Vista previa parcial del texto
¡Descarga biblio temas 1-3 (9) y más Apuntes en PDF de Psicología Fisiológica solo en Docsity!
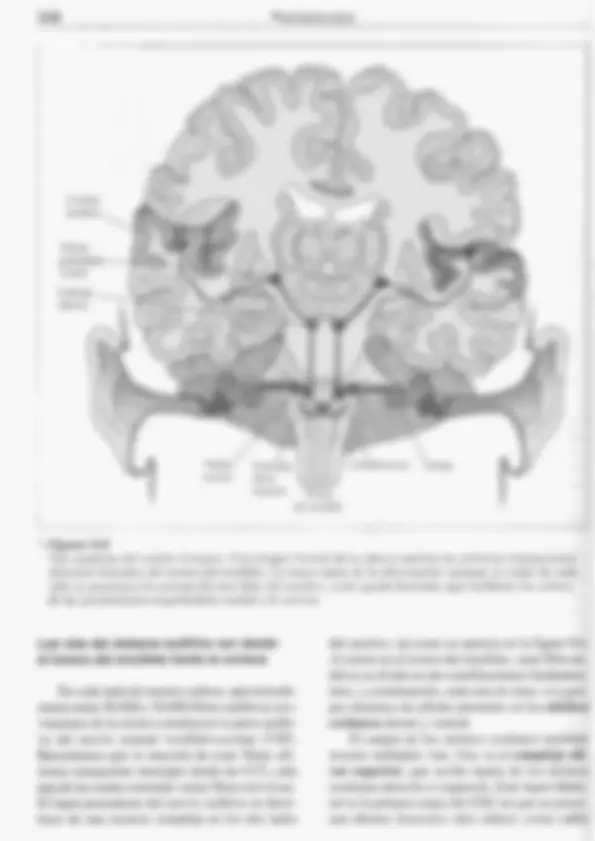
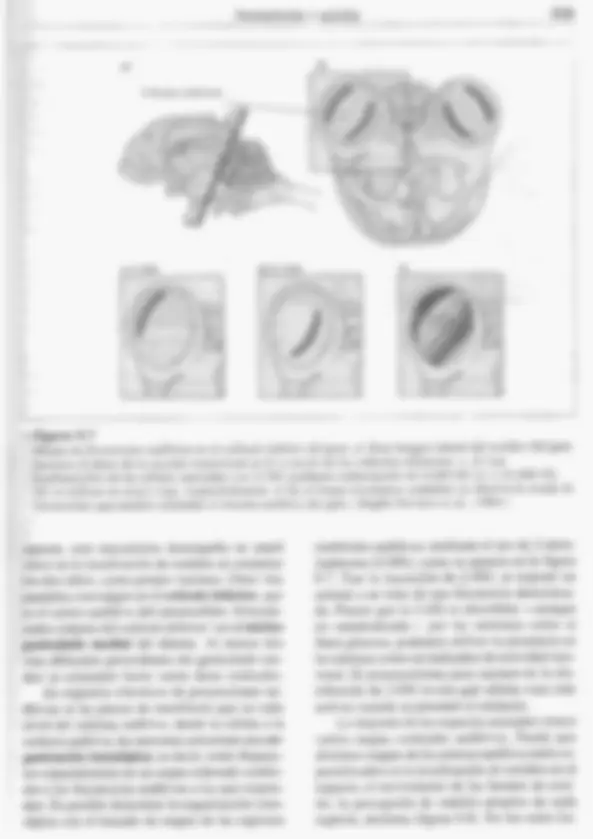
PERCEPCIÓN Y ACCIÓN 335 Las CCE tienen una función diferente: mo- dular la estimulación acústica, Controladas por el SNC mediante sus fibras nerviosas eferentes, las CCE pueden modificar su longitud (Zheng et al.. 2000) y por tanto influir en el mecanismo de la cóclea. Determinados cambios en la longitud de las CCE pueden endurecer o relajar segmen- tos de la membrana basilar y así ajustar activa- mente su sintonía a diferentes frecuencias, como veremos más adelante en este capítulo (Ashmore, 1994), Las CCI también reciben mensajes 'eferentes, seguramente para inhibir parte del input procedente de sonidos fuertes. Ciertos estudios sobre una cepa mutante de rato- nes que carecen de CCT pero tienen CCE nor- males han aportado nuevas pruebas de los dis- tintos papeles de las células ciliadas internas y externas; los ratones citados parecen ser sordos (Deol y Gluecksohn-Waelsch, 1979), Las células ciliadas, ¿cómo convierten el movimiento en actividad neural? Por medio de los mecanismos ya examinados, los sonidos in- ducen vibraciones de la membrana basilar. Estas vibraciones doblan los estereocilios de las células ciliadas que están insertados en la membrana tectorial (véase figura 9.1d). Des- plazamientos muy pequeños de fascículos ci- liados originan cambios rápidos en los canales iónicos de los estereocilios. Estos cambios ini- cian la estimulación de las células ciliadas, y después de los axones aferentes (Hudspeth, 1989, 1992). Se cree que los canales iónicos de los este- reocilios están controlados por energía mecá- nica, como los de los receptores del tacto (véa- se capítulo 8), pero las puertas de los canales para la audición también están especializadas en respuestas rápidas: pueden abrirse y cerrar- se miles de veces por segundo. Esta rapidez descarta el uso de segundos mensajeros, que se hallan en receptores del olfato, el gusto y la vi- sión (como veremos en posteriores apartados de este capítulo y en el 10). Según estimaciones actuales, cada célula ciliada tiene sólo unos cien canales iónicos, aproximadamente uno o dos por estereocilio. Si comparamos registros de diferentes partes de la célula, vemos que algunos de los canales iónicos están cerca de la parte superior de los cilios. Fibras finas, filiformes, denominadas uniones de puntas recorren los extremos de los estereocilios (figura 9,4a) y coordinan me- cánicamente sus movimientos (Eisen et al., 1999), pero también parecen ser elementos cla- ve para generar potenciales de células ciliadas. De acuerdo con ciertos estudios de Hudspeth y sus colegas (1997, 2000). un sonido que balan- cee siquiera ligeramente las células ciliadas in- crementa la tensión en las elásticas uniones de puntas y tira de una «trampilla» abriendo el ca= nal iónico, que se vuelve a cerrar en el espacio de una fracción de milisegundo mientras la cé- lula ciliada efectúa el movimiento oscilatorio de regreso (figura 9.4b y c). La apertura de los canales permite la irrup- ción de iones potasio (K*) y una rápida despo- larización de toda la célula ciliada. Esta des- polarización inicial da lugar a una súbita entra- da de iones calcio (Ca**) en la base de la célula ciliada, debido alo cual las vesículas sinápticas ahí presentes se fusionan con la membrana pre- sináptica y desde ahí liberan su contenido quí- mico —al parecer, el transmisor sináptico glu- tamato—, estimulando la fibra nerviosa afe- rente (figura 9.4c). Los poros iónicos de los cilios son grandes y no diferencian unos ¡ones de otros. Dejan pa- sar incluso moléculas orgánicas pequeñas, mo- tivo por el cual ciertos fármacos como la aspiri- na o los antibióticos pueden dañar las células ciliadas al atacar a sus mitocondrias. La vulne- rabilidad a este tipo de daño se hereda por vía materna, igual que las propias mitocondrias. CIERTOS PROCESOS ELECTROMECÁNICOS ACTIVOS DE.LA CÓCLEA INTENSIFICAN LA DISCRIMINACIÓN DE FRECUENCIAS Los seres humanos son capaces de discri- minar sonidos que difieran en sólo 2 Hz de fre- cuencia, En los primeros estudios sobre curvas de sintonización de fibras nerviosas auditivas individuales, mediante registros eléctricos to- mados del nervio vestíbulo-coclear (VIT), se puso de manifiesto una sintonización mucho más precisa que la observada por Békésy en sus medidas de la sintonización física de la 336 PSICOBIOLOGÍA 2 Figura 9.4 Cómo afecta la estimulación auditiva a los estereocilios de las células ciliadas co- cleares. 4) Esta micrografía de estereocilios muestra las uniones de puntas como estructuras filiformes. b) Hudspeth (1992) sugirió el modelo de estimulación de células ciliadas aquí ilustra- do. c) El desplazamiento de los estereocilios abre los canales de K*. Esta despolarización abre cana- les de:Ca?* en la base de la célula, lo que origina la liberación de neurotransmisor para excitar axo= nes aferentes. (Micrografía, cortesía de A. ].. Hudspeth.) membrana basilar. El hecho de que las caracte- rísticas físicas fundamentales de la membrana basilar no pudieran explicar la agudeza de sin- tonización advertida en el nervio auditivo indi- caba que estaría funcionando un proceso adi- cional de ajuste fino. Hoy sabemos que la pro- pia cóclea posee un sistema electromécánico para incrementar su sintonización. Antes ya indicamos que las CCE exhiben la curiosa propiedad de modificar su longitud cuando son estimuladas eléctricamente: una respuesta electromecánica (Brownell et al., 1985). Con la hiperpolarización, las CCE se alargan; la despolarización las acorta. Estos cambios, que vienen a suponer el 4 % de la lon- gitud de la célula, se producen casi instantá- neamente. Según sugieren los investigadores, estas respuestas mecánicas de las CCE funcio- nan como un amplificador coclear, que am- plía los movimientos de la membrana basilar en unas regiones y los amortigua en otras. Esta modulación activa y progresiva de la membra- na basilar ajusta la sintonización de la cóclea (Ashmore, 1994; Hubbard, 1993). Pero mientras que cada neurona responde a una frecuencia muy precisa en su umbral, a medida que se usan estímulos más intensos la neurona responde a una escala más amplia de frecuencias. Por ejemplo, la fibra cuyas res- puestas aparecen en rojo en la figura 9.5 tiene su mejor frecuencia en 1.200 Hz; esto es, res- ponde a un tono muy débil de 1.200 Hz. No obstante, si los sonidos son 20 dB más fuertes, la fibra responde a cualquier frecuencia com- prendida entre 500 y 1.800 Hz. Así pues, aun- que una fibra auditiva nerviosa transmite €x- clusivamente información auditiva, no respon- de sólo a una frecuencia de estimulación, Si el cerebro recibiera una señal procedente de una fibra así, no sería capaz de determinar si la esti- mulación es un tono débil de 1.200 Hz o uno más fuerte de 500 o 1.800 Hz o de cualquier otra frecuencia intermedia. Como propuso también Békésy, la capaci- 338 PSICOBIOLOGÍA Corteza auditiva Núcleo geniculado medial Colículo inferior Complejo coclear olivar superior Tronco Protuberancia — Cóclea del encéfalo » Figura 9,6 Vías auditivas del cerebro humano. Esta imagen frontal de la cabeza muestra las primeras interacciones aferentes biaurales del tronco del encéfalo. La mayor parte de la información (aunque no toda) de cada oído se proyecta a la corteza del otro lado del cerebro, como queda ilustrado aquí mediante los colores de las proyecciones al geniculado medial y la corteza. Las vías del sistema auditivo van desde el tronco del encéfalo hasta la corteza En cada lado.de nuestra cabeza, aproximada- mente entre 30.000 y 50.000 fibras auditivas pro- venientes de la cóclea constituyen la parte auditi- va del nervio craneal vestíbulo-coclear (VID. Recordemos que la mayoría de estas fibras afe- rentes transportan mensajes desde las CCI, cada una de las cuales estimula varias fibras nerviosas. El input procedente del nervio auditivo se distri- buye de una manera compleja en los dos lados del cerebro, tal como se aprecia en la figura 9.6. Al entrar en el tronco del encéfalo, cada fibra au- ditiva se divide en dos ramificaciones fundamen- tales; a continuación, cada una de éstas va a gru- pos distintos de células presentes en los núcleos cocleares dorsal y ventral. El output de los núcleos cocleares también recorre múltiples vías, Una va al complejo oli- var superior, que recibe inputs de los núcleos cocleares derecho e izquierdo. Este input bilate- ral es la primera etapa del SNC en que se proce- san efectos biaurales (dos oídos); como cabía PERCEPCIÓN Y ACCIÓN 339 a) Colículos inferiores €) 2 kHz d) 21 kHz > Figura 9.7 Mapas de frecuencias auditivas en el colículo inferior del gato. a) Esta imagen lateral del cerebro del gato muestra el plano de la sección transversal en b) a través de los colículos inferiores. e, d) Las localizaciones de las células marcadas con 2-DG mediante estimulación de 2.000 Hz (c) y 21.000 Hz (d) se indican en azul y rojo, respectivamente. e) En el mapa tonotópico completo se observa la escala de frecuencias que pueden estimular el sistema auditivo del gato, (Según Serviere et al., 1984.) esperar, este mecanismo desempeña un papel clave en la localización de sonidos al comparar los dos oídos, como pronto veremos. Otras vías paralelas convergen en el colículo inferior, que es el centro auditivo del mesencéfalo. Determi- nados outputs del colículo inferior van al núcleo geniculado medial del tálamo. Al menos dos vías diferentes procedentes del geniculado me- dial se extienden hasta varias áreas corticales. En registros eléctricos de proyecciones au- ditivas se ha puesto de manifiesto que en cada nivel del sistema auditivo, desde la cóclea a la corteza auditiva, las neuronas presentan una 0r- ganización tonotópica; es decir, están dispues- tas espacialmente en un mapa ordenado confor- me a las frecuencias auditivas a las que respon- den. Es posible demostrar la organización tono- tópica con el trazado de mapas de las regiones cerebrales auditivas mediante el uso de 2-deso- xiglucosa (2-DG), como se aprecia en la figura 9.7. Tras la inyección de 2-DG, se expone un animal a un tono de una frecuencia determina- da. Puesto que la 2-DG es absorbida —aunque no metabolizada— por las neuronas como si fuera glucosa, podemos utilizar su presencia en las mismas como un indicador de actividad neu- ronal, El procesamiento post mortem de la dis- tribución de 2-DG revela qué células eran más activas cuando se presentó el estímulo. La mayoría de las especies animales tienen varios mapas corticales auditivos. Puede que distintos mapas de la corteza auditiva estén es- pecializados en la localización de sonidos en el espacio, el movimiento de las fuentes de soni- do, la percepción de sonidos propios de cada especie, etcétera (figura 9.8). En los seres hu- PERCEPCIÓN Y ACCIÓN 341 a) Sujeto 1 Sonidos del habla | s | : 5 2 E 3 4 5 L Sujeto 2 > Figura 9.9 Respuestas de la corteza auditiva humana a sonidos aleatorios versus el habla. a) Escáneres de RMf de los hemisferios cerebrales muestran que el ruido:o los tonos puros (arriba) activan principalmente el área auditiva primaria de la parte superior del lóbulo temporal, mientras que escuchar sonidos del habla (abajo) activa otras regiones corticales auditivas además del área auditiva primaria. b) Imágenes de TEP lateral (izquierda) y horizontales (derecha) ponen de manifiesto que escuchar palabras activa no sólo diversas regiones de la corteza cerebral sino también regiones del tálamo y el cerebelo. Las líneas horizontales numeradas de la izquierda corresponden a los niveles de las cortes horizontales de la derecha, (Parte a, de Binder et al., 1994, cortesía de Jefhi cortesía de Marcus Raichle.) por segundo. En ambos casos, la descarga del impulso nervioso está sincronizada —con- cuerda— con el estímulo; esto es, se produce en uña parte concreta del ciclo, Varias fibras pueden llevar a cabo una representación así de sincronización con la fase con mayor precisión que una sola: de ahí el término «descarga», como aludiendo a una descarga cerrada, una lluvia de impulsos nerviosos. ¿Estos enfoques —las teorías del lugar y de la descarga— son forzosamente antagonistas? No. De hecho, la idea actual de la percepción del tono incorpora ambas perspectivas. Tal como predice la teoría del lugar, un cam- bio en la frecuencia va acompañado por un cambio en la región de máxima alteración de la membrana basilar, así como por la activación de receptores auditivos conexos. Para los sonidos complejos con componentes de diferentes fre- Binder; b, de Posner y Raichle, 1994, y cuencias, la cóclea lleva a cabo una especie de análisis de Fourier (véase recuadro 9.1), con las distintas frecuencias representadas como valores máximos de vibración en diferentes lugares de la membrana basilar. La precisión de la representa- ción de lugar de la frecuencia auditiva ha mejora- do a lo largo de la evolución a medida que se ha alargado la membrana y ha aumentado el número de células ciliadas y fibras nerviosas auditivas, Tal como pronostica la teoría de la descarga, los patrones temporales de las descargas neura- les parecen codificar frecuencias auditivas. Se- gún ciertos registros directos, para sonidos de frecuencia baja, la frecuencia de los potenciales de acción a menudo codifica la frecuencia audi- tiva en base a una correspondencia unívoca, Para sonidos de frecuencia alta, la frecuencia de los potenciales de acción puede, en cambio, co- dificar una frecuencia auditiva que es un múlti- 342 PSICOBIOLOGÍA plo de número entero. La codificación de una frecuencia múltiple, más que una de carácter unívoco, permite al sistema auditivo superar la tasa máxima de descarga neuronal de aproxima- damente 1.000 potenciales de acción por segun- do. En general, la codificación de la descarga se pone de relieve en el extremo inferior de la esca- la auditiva. hasta unos 4.000 Hz. Así pues, las propiedades de un sonido se codifican de dos maneras: (1) con arreglo a la distribución de la excitación entre las células —esto es, la codificación del lugar o la repre- sentación tonotópica—, y (2) conforme al pa- trón temporal de descarga en células que se ex- tienden desde el nervio auditivo hasta la corte- za auditiva. Algunas especies son sensibles a sonidos con frecuencias muy altas (ultrasónicos) o muy bajas (infrasónicos), y aprovechan esto me- diante estrategias concretas, Por ejemplo, mu- chas especies de murciélagos producen vocali- záciones fuertes, en la escala 50.000-100.000 Hz, y escuchan los ecos reflejados desde los ob- jetos a fin de desplazarse y cazar en la oscuri- dad. Deben su extraordinaria sensibilidad y pre- cisión a complejas adaptaciones que potencian su cuerpo y su cerebro. Enel otro extremo del espectro, se ha obser- “vado que ciertas cepas de palomas mensajeras quedan gravemente trastornadas si las alcanza el estruendo del Concorde al romper la barrera del sonido (una pulsación infrasónica que puede desplazarse centenares de kilómetros), lo que sugiere que las palomas se valen de las señales infrasónicas para trazar un mapa de navegación (Hagstrum, 2000). Puede que los tigres utilicen infrasonidos para añadir impacto a sus rugidos (Walsh er al.. 2003); también ciertos infrasoni- dos introducidos experimentalmente en con- ciertos elevan el efecto emocional de la música en los asistentes; de todos modos, todavía no se sabe cómo los detectamos, Podemos localizar sonidos comparando uno y otro oído Normalmente podemos localizar la posi- ción de una fuente de sonido con gran precisión (con un margen de error de aproximadamente 19) analizando diferencias biaurales (dos of- dos). La localización de una fuente sonora vie- ne indicada por dos tipos de señales biaurales. Las diferencias de intensidad son diferen- cias de volumen en los dos oídos. En los seres humanos, se producen diferencias de intensk dad porque la cabeza proyecta una sombra de sonido, lo que impide que los sonidos de un lado (desplazados respecto al eje) lleguen a los dos oídos con igual intensidad (esto es un fac= tormenos importante en especies con una colo= cación distinta de los oídos, como los perros y los gatos). La sombra de la cabeza:es más acu sada con los sonidos de frecuencia más ele- vada, Los sonidos de baja frecuencia tienen on= das sonoras más largas que circulan alrededor de la cabeza. Las diferencias de latencia son las que existen entre los dos oídos en cuanto al tiempo de llegada de los sonidos. Se producen porque un oído está siempre algo más cerca que el otro de un sonido desplazado respecto al eje. En un sonido hay presentes dos tipos de diferencias de latencia: la disparidad de inicio, que es la diferencia entre los dos oídos al oír el comien- zo de un sonido, y la disparidad de fase en cur- so, que es el desparejamiento continuo entre los dos oídos en cuanto a la llegada de todos los valores máximos y mínimos (picos y valles) que constituyen la onda sonora (en la figura 9.10 aparecen ¡ilustradas estas indicaciones). Actualmente sabemos que la localización de sonidos supone procesamiento de diferen- cias tanto de intensidad como de latencia, lo que se conoce como teoría dúplex. A frecuen- cias bajas. con independencia del acimut con que se presenten los sonidos (horizontalmente alrededor de la cabeza), prácticamente no se aprecian diferencias de intensidad entre los oí- dos. Para estas frecuencias, las diferencias en los tiempos de llegada son las principales indi- caciones de la posición del sonido (y a frecuen- cias muy bajas ninguna señal ayuda demasia- do; por esto podemos colocar el bafle de los ba- jos donde queramos). No obstante, a frecuen- cias más altas, la sombra de sonido proyectada por la cabeza genera importantes diferencias en cuanto a la intensidad biaural. Naturalmen-