Download Comparison of Digestive Function Among Rabbits, Guinea ... and more Schemes and Mind Maps Physiology in PDF only on Docsity!
Comparison of Digestive Function Among Rabbits, Guinea-Pigs, Rats and
Hamsters. I. Performance, Digestibility and Rate of Digesta Passage
Peter Wen-Shyg Chiou, Bi Yu and Chung-Yi Kuo*
- Address reprint request to Peter Wen-Shyg Chiou. Tel: +886-4-287-0613, Fax: +886-4-286-0265, E-mail: wschiou@ dragon.nchu.edu.tw. Received Januaiy 26, 2000; Accepted June 27, 2000
Department of Animal Science, National Chung-Hsing University, 250 Kuo-Kuang Road, Taichung, Taiwan, ROC
ABSTRACT ' This trial was to compare the digestive function of laboratory animals, which included omnivores (rats) and herbivores (rabbits, guinea pigs and hamsters). Ten weanling rats, rabbits, guinea pigs and hamster were randomly allocated in individual metabolic cages for a performance and a digestibility trial. Results showed that guinea pigs were significantly best in digestion of dietary crude protein and fiber. Rabbits, however, had the poorest nutrient digestion among the herbivores. Crude fiber digested significantly less by rats than by the herbivores (p<0.05). The digesta retention in the GI ■ tract was longest in rabbits followed by guinea pigs, rats and hamsters. The relative length of the large intestine was significantly shorter in rats as compared to guinea pigs or hamsters. Both the weight and length ratio of the caecum or colon-rectum to the large intestine reflected the major site for fermentation that was the caecum in rabbits and the colon-rectum in guinea pigs. 10% of crude fiber diet did not result in damaged mucosa in any of the experimental animals in this trial. (Asian-Aus. J. Anim, Sci. 2000. Vol. 13, No. 11 : 1499-1507)
Key Words : Herbivores, Laboratory Animals, Performance, Digestibility, Rate of Digesta Passage, Crude Fiber
INTRODUCTION
The animals that are most extensively use in the laboratory are rabbits, guinea pigs, rats and hamsters. These animals are all hindgut fermenters and herbivores, except rats which are omnivores. Growing rabbits (NRC, 1977) and guinea pigs (NRC, 1995) require 10 — 12% and 15% crude fiber, respectively, in their diet. Although the specific amount of crude fiber required by rats and hamsters has not yet been determined, the crude fiber in their diets has generally been emphasized due to its significance in digestive function. Research data have suggested that a 10% dietary fiber or cell wall is required in the diet for rats (Nyman and Asp, 1985; Pond et al., 1989). The hindgut which includes the caecum, colon and rectum is the major site of fermentation in these herbivore non-ruminants, where volatile fatty acids (VFA), end products of fermentation, are absorbed by the host animal as an. energy source (Parker and McMillan, 1976; Zhao et al., 1995). Cecotomy of these animals significantly depressed rat and hamster ability to digest dietary dry matter and crude fiber (Sakaguchi et al., 1981; Williams and Senior, 1982), but showed no significant impact in rabbits (Gioffre et al., 1980). Rabbits are not good digesters of fiber. They digest crude fiber less well as compared to guinea pigs, but can digest cellulose and hemicellulose as well as rats (Fonnesbeck et al., 1974). Rabbits, however, cannot survive on a low fiber diet (6% of crude fiber). This is also true for rats and hamsters on a crude fiber free diet. Feeding rats and hamsters on
this kind of diet causes high mortality or depressed... growth (Cheeke and Patton, 1980). On the other hand, dietary crude fiber is beneficial to guinea pigs in depressing the growth of pathogenic bacteria,;; Bacteroides ovatus, in the intestine, hence improving the growth of guinea pigs (Johanning et al., 1984). Increased levels of crude fiber in the diet generally decrease feed conversion efficiency in rabbits and rats. This may be attributed to the changes in the GI tract that enhance the digestive function in these animals. In the rats an increase in the level of dietary fiber , significantly increases the intestinal weight and length : of various sections of the GI, but decreases nitrogen retention which results in decreased growth (Zhao et aL, 1995). For non-ruminant herbivores, dietary fiber is therefore, significant as a source of bulk in the digestive physiology, not as a source of nutrients. The biological classifications of these laboratory , animals are closely related. Rabbits belong to the: order of Lagomorpha, Rats and hamsters belong to order of Rodentia- and guinea pigs are hystricomorph, r: Some specific digestive functions in the various ; sections of the GI tract should be similar among these ; animals. Research information is limited on the comparative effects of crude fiber on their digestion. This trial is therefore aimed at study of the differences in digestive function in these laboratory animals and their utilization of crude fiber.
MATERIALS AND METHODS
Animal and diets Ten four-week old weanlings of each animal species with equal numbers of each sex and a mean live-weight of 1,021 g for the California rabbit, 175 g for the Hartley guinea pig, 165 g for the' Wistar rat
1500 CHIOU^ ET^ AL.
and 71 g for the Syrian hamster, were selected for this feeding trial. These animals were from the National Laboratory Animal Breeding and Research Center at Taiwan. The experimental diet is presented in table 1. This diet was formulated to contain 18% crude protein and 10% crude fiber according the nutrients requirement for rabbits (NRC, 1977), and laboratory animals (Sakaguchi et al., 1987; NRC, 1995). The diet was presented in pellet form with size of 0.6 by 1.5 cm.
Feeding tri 이 and digestibility tri 이 All experimental animals were randomly allocated in individual metabolic cages. After one week of adaptation, the animals began the four-week feeding trial in growing period. These animals were individually fed ad libitum once daily at 7:00 AM. Water was provided with free access. Additional ascorbic acid was provided through drinking water (200 mg/L) for guinea-pigs. Light period was 12L:12D and ambient temperature set at 23+1 °C with 40-70% humidity. During the feeding period, feed intake and live-weight were recorded weekly. A digestion trial was conducted at the end of the feeding tri이 that included three days of adaptation followed by five days of total fecal collection.
Determination of digesta retention time Four animals with equal numbers of each sex from each species of animal were selected for the digesta retention study. The animals were fasted for 8 hours before being fed for three hours on a marker diet (1% Cr 2 O 3 ). After 3 hours of feeding, animals were returned to feeding on the basal diet. Feces were collected every single hour within the first 48 hours after animals were fed the marker diet. From the 48th to the 72nd hour after marker diet feeding, feces were collected every four hours for analysis of CdOe The turnover time of each marker was estimated from the decline in the fecal marker concentration according to the following equation (Brandt and Thacker, 1958)
Y = Yo X e-k,
Where Y is the concentration of C12O3 in feces at time t; Yo is the constant depending on the level of CDO3 fed; k is the rate constant; t is the time interval after feeding of the marker (h). Turnover time was calculated as the reciprocal of k. Total mean retention time (MRT) in the gastro-intestinal tract was calculated as the sum of k- and the transit time (TT). Transit time equaled the first appearance of the marker after a dose.
The GI tract histology and morphology by SEM At the end of the fecal collection, all animals were sacrificed after 12 hours fasting. The intestinal tracts were then removed to measure the weight and length of each intestine segment. These gut segments included stomach, duodenum (from pylorus to end of duodenal loop), jejunum, ileum, cecum, colon and rectum. A two-centimeter section of the caecal and the colonic-rectal samples were taken and rinsed with 0. M KC1. A 0.5-cm section of sample was taken with a sharp knife for electron microscope (SEM) scanning according to Paulini et al. (1987) and Moore et al. (1988). All samples were fixed initially in 10% buffered formalin pH 7.0. The specimens were washed (0.1 M phosphate buffer pH 7.3, 3 times for 10 min), and placed in 1% osmium tetroxide for 1 hour. These samples were washed again (phosphate buffer 4 times for 15 min) and then gradually dehydrated by increasing the alcohol concentrations: 50, 70, 80, 90 and 95% for 10 to 15 min each time until finally the concentration was increased to 100% - 3 times. Specimens were then subjected to critical point drying and mounting on aluminium stubs, coated with gold for half an hour and placed in the SEM (Bausch & Lomb Ltd., Nonolab 2100) for scanning.
Analytical methods The fecal samples from the digestion trial and the passage rate trial were oven dried at 60 °C for three days, followed by two days at room temperature, and
Table 1. Composition of experiment diet (%) Ingredients Ground com 38. Wheat bran 6. Soybean meal 21. Alfalfa meal 29. Soybean oil 2. Dicalcium phosphate 2. Salt 0. Premix* 0. Total 100. Analyzed value Dry matter 90. Crude protein 17. Crude fiber 9. Neutral detergent fiber 20. Acid detergent fiber 11. Ash 4.
- Premix (Per kg of diet): Cu(CuSO 4 • 5H?O), 8 mg: Fe(FeSO 4 • 7HQ), 80 mg; Zn(ZnSO 4 ), 120 mg; Mn(MnSO 4 - 5H2O), 140 mg; Choline (chloride), 1 g; Vitamin B 6 , 40 mg; Vitamin B 2 , 80 mg; Niacin, 200 mg; Vitamin E, 50 mg; Ascorbic acid, 200 mg.
1502 CHIOU ET AL.
Table 4. Turnover time (1/k, h), transit time (TT, h) and mean retention time (MRT, h) of digesta in rabbits, guinea pigs, rats and hamsters Rabbits (n=4) Guinea pigs (n=4) Rats (n=4) Hamsters (n=3) l/k 2 36.6±4.6la 28.5 ±4.6邳 18.2±4.6b 19.8±5.3b TT 3 2.5±0.7 3.0±0.7 2.5±0.7 4.7±0. MRT 4 39.1±4.5a 31.5±4.5ab 20.7±4.5b^ 23.8±5.1b Mean±SD. 2 Rate-constant which is considered as a dilution rate (/h) of the marker in the pool of the digestive tract. 3 Time-interval between feeding and first appearance of the marker in the faces. 4 Sum of 1/k and TT. The regression line of the time-course reductions in Cr are expressed as Y = Yoxe'kI; where Y is the concentration of Cr at time t, Yo is the constant depending on the level of Cr feed, k is the rate-constant and t is the time interval after feeding of the marker (h). a,b,c Means within the same row without the same superscripts are significantly different (p<0.05).
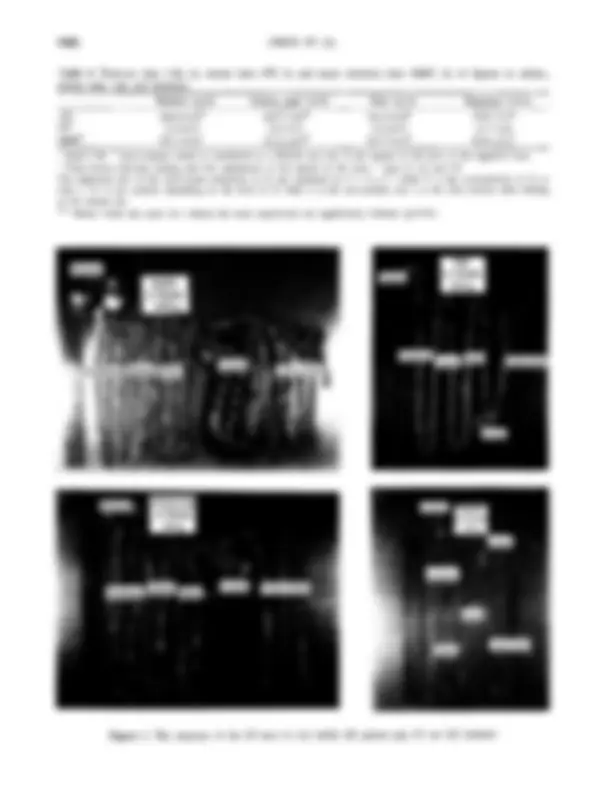
Figure 1. The anatomy of the GI tract in (A) rabbit (B) guinea pig (C) rat (D) hamster
DIGESTIVE FUNCTION ON LABORATORY ANIMALS 1503
parts of the GI tract were also noted. The relative weight and length of various segments of the GI tract differed significantly among the d迁f&ent animals (table 5). The relative gastric weight was the heaviest in rabbits among the four species of animals. In regard to the length of the various segments of the GI tract, the ratio of the jejunum to the entire length of the small intestine was significantly larger in rats than in the other animals. The length of the large intestine in rats was significantly shorter as compared to that in guinea pigs of the same size or hamsters of a smaller size (p<0.05). From the SEM microphotographs of cecal and colonic mucosa showed in figures 2 and 3, the short dense and folded appear in the hindgut mucosa is different from the tongue-like or rod sharp villi in the mucosa of the small intestine. The folded mucosa appears primarily in the larger body sized rabbits,
followed by guinea pigs. This may be related to the volume of GI tract for fermentation.
DISCUSSION
Hamsters made the lowest daily liveweight gain and ate more feed per unit of liveweight than the other animals. Rabbits made the highest daily gain and ate significantly less feed per kg of liveweight than hamsters (p<0.05). When intakes per unit of metabolic bodyweight (W 075 ) are compared, hamsters ate significantly less than the other animals (p<0.05), intakes increasing in the order guinea pigs, rabbits, and rats. The lower feed intake of hamsters per unit metabolic body mass may be attributed to its hibernation characteristic. Hamsters require less energy for maintenance, hence less food, because they hibernate in a comfortable and dim-light environment
Figure 2, SEM microphotographs of caecal mucosa in (A) rabbit (B) guinea pig (C) rat (D) hamster
DIGESTIVE FUNCTION ON LABORATORY ANIMALS 1505
The retention time in the digestive tract and the level of dietary crude fiber is negatively correlated. A level of 10% dietary crude fiber is high for the omnivorous rats. The retention time or turnover time in the GI tract was significantly shorter in rats than in the three herbivores, hence a significantly lower amount of nutrients was digested, crude fiber in particular (p<0.05). Zhao et al. (1995) reported that rats given a high fiber diet demonstrated an increased length in the intestinal tract to compensate for the rate increase in digestive tract passage. Our data, however, did not agree with the results of Zhao et al. (1995). We found that the relative tot이 weight of the small intestine (g/100 g live-weight) in rats did not significantly differ from that in guinea pigs and rabbits (p그0.05) (table 5), but was significantly heavier than that of hamsters (p<0.05). Conversely, the relative weight of the hindgut, which includes the caecum and colon-rectum, was significantly lighter in omnivorous rats than in the herbivores (p<0.05) (table 5). Rats with a shorter and lighter hindgut, exhibited a shorter mean retention time in the dige아ive tract. Both the weight and length ratio of the caecum to the large intestine, 78.6% and 39.4% respectively, were larger in
rabbits than in the other species. Guinea pigs, on the other hand, possess the largest colon-rectum to large intestinal length ratio. This implied that the major site for hindgut fermentation was the caecum in rabbits and colon-rectum in guinea pigs. It appears that the four laboratory animal species fed the diet with same crude fiber (10%) content, not only influenced the volume and appearance, but also the morphology of the inte아in이 mucosa were different among four species. Rabbits or rats produce an under-developed mucosa in the caecum due to a decreased proliferation of epithelial cells when a low fiber diet is fed to rabbits (4.5% crude fiber) (Yu and Chiou, 1996) or a crude fiber free diet is given to rats (Storme and Williams, 1981). Feeding a high fiber diet to these anim이s increases production of VFA from hindgut fermentation, which stimulates epithelial cell proliferation. However, animals will adapt to the prolonged ingestion of a high fiber diet by both morphological and physiological adaptation in the GI tract. Different dietary fiber components act differently upon the intestinal morphology. Supplementation of bulky insoluble fiber in the diet neither stimulated proliferation nor damaged the epithelial cells, but
Table 5. The weight and length of the gastrointestinal tract in rabbits, guinea pigs, rats and hamsters Items Rabbits Guinea pigs Rats Hamsters Weight, g Stomach 79.9 ± 18.77Ia 7.2 ± 2.46° 6.6 ± 1.79° 2.3 ± 0.62° Duodenum 10.1 ± 1.17a 2.8^ ±^ 0.77b^ 1.1 ± 0.1 9 C 0.7^ ±^ 0.32c Jejunum 64.7 ± 14.40a 13.1^ ±^ 3.77b^ 9.5^ ±^ 1.83b^ 1.5^ ±^ 0.26c Ileum 18.8 ± 4.09a 2.7^ ±^ 0.88b^ 3.2^ ±^ 0.35b^ 0.5^ ±^ 0.1^2 C Caecum 148.7 ± 23.65a 34.5^ ±^ 7.41b^ 7.0 ± 1.32c 3.8^ ±^ 0.72c Colon-rectum 40.4 ± 8.32a 18.6^ ±^ 4.14b^ 6.1 ± 2.1c 3.3 ± 0.78c Relative weight, g/lOOg body weight Stomach 4.1 ± 1.04a 1.9 ± 0.52° 2.4 ± 0.91° 2.0 ± 0.52° Small intestine Duodenum 0.5^ ±^ 0.09b^ 0.8^ ±^ 0.21a^ 0.4^ ±^ 0.08b^ 0.6^ ±^ 0.24b Jejunum 3.4 ± 0.85거 3.5^ ±^ 0.87a^ 3.2^ ±^ 0.38a^ 1.3^ ±^ 0.22b Ileum 1.0 ± 0.23거 0.7^ ±^ 0.27b^ 1.1^ ±^ 0.25a^ 0.4^ ±^ 0.08c Total weight 4.9 ± 1.08a 5.0^ ±^ 0.96a^ 4.7^ ±^ 0.64a^ 2.3^ ±^ 0.29b Hindgut Caecum 7.8^ ±^ 1.74b^ 9.2 ± 1.70a 2.4^ ±^ O.35c^ 3.2^ ±^ 0.53° Colon-rectum 2.1 ± 0.58° 4.9 ± 0.67a 2.0 ± 0.5 lc 2.8^ ±^ 0.62b Total weight 9.9 ± 2.18b 14.1 ± 1.90a 4.4^ ±^ 0.58d^ 6.0^ ±^ 0.86c Length, cm Duodenum (^) 40.5 ± 3.78a 24.5 ± 9.18° 7.8 ± 2.05c 11.1 ± 3.96c Jejunum 206.1 ± 24.66a 106.5^ ±^ 8.31b^ 93.2 ± 6.86c 21.1^ ±^ 4.89d Ileum 61.6 ± 6.22a 29.5^ ±^ 2.07b^ 25.2^ ±^ 1.94c^ 8.3^ ±^ 0.67d Caecum 63.8 ± 4.49a 11.3^ ±^ 1.16b^ 4.9^ ±^ 0.85d^ 7.3^ ±^ 0.82c Colon-rectum 98.5 ± 9.55a 86.8^ ±^ 4.66b^ 18.8^ ±^ 2.90d^ 34.5^ ±^ 3.08c 1 Mean±SD (n=10). a,b,c,d Means within the (^) dame row without the same superscripts are significantly di仟erent (p<0.05).
1506 CHIOU^ ET^ AL.
increased rate of digesta passage (Sircar et al., 1983). The provision of pectin or lignin in the diet, however, significantly increased epithelial cell multiplication in the intestinal mucosa (Yu and Chiou, 1996). The crude fiber was supplemented using alfalfa and was only 10% in this experiment. This therefore did not result in damaged mucosa in this trial.
CONCLUSION
A diet of 10% dietary fiber or cell wall did not result in damaged mucosa in these growing animals. Rabbits attained the heaviest daily live-weight gain while guinea pigs grew the most rapidly with the best rate of live-weight gain. Rabbits utilized feed significantly more efficiently than rats and hamsters (p<0.05), because rabbits are economical domesticated animals. Guinea pigs also showed better feed conversion rates than rats and hamsters. Rabbits exhibited the longest retention time of digesta. Guinea pigs on the other hand, showed the significantly longest colon-rectum and the highest ability in the digestion of crude fiber, NDF and ADF (p<0.05). Omnivore rats showed significantly poorer digestion of dry matter and fibrous components than the herbivores. This may be attributed to the lighter relative weight of hindgut in rats.
ACKNOWLEGEMENTS
The authors thank the National Science Council of Taiwan for financial support for this project. The project number is NSC 83-0409-B-005-075.
REFERENCES
AOAC. 1980. Official Methods of Analysis. 13th edn. Washington, DC: Association of Official Analyticial Chemists. Brandt, C. S. and E. J. Thacker. 1958. A concept of rate of food passage through the gastro-intestinal tract. J. Anim. Sci. 17:218-223. Chaffee, R. R. J. 1966. On experimental selection for superhibemating and non-hibemating lines of Syrian hamsters. J. Theor. Biol. 12:151-154. Cheeke, P. R. and N. M. Patton. 1980. Carbohydrate overload of the hindgut - a probable cause of enteritis. J. Appl. Rabbit Res. 3:20-23. Ehle, F. R. and R. G. Warner. 1978. Nutritional implications of the hamster forestomach. J. Nutr. 108:1047-1053. Fonnesbeck, P. V., L. E. Harris and L. C. Keral. 1974. Digestion of plant cell walls by animals. J Anim. Sci. 39:182-186. Gidenne, T., R. Bellier and J. van Eys. 1998. Effect of the dietary fiber origin on the digestion and on the caecal fermentation pattern of the growing rabbit. J. Anim. Sci. 66:509-517. Gioffre, F., C. Gisberti, A. Angelini and V. Proto. 1980.
Digestibility in cecotomized rabbits fed different diets. Annali della Facolta di Scienze Agrarie del Universita di Napoli in Portici Serie 14:84-92. Hume, I. D. and E. Sakaguchi. 1991. Patterns of digesta flow and digestion in foregut and hindgut fermenters. In: Physiological Aspects of Digestion and Metabolism in Ruminants (Ed. T. Tsuda, Y. Sasaki and R. Kawashima). Academic Press, Inc. NY. pp. 427-455. Johanning, G. L., D. F. Barr, E. L. lannotti and B. L. ODell. 1984. Effect of dietary fiber on guinea-pig intestinal anaerobe, Bacteroldes ovatus. J. Nutr. 114:354-360. Moore, R. J., E. T. Kornegay, R. L. Grayson and M. D. Lindemann. 1988. Growth, nutrient utilization and intestinal morphology of pigs fed high-fiber diets. J Anim. Sci. 66:1570-1579. Nutrient Requirements of Laboratory Animals. 1995. 4th edn., The National Research Council, Washington, D.C. Nutrient Requirements of Rabbits. 1977. The National Research Council, Washington, DC. Nyman, M. and N. G. Asp. 1985. Dietary fiber fermentation in the rat intestinal tract: effect of adaptation period, protein and fiber levels, and particle size. Br. J. Nutr. 54:635-643. Parker, D.S. and R. T. McMillan. 1976. The determination of volatile fatty acids in the caecum of the conscious rabbit. Br. J. Nutr. 35:365-371. Paulini, I., T. Mehta and A. Hargis. 1987. Intestinal structural changes in African green monkeys after long term psyllium or cellulose feeding. J. Nutr. 117:253-266. Pond, W. G., V H. Varel, J. S. Dickson and W. N. Haschek. 1989. Comparative response of swine and rats to high-fiber or high-protein diets. J. Anim. Sci. 67:716-723. Sakaguchi, E., J. Itoh, S. Uchida and T. Horigome. 1987. Comparison of fiber digestion and digesta retention time between rabbits, guinea pigs, rats and hamster. Br. J. Nutr. 58:149-158. Sakaguchi, E., J. Itoh, H. Shinohara and T. Matsumoto.
- Effects of removal of the forestomach and caecum on the utilization of dietary urea in golden hamster (Mesocrlcetus auratus) given two different diets. Br. J. Nutr. 46:503-512. Sircar, B., L. R. Johnson and L. M. Lichtenberger. 1983. Effect of synthetic diets on gastrointestinal mucosal DNA synthesis in rats. Amer. J. Physiol. 244:G327-335. Slade, L. M. and H. F. Hintz. 1969. Comparison of digestion in horses, ponies, rabbits and guinea pigs. J. Anim. Sci. 28:842-843. Statistical Analysis System. 1985. SAS Users Guide, version
- Cary, SAS Institude Inc, NC. Steel, R. G. D. and J. H. Torrie. 1960. Principles and Procedures of Statistics. McGraw-Hill Book Company, New York. Storme, G. and G. Williams. 1981. The effect of liquid element diet on cell proliferation in the colon of rats. Cell Tissue Res. 216:221-225. Takashi, S. and H. Tamate. 1976. Light and electron microscopic observation of the forestomach mucosa in the golden hamster. Tohoku J. Agric. Res. 27, 26-39. Van Soest, P. J., J. B. Robertson and B. A. Lewis. 1991. Methods for dietary fiber, neutral detergent fiber, and