¡Descarga Cap46 - Capitulo 46 de Guyton y más Transcripciones en PDF de Fisiología solo en Docsity!
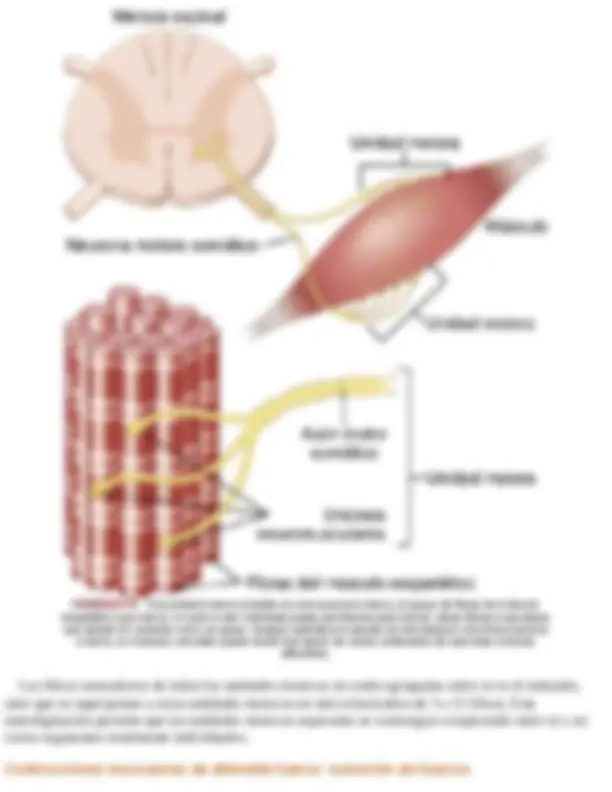
Contracción del músculo esquelético
Aproximadamente el 40% del cuerpo es músculo esquelético, y tal vez otro 10% es músculo liso y cardíaco. Algunos de los principios básicos de la contracción se aplican también a los diferentes tipos de músculos. En este capítulo se considera principalmente la función del músculo esquelético; las funciones especializadas del músculo liso se analizan en el capítulo 8 y el músculo cardíaco se analiza en el capítulo 9.
Anatomía fisiológica del músculo esquelético
Fibras del músculo esquelético
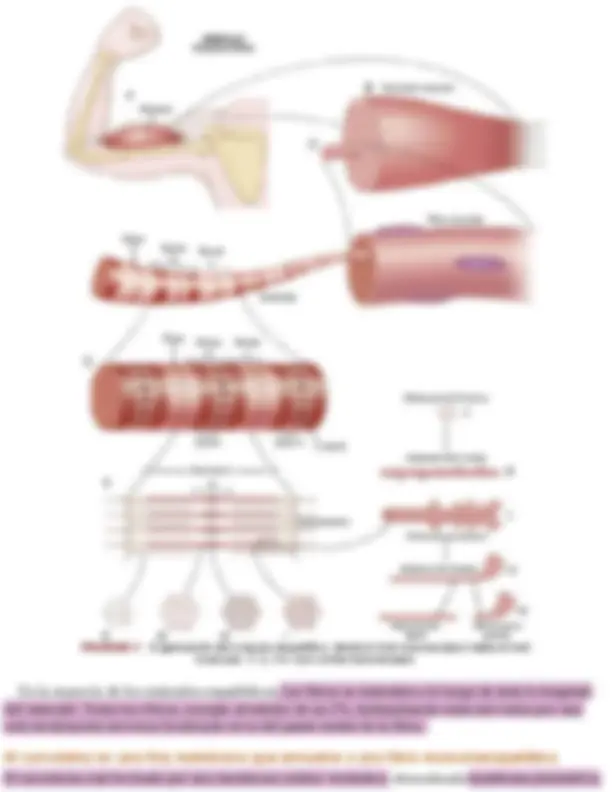
La figura 6-1 ilustra la organización del músculo esquelético y muestra que todos los músculos esqueléticos están formados por numerosas fibras cuyo diámetro varía entre 10 y 80 μm. Cada una de estas fibras está formada por subunidades cada vez más pequeñas, que también se muestran en la figura 6-1 y que se describen en los párrafos siguientes.
y una cubierta externa formada por una capa delgada de material polisacárido que contiene numerosas fibrillas delgadas de colágeno. En cada uno de los dos extremos de la fibra muscular la capa superficial del sarcolema se fusiona con una fibra tendinosa. Las fibras tendinosas a su vez se agrupan en haces para formar los tendones musculares, que después insertan los músculos en los huesos.
Las miofibrillas están formadas por filamentos de actina y miosina
Cada fibra muscular contiene varios cientos a varios miles de miofibrillas, que se representan en la vista en sección transversal de la figura 6-1 C. Cada miofibrilla (v. fig. 6-1 D y E ) está formada por aproximadamente 1.500 filamentos de miosina y 3.000 filamentos de actina adyacentes entre sí, que son grandes moléculas proteicas polimerizadas responsables de la contracción muscular real. Estos filamentos se pueden ver en una imagen longitudinal en la microfotografía electrónica de la figura 6- 2 y se representan esquemáticamente en la figura 6-1 , partes E a L. Los filamentos gruesos de los diagramas son miosina y los filamentos delgados son actina.
FIGURA 6-2 Microfotografía electrónica de las miofibrillas musculares que muestra la organización detallada de los filamentos de actina y miosina. Obsérvense las mitocondrias situadas entre las miofibrillas. (Tomado de Fawcett DW: The Cell. Philadelphia: WB Saunders, 1981.)
Obsérvese en la figura 6-1 E que los filamentos de miosina y de actina se interdigitan parcialmente y de esta manera hacen que las miofibrillas tengan bandas claras y oscuras alternas, como se ilustra en la figura 6-2. Las bandas claras contienen solo filamentos de actina y se denominan bandas I porque son isótropas a la luz polarizada. Las bandas oscuras contienen filamentos de miosina, así como los extremos de los filamentos de actina en el punto en el que se superponen con la miosina, y se denominan bandas A porque son anisótropas a la luz polarizada. Obsérvense también las pequeñas
proyecciones que se originan en los lados de los filamentos de miosina en la figura 6-1 E y L, y que se denominan puentes cruzados. La interacción entre estos puentes cruzados y los filamentos de actina produce la contracción. La figura 6-1 E también muestra que los extremos de los filamentos de actina están unidos al disco Z. Desde este disco estos filamentos se extienden en ambas direcciones para interdigitarse con los filamentos de miosina. El disco Z, que está formado por proteínas filamentosas distintas de los filamentos de actina y miosina, atraviesa las miofibrillas y también pasa desde unas miofibrillas a otras, uniéndolas entre sí a lo largo de toda la longitud de la fibra muscular. Por tanto, toda la fibra muscular tiene bandas claras y oscuras, al igual que las miofibrillas individuales. Estas bandas dan al músculo esquelético y cardíaco su aspecto estriado. La porción de la miofibrilla (o de la fibra muscular entera) que está entre dos discos Z sucesivos se denomina sarcómero. Cuando la fibra muscular está contraída, como se muestra en la parte inferior de la figura 6-5 , la longitud del sarcómero es de aproximadamente 2 μm. Cuando el sarcómero tiene esta longitud, los filamentos de actina se superponen completamente con los filamentos de miosina y las puntas de los filamentos de actina están comenzando ya a superponerse entre sí. Como se expone más adelante, a esta longitud el músculo es capaz de generar su máxima fuerza de contracción.
Las moléculas filamentosas de titina mantienen en su lugar los filamentos de miosina y actina
La relación de yuxtaposición entre los filamentos de miosina y de actina se mantiene por medio de un gran número de moléculas filamentosas de una proteína denominada titina (fig. 6-3). Cada molécula de titina tiene un peso molecular de aproximadamente 3 millones, lo que hace que sea una de las mayores moléculas proteicas del cuerpo. Además, como es filamentosa, es muy elástica. Estas moléculas elásticas de titina actúan como armazón que mantiene en su posición los filamentos de miosina y de actina, de modo que funcione la maquinaria contráctil del sarcómero. Un extremo de la molécula de titina es elástico y está unido al disco Z; para actuar a modo de muelle y con una longitud que cambia según el sarcómero se contrae y se relaja. La otra parte de la molécula de titina la une al grueso filamento de miosina. La molécula de titina también parece actuar como molde para la formación inicial de porciones de los filamentos contráctiles del sarcómero, especialmente los filamentos de miosina.
FIGURA 6-4 Retículo sarcoplásmico en los espacios extracelulares que hay entre las miofibrillas, que muestra un sistema longitudinal que sigue un trayecto a las miofibrillas. También se muestran en sección transversal los túbulos T (flechas) que se dirigen hacia el exterior de la membrana de la fibra y que participan en la transmisión de la señal eléctrica hacia el centro de la fibra muscular. (Tomado de Fawcett DW: The Cell. Philadelphia: WB Saunders, 1981.)
Mecanismo general de la contracción muscular
El inicio y la ejecución de la contracción muscular se producen en las siguientes etapas secuenciales:
- Un potencial de acción viaja a lo largo de una fibra motora hasta sus terminales sobre las fibras musculares.
- En cada terminal, el nervio secreta una pequeña cantidad de la sustancia neurotransmisora acetilcolina.
- La acetilcolina actúa en una zona local de la membrana de la fibra muscular para abrir múltiples canales de cationes «activados por acetilcolina» a través de moléculas proteicas que flotan en la membrana.
- La apertura de los canales activados por acetilcolina permite que grandes cantidades de iones sodio difundan hacia el interior de la membrana de la fibra muscular. Esta acción provoca una despolarización local que, a su vez, conduce a la apertura de los canales de sodio activados por el voltaje, que inicia un potencial de acción en la membrana.
- El potencial de acción viaja a lo largo de la membrana de la fibra muscular de la misma manera que los potenciales de acción viajan a lo largo de las membranas de las fibras nerviosas.
- El potencial de acción despolariza la membrana muscular, y buena parte de la electricidad del potencial de acción fluye a través del centro de la fibra muscular, donde hace que el retículo sarcoplásmico libere grandes cantidades de iones calcio que se han almacenado en el interior de este retículo.
- Los iones calcio inician fuerzas de atracción entre los filamentos de actina y miosina, haciendo que se deslicen unos sobre otros en sentido longitudinal, lo que constituye el proceso contráctil.
- Después de una fracción de segundo los iones calcio son bombeados de nuevo hacia el retículo sarcoplásmico por una bomba de Ca++^ de la membrana y permanecen almacenados en el retículo hasta que llega un nuevo potencial de acción muscular; esta retirada de los iones calcio desde las miofibrillas hace que cese la contracción muscular. A continuación describimos la maquinaria molecular del proceso de la contracción muscular.
rodean rápidamente a las miofibrillas. A su vez, los iones calcio activan las fuerzas de atracción entre los filamentos de miosina y de actina y comienza la contracción. Sin embargo, es necesaria energía para que se realice el proceso contráctil. Esta energía procede de los enlaces de alta energía de la molécula de ATP, que es degradada a difosfato de adenosina (ADP) para liberarla. En las siguientes secciones describimos estos procesos moleculares de la contracción.
Características moleculares de los filamentos contráctiles
Los filamentos de miosina están compuestos por múltiples moléculas de miosina
Cada una de las moléculas de miosina, mostradas en la figura 6-6 A, tiene un peso molecular de aproximadamente 480.000. La figura 6-6 B muestra la organización de muchas moléculas para formar un filamento de miosina, así como la interacción de este filamento por un lado con los extremos de dos filamentos de actina.
FIGURA 6-6 A. Molécula de miosina. B. Combinación de muchas moléculas de miosina para formar un filamento de miosina. También se muestran miles de puentes cruzados de miosina y la interacción entre las cabezas de los puentes cruzados con los filamentos de actina adyacentes.
La molécula de miosina (v. fig. 6-6 A ) está formada por seis cadenas polipeptídicas, dos cadenas pesadas, cada una de las cuales tiene un peso molecular de aproximadamente 200.000, y cuatro cadenas ligeras, que tienen un peso molecular de aproximadamente 20.000 cada una. Las dos cadenas pesadas se enrollan entre sí en espiral para formar una hélice doble, que se denomina cola de la molécula de miosina. Un extremo de cada una de estas cadenas se pliega bilateralmente para formar una estructura polipeptídica globular denominada cabeza de la miosina. Así, hay dos cabezas libres en un extremo de la molécula de miosina de doble hélice. Las cuatro cadenas ligeras también forman parte de la cabeza de la miosina, dos en cada cabeza. Estas cadenas ligeras ayudan a controlar la función de la cabeza durante la contracción muscular. El filamento de miosina está formado por 200 o más moléculas individuales de miosina. En la figura 6-6 B se muestran la porción central de uno de estos filamentos y las colas de las moléculas de miosina agrupadas entre sí para formar el cuerpo del filamento, mientras que hay muchas cabezas de las moléculas por fuera de los lados del cuerpo. Además, parte del cuerpo de cada una de las moléculas de miosina se prolonga hacia la región lateral junto a la cabeza, formando de esta manera un brazo que separa la cabeza del cuerpo, como se muestra en la figura. Los brazos y las cabezas que
hebras de F-actina están escalonados, lo que permite que haya un punto activo en toda la longitud del filamento de actina cada 2,7 nm. Cada uno de los filamentos de actina tiene una longitud de aproximadamente 1 μm. Las bases de los filamentos de actina se anclan fuertemente en los discos Z; los extremos de los filamentos protruyen en ambas direcciones para situarse en los espacios que hay entre las moléculas de miosina, como se muestra en la figura 6-.
Moléculas de tropomiosina
El filamento de actina también contiene otra proteína, la tropomiosina. Cada molécula de tropomiosina tiene un peso molecular de 70.000 y una longitud de 40 nm. Estas moléculas están enrolladas en espiral alrededor de los lados de la hélice de F-actina. En estado de reposo las moléculas de tropomiosina recubren los puntos activos de las hebras de actina, de modo que no se puede producir atracción entre los filamentos de actina y de miosina para producir la contracción.
Troponina y su función en la contracción muscular
Unidas de manera intermitente a lo largo de los lados de las moléculas de tropomiosina hay otras moléculas proteicas denominadas troponina. Estas moléculas proteicas son en realidad complejos de tres subunidades proteicas unidas entre sí de manera laxa, cada una de las cuales tiene una función específica en el control de la contracción muscular. Una de las subunidades (troponina I) tiene una gran afinidad por la actina, otra (troponina T) por la tropomiosina y la tercera (troponina C) por los iones calcio. Se cree que este complejo une la tropomiosina a la actina y que la intensa afinidad de la troponina por los iones calcio inicia el proceso de la contracción, como se explica en la sección siguiente.
Interacción de un filamento de miosina, dos filamentos de
actina y los iones calcio para producir la contracción
Inhibición del filamento de actina por el complejo troponina-tropomiosina
Un filamento de actina puro sin la presencia del complejo troponina-tropomiosina (pero en presencia de iones magnesio y ATP) se une instantánea e intensamente a las cabezas de las moléculas de miosina. Después, si se añade el complejo troponina-tropomiosina al filamento de actina, no se produce la unión entre la miosina y la actina. Por tanto, se piensa que los puntos activos del filamento de actina normal del músculo relajado son inhibidos o cubiertos físicamente por el complejo troponina-tropomiosina. En consecuencia, estos puntos no se pueden unir a las cabezas de los filamentos de miosina para producir la contracción. Antes de que se produzca la contracción, se debe inhibir el efecto bloqueante del complejo troponina-tropomiosina.
Activación del filamento de actina por iones calcio
En presencia de grandes cantidades de iones calcio, se inhibe el propio efecto inhibidor del complejo troponina-tropomiosina sobre los filamentos de actina. No se conoce el mecanismo de esta inhibición, aunque una hipótesis es la siguiente: cuando los iones calcio se combinan con la troponina C, de la que una molécula se puede unir intensamente con hasta cuatro iones calcio, el complejo de troponina probablemente experimenta un cambio conformacional que en cierto modo tira de la molécula de tropomiosina y la desplaza hacia zonas más profundas del surco que hay entre las dos
hebras de actina. Esta acción «descubre» los puntos activos de la actina, permitiendo de esta manera que atraigan a las cabezas del puente cruzado de miosina y que produzcan la contracción. Aunque este mecanismo es hipotético, pone de relieve que la relación normal entre el complejo troponina- tropomiosina y la actina es alterada por los iones calcio, dando lugar a una nueva situación que lleva a la contracción.
Interacción entre el filamento de actina «activado» y los puentes cruzados de miosina: teoría de la «cremallera» de la contracción
Tan pronto como el filamento de actina es activado por los iones calcio, las cabezas de los puentes cruzados de los filamentos de miosina son atraídos hacia los puntos activos del filamento de actina y de algún modo esto hace que se produzca la contracción. Aunque el mecanismo preciso mediante el que esta interacción entre los puentes cruzados y la actina produce la contracción sigue siendo en parte teórico, una hipótesis para la que hay datos considerables es la teoría de la «cremallera» (o teoría del « trinquete ») de la contracción. La figura 6-8 muestra este hipotético mecanismo de la cremallera de la contracción. La figura muestra las cabezas de los puentes cruzados uniéndose y liberándose de los puntos activos de un filamento de miosina. Cuando una cabeza se une a un punto activo, esta unión produce simultáneamente cambios profundos en las fuerzas intramoleculares entre la cabeza y el brazo de este puente cruzado. La nueva alineación de las fuerzas hace que la cabeza se desplace hacia el brazo y que arrastre con ella al filamento de actina. Este desplazamiento de la cabeza se denomina golpe activo. Inmediatamente después del desplazamiento, la cabeza se separa automáticamente del punto activo; a continuación la cabeza recupera su dirección extendida. En esta posición se combina con un nuevo punto activo que está más abajo a lo largo del filamento de actina; después la cabeza se desplaza una vez más para producir un nuevo golpe activo, y el filamento de actina avanza otro paso. Así, las cabezas de los puentes cruzados se incurvan hacia atrás y hacia delante y paso a paso recorren el filamento de actina, desplazando los extremos de dos filamentos de actina sucesivos hacia el centro del filamento de miosina.
FIGURA 6-8 Mecanismo de «cremallera» de la contracción muscular.
músculo en contracción
La figura 6-9 muestra el efecto de la longitud del sarcómero y de la cantidad de la superposición entre los filamentos de miosina y de actina sobre la tensión activa que desarrolla una fibra muscular en contracción. A la derecha se muestran distintos grados de superposición entre los filamentos de miosina y actina a diversas longitudes del sarcómero. En el punto D del diagrama el filamento de actina ha producido una tracción de toda la longitud hasta el final del filamento de miosina, sin superposición entre la actina y la miosina. En este punto la tensión que desarrolla el músculo activado es cero. Después, a medida que el sarcómero se acorta y que el filamento de actina comienza a superponerse al filamento de miosina, la tensión aumenta progresivamente hasta que la longitud del sarcómero disminuye a aproximadamente 2,2 μm. En este punto el filamento de actina ya se ha superpuesto a todos los puentes cruzados del filamento de miosina, aunque todavía no ha alcanzado el centro del filamento de miosina. Con un acortamiento adicional el sarcómero mantiene la tensión completa hasta que se llega al punto B, a una longitud del sarcómero de aproximadamente 2 μm. En este punto los extremos de los dos filamentos de actina comienzan a superponerse entre sí además de superponerse a los filamentos de miosina. A medida que la longitud del sarcómero disminuye desde 2 μm hasta aproximadamente 1,65 μm, en el punto A, se produce una rápida disminución de la fuerza de la contracción. En este punto los dos discos Z del sarcómero se encuentran apoyados en los extremos de los filamentos de miosina. Después, a medida que se produce la contracción hasta longitudes del sarcómero aún menores, los extremos de los filamentos de miosina están corrugados y, como se muestra en la figura, la fuerza de la contracción se aproxima a cero, aunque el sarcómero ya se ha contraído hasta su mínima longitud.
FIGURA 6-9 Diagrama longitud-tensión de un sarcómero único contraído totalmente, que muestra la máxima fuerza de contracción cuando el sarcómero mide de 2 a 2,2 μm de longitud. En la parte superior derecha están las posiciones relativas de los filamentos de actina y miosina a diferentes longitudes del sarcómero desde el punto A al punto D. (Modificado de Gordon AM, Huxley AF, Julian FJ: The length-tension diagram of single vertebrate striated muscle fibers. J Physiol 171:28P, 1964.)
Efecto de la longitud muscular sobre la fuerza de contracción en el músculo intacto entero
La curva superior de la figura 6-10 es similar a la de la figura 6-9, pero la curva de la figura 6- representa la tensión del músculo entero intacto y no la de una única fibra muscular. El músculo entero tiene una gran cantidad de tejido conjuntivo; además, los sarcómeros de diferentes partes del músculo no siempre se contraen la misma magnitud. Por tanto, la curva tiene unas dimensiones algo diferentes de las que se muestran para la fibra muscular individual, aunque muestra la misma forma general para la pendiente en el intervalo normal de contracción, como se señala en la figura 6-.
FIGURA 6-11 Relación entre la carga y la velocidad de contracción de un músculo esquelético que tiene una sección transversal de 1 cm^2 y una longitud de 8 cm.
La disminución de la velocidad de contracción al aumentar la carga está producida por el hecho de que una carga sobre un músculo en contracción es una fuerza inversa que se opone a la fuerza contráctil que produce la contracción muscular. Por tanto, la fuerza neta de que se dispone para producir la velocidad de acortamiento está reducida de manera proporcional.
Energética de la contracción muscular
Generación de trabajo durante la contracción muscular
Cuando un músculo se contrae contra una carga realiza un trabajo. Esto significa que se transfiere energía desde el músculo hasta la carga externa para levantar un objeto hasta una mayor altura o para superar la resistencia al movimiento. En términos matemáticos el trabajo se define mediante la siguiente ecuación:
donde T es el trabajo generado, C es la carga y D es la distancia del movimiento que se opone a la carga. La energía necesaria para realizar el trabajo procede de las reacciones químicas de las células musculares durante la contracción, como se describe en las secciones siguientes.
Tres fuentes de energía para la contracción muscular
La mayor parte de la energía necesaria para la contracción muscular se utiliza para activar el mecanismo de cremallera mediante el cual los puentes cruzados tiran de los filamentos de actina, aunque son necesarias cantidades pequeñas para: 1) bombear iones calcio desde el sarcoplasma hacia el interior del retículo sarcoplásmico después de que haya finalizado la contracción, y 2) para bombear iones sodio y potasio a través de la membrana de la fibra muscular para mantener un entorno iónico adecuado para la propagación de los potenciales de acción de la fibra muscular. La contracción de ATP en la fibra muscular, de aproximadamente 4 milimolar, es suficiente para mantener la contracción completa durante solo 1 a 2 s como máximo. El ATP se escinde para formar ADP, que transfiere la energía de la molécula de ATP a la maquinaria contráctil de la fibra muscular. Después, como se describe en el capítulo 2, el ADP se vuelve a fosforilar para formar nuevo ATP en otra fracción de segundo, lo que permite que el músculo mantenga su contracción. Hay varias fuentes de energía para esta nueva fosforilación. La primera fuente de energía que se utiliza para reconstituir el ATP es la sustancia fosfocreatina, que contiene un enlace fosfato de alta energía similar a los enlaces del ATP. El enlace fosfato de alta energía de la fosfocreatina tiene una cantidad ligeramente mayor de energía libre que la de cada uno de los enlaces del ATP, como se analiza con más detalle en los capítulos 68 y 73. Por tanto, la fosfocreatina se escinde inmediatamente y la energía que se libera produce el enlace de un nuevo ion fosfato al ADP para reconstituir el ATP. Sin embargo, la cantidad total de fosfocreatina en la fibra muscular también es pequeña, solo aproximadamente cinco veces mayor que la de ATP. Por tanto, la energía combinada del ATP y de la fosfocreatina almacenados en el músculo es capaz de producir una contracción muscular máxima durante solo 5 a 8 s. La segunda fuente importante de energía, que se utiliza para reconstituir tanto el ATP como la fosfocreatina, es la «glucólisis» del glucógeno que se ha almacenado previamente en las células musculares. La escisión enzimática rápida del glucógeno en ácido pirúvico y ácido láctico libera energía que se utiliza para convertir el ADP en ATP; después se puede utilizar directamente el ATP para aportar energía a la contracción muscular adicional y también para reconstituir los almacenes de