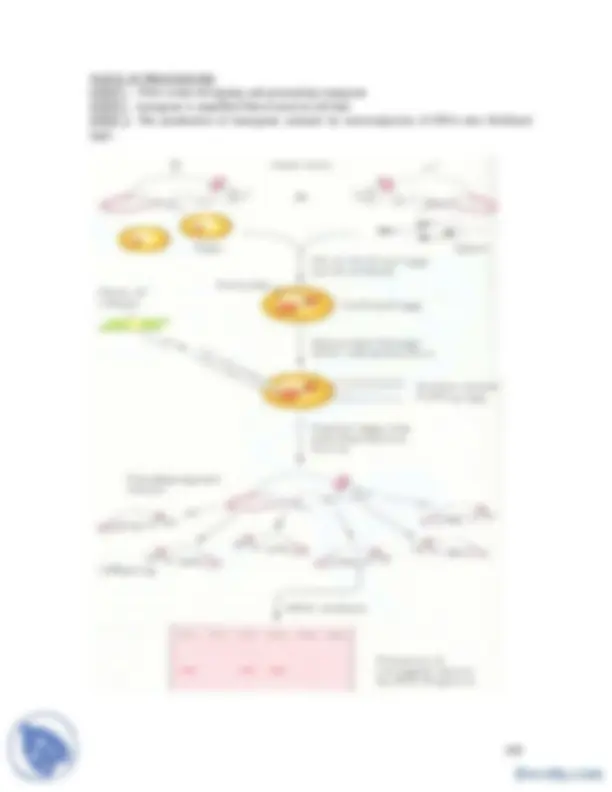
Accumulation of mutations during tumor progression
Genetic pathways to colon cancer - “Multi-stage carcinogenesis”
EPIGENETIC TUMOR SUPPRESSOR GENE SILENCING ------------
REAL-TIME PCR
But first, some basic things:
Slika 1. Energetski nivoi molekula So-osnovni elektronski energetski nivo S1-ekscitirano stanje U okviru oba ova stanja postoji veliki broj definisanih vibracionih energetskih nivoa, koji opisuju opseg dozvoljenih medjunuklearnih rastojanja. Krive imaju dati oblik jer na malim rastojanjima izmedju vibrirajucih nukleusa dolazi do odbijanja, i otud nagli rast energije, dok sa “preteranim” povecanjem rastojanja dolazi do kidanja veze. Osnovni energetski nivoi imaju najmanju energiju.
Slika 2. Apsorpcioni I emisioni spektri fluorofore. Fluorofora je molekul, anajcesce deo molekula koji moze da fluorescira. Dati spektri su simetricni zbog slicnosti vibracionih nivoa u okviru dva elektronska energetska nivoa (So i S1). Preklapaju se za prelaz iz najnizeg u najnizi vibracioni nivo So i S1. Takodje je ilustrovana cinjenica da su emisioni spektri pomerani ka vecim talasnim duzinama (crveni sift) u odnosu na apsorpcine zbog brze vibracione relaksacije (VR) na osnovni vibracioni nivo u okviru S1.
exponential amplification is not ideally suited to quantification. Small differences in amplification efficiency between samples can become huge differences in results when they are amplified through forty doublings. Anyone working with quantitative PCR who forgets this fact is in danger of making mistakes that are measured in orders of magnitude. Why then the continuing increase in the use of quantitative PCR? It has a sensitivity five orders of magnitude better than the best blotting procedures and a dynamic range of 10 orders of magnitude. This unsurpassable sensitivity and range has made the work of turning PCR into a quantitative tool worthwhile. Real time PCR: Applications in cancer research Detection of minimal residual disease Tumor immunology Gene copy number (oncogene amplification) Genomic mutation and polymorphism Tumor specific alterations in transcription levels Tumor specific alterations in CpG island methylation levels
Real-time PR is the very best way to do quantitative PCR
A reaction profile can be thought of has having three segments: an early background phase, an exponential growth phase (or log phase) and a plateau. The background phase lasts until the fluorescence signal from the PCR product is greater than the background fluorescence of the probe system. The exponential growth phase begins when sufficient product has accumulated to be detected above background, and ends when the reaction efficiency falls as the reaction enters the plateau.
In real-time PCR fluorescence values are recorded during every cycle and represent the amount of product amplified to that point in the amplification reaction. Moreover, these values are recorded separately for each of the reaction steps: denaturation, annealing and extension. The more templates present at the beginning of the reaction, the fewer number of cycles it takes to reach a point in which the fluorescent signal is first recorded as statistically significant above background, which is the
definition of the (Ct) values. This will increase the throughput, because it is no longer necessary to analyze dilutions of each sample in order to obtain accurate results as it is the case with competitive PCR. A number of options are available for implementing real- time PCR in quantitative analysis. Homogeneous detection of PCR products can be done using double- stranded DNA binding dyes; fluorogenic probes, direct labeled primers and a primer-probe combination termed scorpion.
Advantages – collects data in the exponential growth phase
- rael time: permanent record of amplification
- increased dynamic range of detection
- less RNA needed, Requirement of 1000-fold less RNA than conventional assays
- fast; No-post PCR processing
- Sensible: Detection is capable down to a 2-fold change
Real time PCR offers numerous advantages over previous attempts at quantitating PCR. Other methods typically rely on end-point measurements, when often the reaction has gone beyond the exponential phase because of limiting reagents. To compensate for such problems, competitive PCR was devised, which allows for normalization of the end product based on the ratio between target and competitor. Because this method is cumbersome, requiring a carefully constructed competitor target for each PCR reaction and a series of dilutions to ensure that there is a suitable ratio of target to competitor, it is seldom used successfully. In contrast, with real time PCR, the dynamic range is much greater than that of competitive PCR (over six orders of magnitude as compared to one with competitive PCR), post-reaction processing is eliminated, and the measurements are taken from the exponential range of the reaction, where component concentrations are not limiting. And best of all, the entire process is automated. The theory is straightforward, but a number of technical caveats are associated with the use of conventional end-point methodologies for quantitative PCR. In these techniques, PCR results are monitored after a given number of cycles, by which point factors such as limiting reagent concentrations and side reactions may have played a significant role in affecting final product concentration. Quantitative competitive PCR was developed in response to some of these difficulties. In this approach, the starting amount of target is calculated based on the ratio of target to competitor after amplification. However, quantitative competitive PCR is cumbersome, and it can be associated with a number of drawbacks including a limited dynamic range and the need to screen multiple dilutions. The first on the market, the ABI 7700 Sequence Detection System includes a built- in thermal cycler, a fluorogenic 5' nuclease assay, a laser for inducing fluorescence, charge-coupled device (CCD) detection, and PCR application software. The specially
Applied Biosystems has commercialized the requisite reagents in its well-known TaqMan® product line still the most widely used real-time reagent. The fluorogenic probe is complementary to the target sequence, and initially contains both reporter and quencher moieties. When the probe is not bound to template DNA, its reporter and quencher dyes are in close proximity, and the reporter's fluorescent emission is quenched.
Diagram of quantitative RT-PCR (TaqMan) technology
A , in the PCR reaction, an oligonucleotide probe tagged with a 5’ fluorescent reporter and a 3’ quencher is added in addition to the standard PCR components. The probe is complementary to the target sequence of interest and anneals during extension. The close proximity of the quencher to the fluorescent reporter represses fluorescence in the intact probe. As the Taq polymerase synthesizes the new strand, its 5’ to 3’ nuclease activity cleaves the probe, separating the quencher and fluorescent reporter. The fluorescence emitted is proportional to the amount of product accumulated with each cycle. B , the plot represents a sample analysis from the experiments shown in the Figure. The plot shows the expression values obtained for three genes (as indicated) in a matched tumor and normal tissue sample. The horizontal bold line indicates the fluorescence level
used for the threshold cycle (C t ) determination in this particular example. D Rn is defined as the cycle-to-cycle change in the reporter fluorescence signal normalized to a passive reference fluorescence signal (background).
DNA Quantification
Using either the DNA binding dye SYBR Green I or Hybridization Probes, the progress of amplification is monitored at each cycle. In many cases, it only takes a few minutes to determine whether or not a sample contains a certain template molecule. In the following, the principles of PCR data quantification are explained using schematic drawings.
In this model experiment, 10^6 , 10^5 and 10^4 template molecules were amplified. The Y-axis shows the fluorescence whereas the X-axis shows the number of cycles. As expected, the less template molecules are contained in a sample the more PCR cycles are required before the amplification signal can be detected. This interdependency between the number of PCR cycles that are required to detect a template (that is fluorescence raises above the background – threshold value - Ct) and the number of template molecules initially present in a sample can be used for the quantification of unknown samples.
To this end, first a calibration graph is calculated. For each sample with known template concentration it is determined at which PCR cycle the amplification signal enters the log linear region (black dots in upper chart). These points are located on a line
which is referred to as the crossing line (Ct value). Using these intersection points a calibration graph can be calculated which is shown in the lower part of the figure. This calibration graph defines a relationship between the cycle number at which the amplification signal intersects the crossing line and the template concentration initially present in the sample. Thus, if the intersection point of an amplification signal (expressed as the cycle number) is known, the initial template concentration can be directly derived from the calibration graph. This is how the template concentration of samples is determined after the calibration graph has been calculated. SYBR Green I exhibits very little fluorescence when free in solution; emission is greatly enhanced when it binds to the minor groove of the DNA double helix. Prior to amplification, the reaction mixture contains the denatured DNA, the primers, and the dye. The low-level background fluorescence signal generated by the unbound dye molecules is subtracted during computer analysis. After annealing of the primers, a few dye molecules can bind to the double strand. During elongation, more and more dye molecules bind to the newly synthesized DNA, resulting in dramatically increased light emission. If the reaction is monitored continuously, this increase in fluorescence can be viewed in real time. After denaturation of the DNA during the next heating cycle, the dye molecules are released and the fluorescence signal falls. A fluorescence measurement is performed at the end of the elongation step of every PCR cycle to monitor the increasing amount of amplified DNA. The major disadvantage of the method is that the SYBR Green I binds both specific and non-specific DNA amplicons.
Molecular Scorpions Scorpions are bi-functional molecules containing a PCR primer element covalently linked to a probe element. The molecules also contain a fluorophore that can interact with a quencher to reduce fluorescence. When the molecules are used in a PCR reaction the fluorophore and the quencher are separated which leads to an increase in light output from the reaction tube. The benefits of Scorpions derive from the fact that the probe element is physically coupled to the primer element - this means that the reaction leading to signal generation is a unimolecular rearrangement. This contrasts to the bi-molecular collisions required by other technologies such as TaqMan or Molecular Beacons. The benefits of a unimolecular rearrangement are significant - as the reaction is effectively instantaneous it occurs prior to any competing or side reactions such as target amplicon reannealing or inappropriate target folding. This leads to stronger signals, more reliable probe design, shorter reaction times and better discrimination. The presence of the blocker group is an essential element of the Scorpions invention. Without such a blocker the Taq DNA polymerase would be able to read through the Scorpions primer and copy the probe region. This would generate signal but not in a target specific fashion. Copying the tail in this way would completely negate the benefits of the Scorpions reaction as any inappropriate side-reactions, including the formation of primer dimers, would also generate a signal.
Figure: Detection of PCR products with self-probing amplicons. (A) A Scorpions primer carries a 5’ extension comprising a probe element, a pair of self-complementary stem sequences, and a fluorophore/quencher pair. The extension is “protected” from copying by the inclusion of a blocking HEG monomer. (B) After a round of PCR extension from the primer, a newly synthesized target region is now attached to the same strand as the probe. (C) Following a second round of denaturation and annealing, the probe and target hybridize.
ALLEL SPECIFIC OLIGONUCLEOTIDE (ASO)- Allelic Discrimination Using the 5´-Nuclease Assay (Mutation detection). In allelic discrimination assays, the PCR assay includes a specific, fluorescent, dye- labeled probe for each allele. The probes contain different fluorescent reporter dyes (FAM and VIC™) to differentiate the amplification of each allele. Mechanism: relies on competition between the two probes. Tm of the mismatched probe < Tm of perfectly matched probe
EPIGENETIKA KANCERA
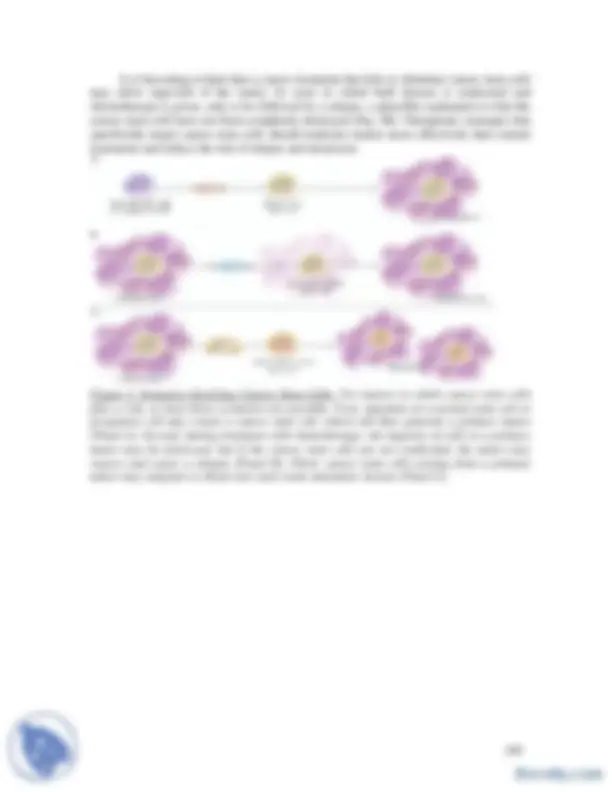
The term “epigenetic” refers to a heritable change in the pattern of gene expression that is mediated by mechanisms other than alterations in the primary nucleotide sequence of a gene. Methylome analysis increased dramatically in more recent years, however, because of convincing evidence of abnormal silencing of genes in cancer cells. The change in gene expression in cancer cells may involve the methylation of DNA in promoter regions of these genes, the sites where transcription of DNA into RNA begins. An important aspect of the methylation mechanism is that it inactivates tumor-suppressor genes. Current efforts to understand the mechanisms of gene silencing, and to find ways of exploiting the diagnostic and therapeutic implications of the abnormality, fit together, benefit from, and contribute to the explosion of investigations into the control of gene expression, one of the most vibrant areas of cancer research. Cytosine methylation is a modification of the genome, as opposed to being part of the genome, so is known as "epigenetics" (Greek for "upon" genetics). Dense methylation of promoter regions is associated with a compacted chromatin structure, and accompanying transcriptional silencing of the affiliated gene. This form of information storage is flexible enough to be adapted for different somatic cell types, yet is stable enough to be retained during mitosis and / or meiosis and is referred to as EPIGENOME In recent years, it has become apparent that the transcriptional silencing that is associated with 5-methylcytosine is important in mammalian development, protection against intragenomic parasites, genomic imprinting, x-inactivation, mental health, aging and cancer CpG methylation is the most common epigenetic modification of the genome. The haploid human genome contains approximately 50 million CpG dinucleotides, which are capable of encoding 250,000,000 different permutations per haploid genome. The number of methylation changes in cancer cells seems to outnumber genetic alterations. The problem is not the lack of information in the epigenome, but the difficulty in accessing that information. Epigenetics creates transcriptional chaos in cancer and provides molecular basis for an ever increasing malignant potential Oncogenic DNA methylation alterations occur in two directions:
- hypermethylation of CpG islands that leads to the silencing of tumour suppressor genes
- hypomethylation of dispersed CpGs increases transposition of movable genetic elements and transcription of normally dormant proto-oncogenes It is important to define whether aberrant profiles of CpG island hypermethylation cancer are a consequence of a targeted process or a random process followed by clonal selection.
A study undertaken to identify all differentially methylated CpG islands in cancer estimated that of the 45 000 CpG islands in the human genome, 600 exhibit methylation patterns in tumors different from those in normal tissues. CpG is the only dinucleotide to be severely under-represented in the human genome (5 to 10-fold) due to methylcytosine- to-thymine transition mutations
The normal silencing of the imprinted allele is equivalent to a first ‘hit’ in the Knudson’s "two-hit hypothesis" for carcinogenesis. Since most imprinted genes are involved in cell growth and all are functionally haploid, they represent unique susceptibility loci for cancer development. There is now compelling evidence of a mechanistic link between the ageing process and tumorigenesis in that age-related promoter hypermethylation frequently occurs in genes known to be involved in cancer formation.
A TUMOR-SUPPRESSOR GENE INACTIVATED BY CpG ISLAND HYPERMETHYLATION WOULD HAVE A NUMBER OF IMPORTANT CHARACTERISTICS. THESE INCLUDE:
- Dense methylation of the normally unmethylated CpG island present in the promoter region
- An absence of coding region mutations in the tumor
- A deficiency of gene-specific transcripts in the tumor
- Gene reactivation in the tumor with DNMT inhibitors (e.g. 5-azaC)
- Loss of gene function from hypermethylation comparable to that seen for inactivating mutations
promoter region within the barriers. The lower panel depicts the breakdown of the barriers in a cancer cell, in which the transcriptionally repressive chromatin and DNA methylation have spread into the CpG island in the promoter region and correlate with transcriptional repression (red arrow with X) of the gene. The DNA-methylating complex now has access to the region, and the transcriptional machinery (transcriptional co activators, histone acetyltransferase, and transcription factors) is excluded
DNA methylation profiles offer several advantages over gene-expression microarrays and proteomic approaches. First, the DNA molecule is very stable, surviving routine processing for histopathology. Second, measurements of DNA methylation can be compared with absolute reference points (completely methylated or completely unmethylated DNA). This greatly simplifies the design of internal references for methylation assays. Third, abnormal methylation patterns in cancer cells differ qualitatively from normal cells, not just quantitatively. This allows for the development of assays with high specificity and sensitivity. This can be an advantage in analyses of samples that contain substantial amounts of stromal tissue or non-malignant epithelium. A fourth advantage is that methylation assays for individual markers tend to be universal, just like gene-expression markers. Genetic mutation assays usually have to be tailored to the individual tumor. Finally, a fifth advantage is that DNA methylation patterns are fairly stable over time. They do not fluctuate in response to short-term stimuli, as gene-expression profiles do.
The methylome analysis aims to the following endpoints: ► The total genomic methylation content represents the total amount of methylated cytosine in the genome, expressed either as a fraction of cytosines, or as a fraction of CpG dinucleotides. ► Methylation level refers to the average methylation occupancy at a single CpG dinucleotide.. ► Methylation pattern refers to a series of CpG dinucleotides located in cis on a single DNA strand. DNA methylation patterns can be determined by bisulphite genomic sequencing of subclones. Methylation-specific polymerase chain reaction measures the presence of one particular methylation pattern in a pool of DNA. ► A methylation profile refers to the measurement of DNA methylation at multiple sites throughout the genome. This can either be at individual CpG dinucleotides as a level profile or by the measurement of multiple methylation patterns.