¡Descarga anatomía de la fruta y más Apuntes en PDF de Entomología solo en Docsity!
UN UNIIVVEERRSSIIDDAADD DDEE PPAANNAAMMAA
PRPROOGGRRAAMMAA DDEE MMAAEESSTTRRIIAA EENN
ENENTTOOMMOOLLOOGGIIAA
VIVICCEERRRREECCTTOORRIIAA DDEE IINNVVEESSTTIIGGAACCIIOONN YY
POPOSSTTGGRRAADDOO
MA MANNUUAALL PPAARRAA LLAA IIDDEENNTTIIFFIICCAACCIIOONN DDEE MMOOSSCCAASS DDEE
LLAA FFRRUUTTAA
GEGENNEERROO AAnnaassttrreepphhaa SCSCHHIINNEERR,, 1 1886688
Cheslavo A. Korytkowski
Agosto 2009
CONTENIDO
- Género Anastrepha SCHINER,
- MORFOLOGIA DE Anastrepha
- CABEZA
- Chaetotaxia
- Apéndices bucales
- Antenas
- TORAX
- Patas
- Alas
- Patrón de coloración de las alas
- ABDOMEN
- Pre-abdomen
- Terminalia del macho
- Terminalia de la hembra
- CONCEPTO DE ESPECIE EN ANASTREPHA
- ORIGEN Y EVOLUCIÓN DEL GÉNERO ANASTREPHA
- UBICACION DE ESPECIES DE Anastrepha POR GRUPOS
- INDICE DE ESPECIES INCLUIDAS
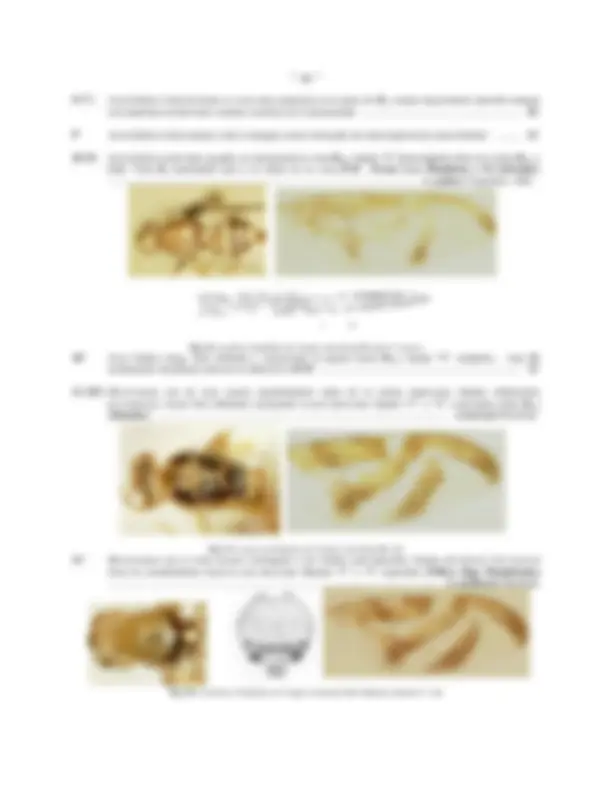
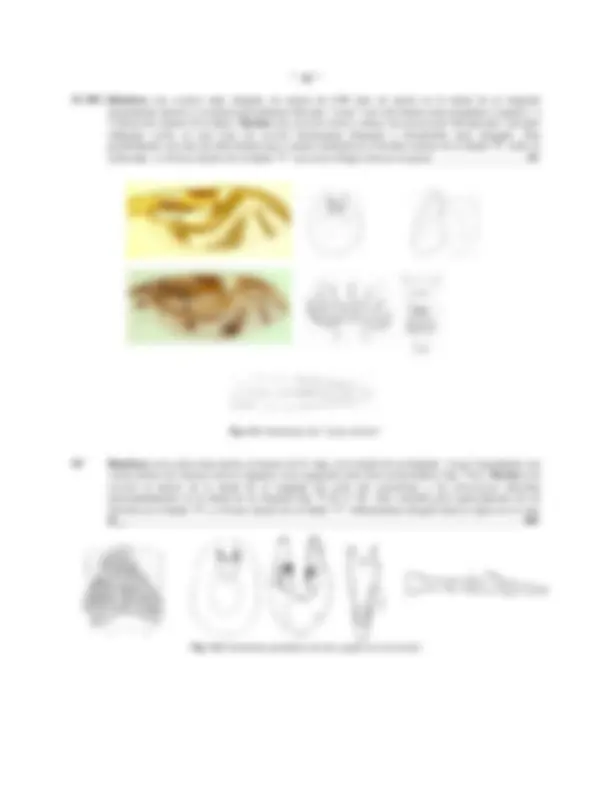
CABEZA DE Anastrepha obliqua: Fig.1: Vista frontal; Fig.2: Vista lateral; Fig.3: Vista caudal; Fig.4: Palpo maxilar (Vista lateral)
CABEZA (Figs. 1 - 3)
Usualmente de forma hemi-esférica o sub-globosa, con el ángulo facial ampliamente obtuso (excepto en A. atrox ), el área post-ocular ventral (debajo de la mitad de la altura ocular) considerablemente mas ancha que la dorsal. Estructuralmente ofrece pocas variaciones, excepto en la facia ( fc , Figs. 1, 2) que posee una carina facial definida en todas las especies, y es usualmente recta, aunque una protuberancia bulbosa puede estar presente en ella debajo de la altura de la base de las antenas (grupo benjamini ) o fuertemente proyectada exactamente entre la base de ellas ( A. atrox ); hacia abajo, la facia se expande moderadamente y tiende a aparecer solo levemente proyectada en un aspecto lateral formando un margen oral ( MO , Fig. 1) débil pero claramente definido, en algunas especies (grupo schausi ), este margen es notoriamente ensanchado (en un aspecto frontal) y usualmente de color mas oscuro o mas claro que el resto de la facia; surcos antenales usualmente bien definidos y angulo vibrissal amplio y obtuso,
con microsetas vibrissales pequeñas y débiles; el triángulo ocelar ( oc , Fig. 1) es pequeño y usualmente de color negro, contrastando con el resto de la cabeza que es uniformemente amarilla o marrón-amarillenta (algunas veces manchas mas oscuras pueden ser observadas en las genas ( gn , Fig. 2), márgenes de los ojos, facia, vertex y post- cranium; area post-vertical ( pcr , Fig. 3), sobre el margen del foramen occipital ( Fo , Fig. 3) con una ruga post- vertical ( rpv , Fig. 3) bien definida.
Chaetotaxia: post-verticales , verticales y ocelares siempre presentes, estas últimas muy reducidas excepto en A. tripunctata donde están bien desarrolladas; dos pares de orbitales superiores ( ors , Figs. 1, 2) reclinadas (solo uno, en el grupo daciformis , ya que el par posterior esta ausente) y 1 a 7 pares de orbitales inferiores ( ori , Figs. 1, 2) inclinadas; Fischer (1934) ha demostrado la variabilidad e inconsistencia de este carácter, ya que aún en un mismo ejemplar puede existir un número diferente a cada lado de la frente. La vitta frontal ( fr vit , Fig. 1) y usualmente también las orbitas ( OR , Figs. 1, 2) están esparcidamente cubiertas de microsetas; setas genales ( ge , Fig. 2) numerosas y pequeñas, usualmente una y raras veces dos posteriores (ubicadas en el ángulo postero-ventral de la cabeza) definidamente mas grandes que el resto; un par de post-genales ( pgn , Fig. 2) bien diferenciadas de las microsetas de ese esclerito; cilia del post-cranium ( poc , Figs. 2, 3) decreciendo uniformemente en tamaño hacia abajo y una hilera bien definida de setas débiles en el extremo posterior (cerca del foramen occipital), debajo de ellas y la cilia post-cranial, numerosas microsetas post-craniales ( c.pcr , Fig. 3) débiles.
Fig.5: Haustellum de Anastrepha obliqua
Los apéndices bucales (Figs. 2, 4, 5) no parecen ofrecer mayores diferencias, aunque en términos generales se puede decir que han sido muy poco estudiados, por otro lado, la coloración de los palpos ( plp , Fig.2, Fig. 9) empleada por algunos autores como carácter descriptivo de algunas especies ( v.g. A. nigripalpis ) parece no ofrecer mucha seguridad, pues el autor ha observado ejemplares de A. striata con estos apéndices oscuros (normalmente claros); la bomba cibo-faringeal ( BF , Fig. 5) puede ser de gran importancia y a parte de Baker et al. (1942), nada se ha trabajado sobre el particular, esta estructura pudiera explicar las modificaciones morfológicas la carina facial y margen oral , observadas en especies de los grupos benjamini y schausi , debido a la inserción de los músculos faringo-clipeales
Antenas : son típicamente anguladas y dirigidas hacia abajo, el post-pedicelum es oval-alargado y se proyecta normalmente hasta 2/3 o 3/4 de la longitud de la carina facial , sin sobrepasar el margen oral , en el grupo daciformis este segmento es estrecho lo que le da un aspecto mas alargado; la arista es micro pubescente y no parece ofrecer mayores características de importancia taxonómica.
TORAX
De aspecto oval hasta oval-alargado, la proporción largo/ancho del scutum puede ser de alguna importancia, las longitudes comparativas presentadas por algunos autores (Stone, 1942a, 1942b) comprenden el scutum ( SC , Figs. 6,
- y scutellum ( SCT , Figs. 6, 7) medidos dorsalmente. Las suturas y escleritos no ofrecen mayores diferencias con
Chaetotaxia: aquí es importante diferenciar las macrosetas (o simplemente setas ) con una base o cicatriz bien definida y las microsetas sin cicatriz o con esta muy reducida; las microsetas pueden ser de gran importancia para la separación práctica de algunas de las especies mas comunes, debido a la coloración muy peculiar (que algunas veces esta ligada estrechamente a su forma, usualmente cónicas, pero anchas y espatuladas o "escamosas" en el grupo striata , que les dan un aspecto "aterciopelado"), las mas importantes son las microsetas del scutum usualmente claras (amarillentas), pero en A. obliqua y muchas "formas" de A. fraterculus las microsetas de las bandas sub- laterales del scutum son negras contrastando con el resto que son amarillentas, el área cubierta por estas microsetas oscuras puede ser muy reducida (unas pocas) y presentes solo en la porción posterior del scutum en otras especies ( p.e. A. distincta , A. perdita ), finalmente la extensión de cobertura por microsetas en algunos escleritos (especialmente del scutellum y anepisternum ), parecen ofrecer algunas evidencia taxonómicas de importancia; en términos generales, la chaetotaxia es poco variable, aunque la ubicación, coloración, tamaño y grosor de algunas setas puede ser de notable importancia en la separación de algunas especies; scutum con dos scapulares ( sca , Figs. 6, 7), la interna usualmente débil y la externa proporcionalmente fuerte, una humeral ( hu , Fig. 6) (= post-pronotal ), dos notopleurales ( ntpl , Fig. 6), una supra-alar presutural ( saps , Fig. 6) y una postsutural ( sapo , Fig. 6), una intra- alar ( ia , Fig. 6), una post-alar ( pa , Fig. 6), una dorso-central ( dc , Fig. 6) y una acrostical ( ac , Fig. 6); scutellum con dos pares de scutellares ( sc , Fig. 6); en la pro-pleura , pro-episternum ( PEP , Fig 7) pubescente, pro-epimeron ( PEPI , Fig. 7) sin setas; en la mesopleura , anepisternum ( ANEI , Fig. 7) sin anepisternales anteriores (aneia) pero con 1-2 anepisternales posteriores ( anei , Fig. 7), anepimeron ( ANEP , Fig. 7) con una anepisternal ( ane , Fig. 7) usualmente bien definida, katepisternum ( KTP , Fig. 7) con una katepisternal ( ktpl , Fig. 7), algunas veces muy reducida (ausente) y no diferenciable de las microsetas katepisternales; algunas de las variaciones mas importantes en la chaetotaxia que se han usado y aún se emplean para la caracterización y diferenciación de especies cor- responden a la presencia y tamaño de la setas scapular mesal (o interna), tamaño y ubicación de las setas acrosticales con relación a las dorsocentrales (usualmente ligeramente detrás de ellas), ubicación de las dorsocentrales con relación a las supra-alares post-suturales (usualmente mas cercanas a las acrosticales que a las supra-alares ), y probablemente uno de los caracteres mas empleados corresponde a la presencia o ausencia y tamaño de la seta katepisternal (mayormente presente y fuerte).
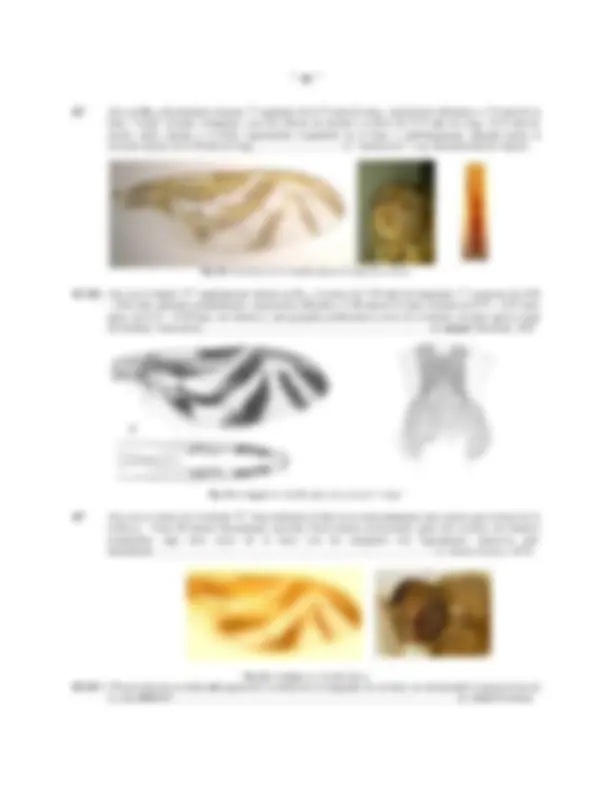
Fig.8: PATRONES DE COLORACION DE Anastrepha****. A: Patrón General; B: A. obliqua ; C: A. shannoni
El color de las macrosetas (o simplemente setas) ha sido y es actualmente empleado, aunque este carácter es poco confiable debido a la gran variabilidad observada en algunas especies, de acuerdo a una categorización cualitativa, las setas pueden ser "negras" u "oscuras", "amarillentas" o "claras", para ello es importante considerar la
edad del ejemplar ya que en individuos tenerales las setas son siempre claras y el estado de preservación (las diferencias de color son mas fáciles de observar en individuos conservados en alcohol que en aquellos montados en seco).
La coloración general (Figs. 8 a, b, c) del cuerpo es usualmente amarilla o marrón-amarillento (algunas veces con un tinte naranja), sin embargo en algunas especies (grupo serpentina ), la coloración es mayormente marrón oscuro hasta casi negro. El scutum cuya coloración general varía de amarillo hasta negro, presenta clásicamente las siguientes áreas claras, generalmente blanquizcas: una banda central (b.c, Fig. 8a)) mas clara, alargada en forma de "fiola" que se proyecta desde la scapula hasta cerca de la sutura scuto-scutellar ( S.scu , Fig. 8a), nace estrecha ade- lante y se expande detrás de la sutura transversa ( s.t , Fig. 8a) en forma mas o menos ovoide (algunas veces cuadran- gular) entre las setas dorsocentrales y acrosticales ; los humeri ( Hu , Fig. 6; H , Fig. 8a), las notopleuras ( NTP , Fig. 6; NTPL , Fig. 8a) y una banda lateral ( b.l. , Fig. 8a)) longitudinal, usualmente proyectada desde la sutura transversa (algunas veces delante de ella) hasta poco antes de la sutura scuto-scutellar entre las setas supra-alares y la intra-alar , estas bandas aunque están generalmente presentes, pueden ser muy difusas y difíciles de diferenciar (especialmente en especímenes secos), algunas veces la "banda central" es realmente ausente. Las áreas comprendidas entre las bandas claras pueden encontrarse oscurecidas hasta negras, especialmente las zonas ubicadas entre la banda central y las dos laterales, formando dos bandas oscuras longitudinales ( b.o.l. , Fig. 8a, 8c), estas pueden ser completas ( A. cordata, A. grandis, A. striata ) abarcando desde el área pre-sutural hasta cerca de la sutura scuto-scutellar o encontrarse divididas ( A. bistrigata ) o aún reducidas a pequeñas manchas de aspecto circular ( A. aczeli, A. punctata, A. tripunctata ) u ovales especialmente en la porción post-sutural; finalmente una banda transversa pre-scutellar ( b.tps , Fig. 8a) oscura de mayor o menor amplitud ( A. sagittata, A. furcata, A. robusta ) puede proyectarse y unir a todas las otras bandas oscuras longitudinales, cerca de la sutura scuto-scutellar en la porción posterior del scutum , o encontrase reducida a una pequeña mancha circular pre-scutellar ( m.c. , Fig. 8a, 8b) ( A. ramosa, A. suspensa, A. spatulata , etc.) que puede ser muy bien definida y negra, difusa y apenas definida, hasta ausente, variando aún dentro de la misma especie (p.e. A. fraterculus ); debajo de la seta post-alar , en la pared lateral entre el scutum y la base del ala hay siempre una mancha supra-alar ( m.s.a. , Fig. 7) negra de aspecto cuadrangular.
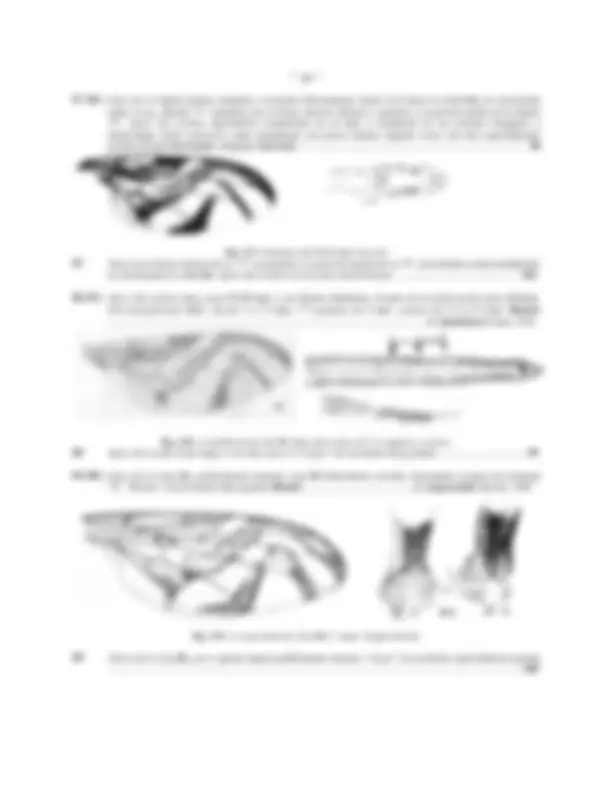
Fig.9: PATAS DE Anastrepha obliqua: A: anterior, B: media, C: posterior
1971), aunque este carácter no ha sido estadísticamente analizado; la longitud de las diferentes porciones de la vena medius fue propuesto por Norrbom y Kim (1985) sin haberse estudiado objetivamente hasta la fecha.
Probablemente el carácter mas estudiado y utilizado para este grupo corresponde al patrón de coloración de las alas , sin embargo, aunque resulta muy consistente y desde luego confiable para algunas especies, en otras (especialmente aquellas ampliamente distribuidas y polífagas) como A. fraterculus, A. ludens y particularmente en A. obliqua este es un carácter extremadamente variable aún dentro de un mismo ejemplar (ala derecha con patrón de color diferente a la izquierda) o entre sexos. El patrón de coloración usualmente de tonalidad amarilla o amarillo naranja con bordes y algunas áreas negruzcas, algunas veces sin embargo mayoritariamente marrón-oscuro, corres- ponde esencialmente a tres bandas: banda costal ( b.c. , Fig. 11) extendida entre el extremo basal del ala hasta el ápice de la vena R 1 sobre el borde anterior del ala y extendiéndose hacia atrás hasta la celda basal medial ( bm , Fig. 10); la banda "S" ( b. "S" , Fig. 11) proyectada en forma curva a modo de una letra "S" desde la celda cubital posterior ( Cub , Fig. 10), subiendo diagonalmente sobre la vena transversa radial-medial ( r-m , Fig. 10), alcanzando el borde anterior del ala en la celda primera radial ( r 1 , Fig. 10) y desde allí continuando sobre el borde anterior hasta el ápice de la tercera celda radial ( r 4+5 , Fig. 10); finalmente la banda "V" ( b."V" , Fig. 10) con aspecto de una letra "V" invertida cuyo brazo interno se proyecta desde el margen posterior del ala hacia adelante sobre la vena transversa distal medial-cubital ( dm-cu , Fig. 10), hasta cerca de o tocando la vena R 4+5 y el brazo externo proyectado desde el borde del ala, detrás de ápice de la vena M hasta tocar o casi tocar el "brazo interno" cerca o en la vena R 4+5 formando el "vértice de la V". Entre las bandas de color, se encuentran áreas claras e hialinas de la membrana alar, entre las cuales la mas importante y frecuentemente empleada es el área hialina costal usualmente de forma triangular, que se encuentra entre la "banda costal" y la "banda S" en el borde anterior del ala, distal al extremo de la vena R 1 , el área hialina costal se puede proyectar a modo de una "banda diagonal hialina" hasta la celda basal medial ( bm , Fig. 10).
Fig.11: ALA DE Anastrepha: Distribución de las bandas de coloración
La amplitud, tonalidad, conexiones y ausencia de las bandas o parte de ellas en el patrón alar son características de gran utilidad para la identificación de especies; algunas veces el patrón es difuso y cubre prácticamente toda el ala, de modo que las bandas son indefinidas (grupo doryphoros ); otras veces, las bandas "costal" y "S" están completamente conectadas en el borde anterior del ala (no existe el área hialina costal) (grupo grandis y grupo daciformis ), la banda "S" puede ser incompleta o discontinua ( A. stonei ), o las bandas "costal" y "S" se encuentran muy ampliamente fusionadas en casi todo el ancho del ala (grupo bivittata).
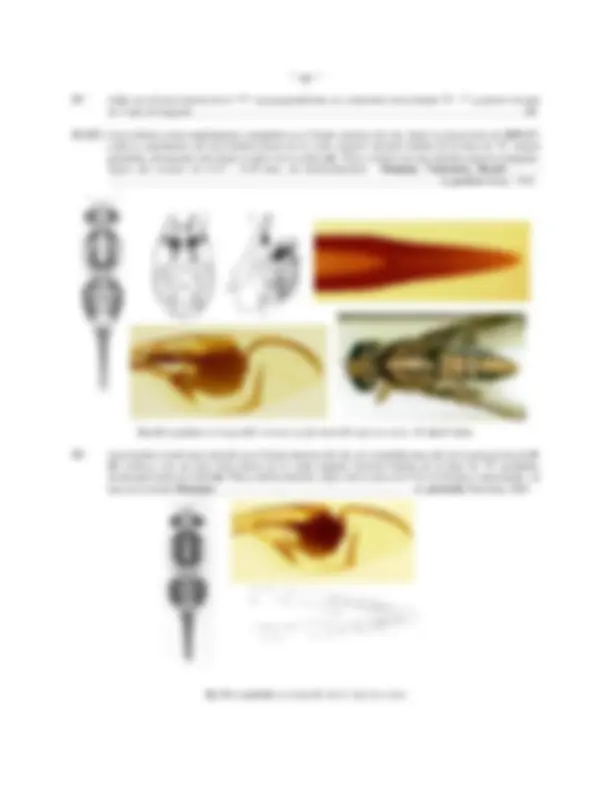
Fig. 12: ABDOMEN DE Anastrepha obliqua: A: Macho, vista dorsal; B: Macho, ventral; C: Hembra, ventral
último es variable en forma y grado de esclerotización pudiendo ofrecer algunas posibilidades taxonómicas, a este se insertan los cerci ( Cer , Figs. 13 a, b, c, d; 14b) membranosos, fusionados mesalmente y densamente pilosos; el arco hipandrial ( Hip , Fig. a, c, e) es relativamente grueso y provisto de un apodema hipandrial ( Aphi , Fig. 13 a, e) corto.
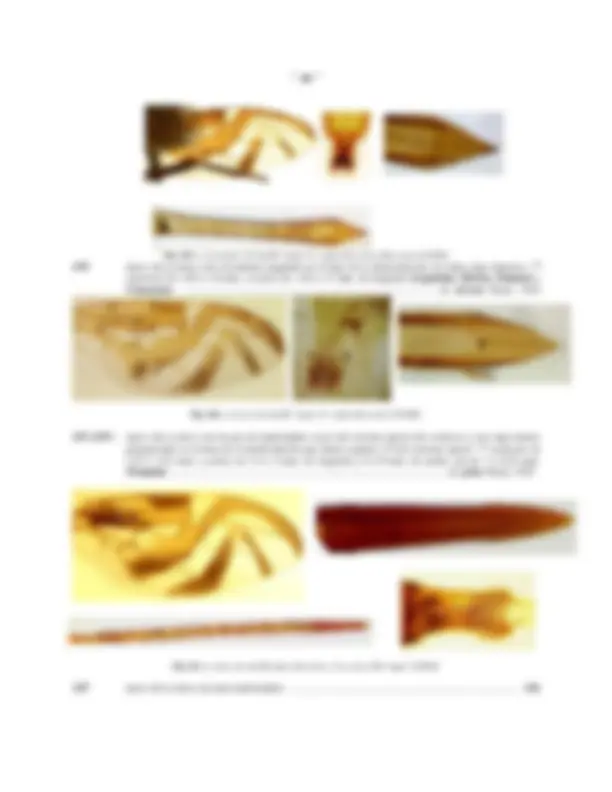
Fig.13: TERMINALIA DEL MACHO, Anastrepha obliqua: A: Andrium y Aedeagus (ventral); B: Cápsula genital (Ventral); C: Andrium, Apodema eyaculador y cerci (lateral); D: Cápsula genital (lateral); E: arco hipandrial y apodema del aedeagus (ventral)
Fig.14: Andrium (órganos perifálicos del macho) de A. dissimilis Stone, 1942; A: ventral; B: lateral
Fig.15: Distiphallus de Anastrepha: A: A. dissimilis Stone; B: A. tecta Zucchi; C: A. manihoti Lima; D: A. pickeli Lima
El aedeagus (Ad) esta conformado por dos tubos fusionados mesalmente denominados paraphalli ( PaPh , Fig 13a), es usualmente largo y recurvado helicoidalmente en la cámara genital (en reposo). excepto en el grupo daciformis , donde es corto y recto; de acuerdo a investigaciones recientes, el aedeagus tiene (en las pocas especies estudiadas) una longitud ligeramente mayor hasta 1 2 veces la longitud del aculeus de la hembra, probablemente debido a que debe alcanzar hasta la vagina de ésta; la porción basal o basiphallus ( Bph , Fig. 13 c, e) es escasamente definida, a modo de un corto anillo esclerosado y se proyecta hacia la cama genital constituyendo el apodema del aedeagus ( A.aed , Fig. 13 a, c, e); el ápice del aedeagus se encuentra ensanchado constituyendo una bolsa eversible o inflable, el distiphallus ( Dph , Figs. 13a; 15 a-d), provisto de una vesícula ventro-basal cubierta de espículas en su
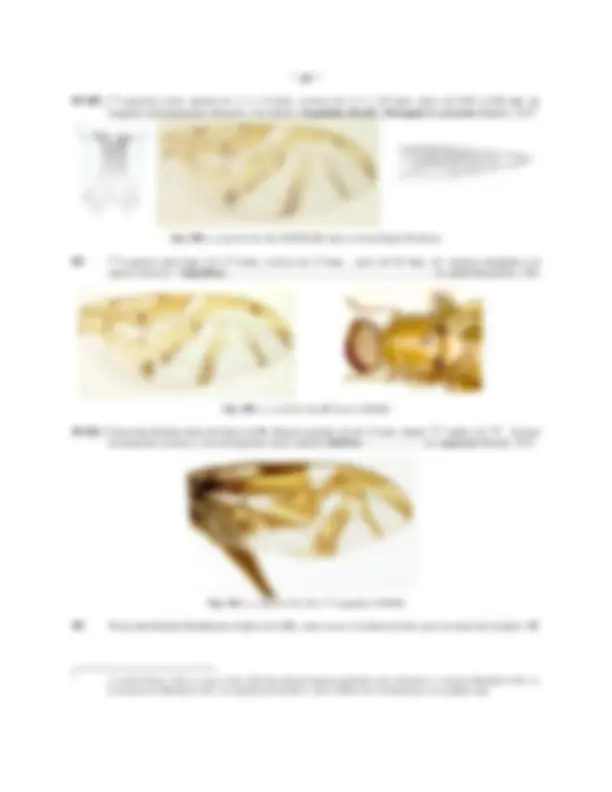
Fig.16: TERMINALIA DE LA HEMBRA DE A. obliqua : A: 7 1 segmento, "raspa" y aculeus (vista dorsal; B: (vista ventral); C: aculeus (dorsal); D: ápice del aculeus (ventral); E: "raspa" ; F: espermatecas
Fig.17: DEFINICION DEL REGISTRO DE MEDIDAS DEL APICE DEL ACULEUS ( Anastrepha obliqua ): A (4-4'): longitud del ápice; B (5-5'): ancho del ápice; C: (6-6') longitud del área denticulada ( "serra" ), (a-a') ancho de la base de la "serra"
El apice ( APA , Figs. 16 C,D; 17) del aculeus , esta constituido por la fusión e integración de los cerci , esta es la estructura mas importante en la identificación de especies del género Anastrepha y en muchos casos la única que permite una identificación "segura", por lo que resulta vital interpretar claramente algunos conceptos relacionados con la forma, características y dimensiones de ella.
La longitud del ápice se mide a lo largo de la línea central (mesal) entre el extremo final del oviducto ( Fiov , Fig. 17 A), generalmente definido por una línea curva, convexa hacia el ápice que delimita una zona mas clara y el extremo del ápice (4 - 4', Fig. 17 A); el ancho se mide a través de la base, cruzando el extremo del oviducto (5 - 5', Fig. 17 B); la proporción largo-ancho corresponde a la división de las dos medidas previas largo/ancho; los margenes pueden ser rectos, cóncavos o convexos y frecuentemente están provistos de una serie de dentículos constituyendo la serra ( SRR , Fig. 17 C), el número, tamaño y forma de los dentículos de la "serra" representan caracteres de notable importancia, estos pueden variar de 3-4 ( A. ramosa ) hasta mas de 100 ( A. leptozona ) a cada lado, extremadamente grandes y toscos ( A. ramosa ) hasta casi imperceptibles (solo visibles a 100 aumentos o mas en A. sodalis, A. mixta ); el ancho de la serra se mide trazando una línea imaginaria que conecte el último diente (el mas basal) de cada lado (a - a', Fig. 17C).
La longitud de la serra desde la línea imaginaria a-a' hasta el extremo final del ápice (6-6', Fig. 17C), la proporción largo/ancho de la serra, así como la proporción entre la serra y el ápice son caracteres de gran utilidad; en algunas especies la "serra" es apenas definida en el extremo apical del ápice (algunos especímenes de A. distincta , A. barbiellini ) y en otros la serra es mas larga que el ápice prolongandose por detrás del extremo del oviducto ( A. umbrosa , A. alveatoides ); finalmente, en los márgenes del ápice, entre la base de la "serra" y del "ápice del aculeus"
el origen alopátrico de otras pareciera ser evidente como sucede en A. ludens (México) y A. distincta (Perú), A. suspensa (Antillas) y A. fraterculus (probablemente la Amazonia); aunque en ciertos casos, la separación de especies (y ciertamente también las asociaciones entre ellas), pueden en cierto grado ser demostradas y bien definidas morfológicamente, en otros, tal situación no puede ser demostrada con la misma facilidad y consistencia.
Los requerimientos sobre información biológica, distribución biogeográficas y ecológica resultan imprescindibles para un mejor entendimiento y aplicación del concepto de especie; mas aún, estudios cariológicos e isoenzimaticos llevados a cabo por algunos autores (Steck, 1991), parecen demostrar la existencia de especies "sibling" (o especies gemelas) dentro del concepto que actualmente poseemos para ciertas "especies" como parece ser el caso de A. obliqua, A. fraterculus, A. ludens y A. distincta , las notables diferencias morfológicas de ciertas poblaciones de A. fraterculus así como sus características de "agresividad", parecieran estar demostrando que las actualmente consideradas "poblaciones" de Brasil, Perú, Ecuador y centroamericanas, así como también las poblaciones centro y sudamericana de A. obliqua , los pocos especímenes conocidos de A. ludens recuperados mas allá de su rango conocido de distribución (Costa Rica y Colombia), las diferencias en el tamaño del 7E tergosternito y ápice del aculeus de A. distincta , pudieran corresponder a evidencias de que mas de una especie esta siendo interpretada erróneamente.
ORIGEN Y EVOLUCIÓN DEL GÉNERO ANASTREPHA
En base a las premisas previamente planteadas, se asume que Toxotrypana y Anastrepha son grupos hermanos y consecuentemente monofiléticos, disgregados de algún ancestro Phytalmiini del Paleantartico.
Siguiendo a Vogt, McGuire y Cushman (1979), podríamos establecer como centro de origen de estos géneros a la cuenca del río de La Plata, en base al número de especies descritas para Toxotrypana (7) (Blanchard, 1959) en áreas aledañas a esa zona, la distribución restringida (Argentina, Paraguay y Brasil) de algunas de las especies mas primitivas del género Anastrepha (v.g. A. daciformis , A. macrura , grupo punctata ) y el gran número de especies descritas de la floresta atlántica del sur de Brasil (reconocida por Haffer, 1974; Vanzolini y Williams, 1970; y Brown et al. , 1974; como uno de los mas importantes refugios faunísticos en el neotropico).
Desde estos refugios la dispersión del genoma podría haber alcanzado rápidamente los límites australes del área de influencia del Amazonas a través del Chaco colonizando los refugios faunísticos de la vertiente atlántica de los Andes hasta los llanos amazónicos de Ecuador; desde allí, por un lado internándose por las cuencas del Cauca y Magdalena en Colombia hasta alcanzar el Darién en Panamá para dispersarse hacia el mas importante refugio de Meso-américa entre el sur de México y Honduras, y de otro lado a través de los llanos colombianos internándose en la cuenca del Orinoco hacia el sur de Venezuela.
Esta teoría podría explicar que la especiación en el género Anastrepha se dio por simpatria (Bush 1966,1969; Futuyma & Mayer 1980) en la mayoría de especies (grupo fraterculus ) con algunas formas de allopatria como se observa en la disgregación de algunas especies del género (v.g. A. ludens ), así como también la prevalencia y adaptación de ciertas especies en zonas de elevada altitud andina que se dispersan hasta las tierras bajas de Brasil y Panamá, con especies caracterizadas por presentar microsetas engrosadas y oscuras (v.g A. cryptostrepha , A. lambda de los andes, A. speciosa en Panamá y A. fenestrata de Brasil y Panamá), presumiblemente las primeras derivadas de las últimas y estas a su vez al parecer de las formas primitivas del "grupo" dentata.
Fig.19: PRESUNTO CENTRO DE ORIGEN (A), REFUGIOS FAUNISTICOS (B-1: MADRE DE DIOS-BENI; B-2: NAPO- AMAZONA; B-3: MACIZO DE GUAYANAS-VENEZUELA; B-4: MACIZO DE YUCATAN) Y PROBABLES RUTAS DE DISPERSION DE Anastrepha.