¡Descarga Metabolismo del Glucógeno: Glucogénesis, Glucogenólisis y Regulación Hormonal y más Transcripciones en PDF de Química solo en Docsity!
Introducción
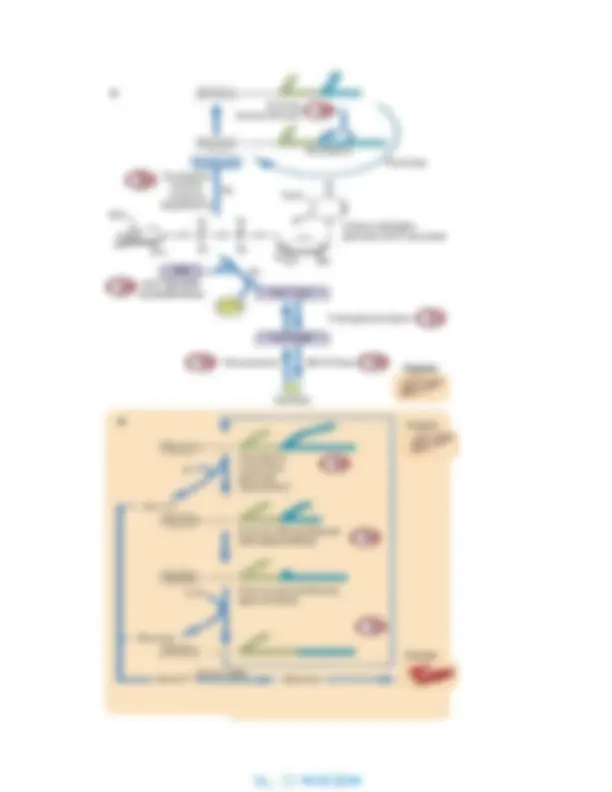
Los eritrocitos y el cerebro tienen una necesidad absoluta de glucosa sanguínea para el metabolismo energético. Estas células consumen aproximadamente el 80% de los 200 g de glucosa que consume el organismo cada día. En el plasma y en el volumen de líquido extracelular solo hay unos 10 g de glucosa, lo que equivale aproximadamente al 5% de las necesidades diarias, de manera que el contenido de glucosa sanguínea debe rellenarse constantemente. De lo contrario, aparece hipoglucemia, que compromete a la función cerebral dando lugar a confusión, desorientación y, posiblemente, a un estado de coma de riesgo vital a concentraciones de glucosa por debajo de 2,5 mmol/l (45 mg/dl). Absorbemos glucosa de nuestro intestino solo durante las 2-3 h siguientes a una comida que contenga hidratos de carbono, así que debe haber un mecanismo para mantener la glucosa en sangre entre las comidas. El glucógeno, un polisacárido de almacenamiento de la glucosa, es nuestra primera línea de defensa contra la disminución de la concentración de glucosa en sangre. Durante e inmediatamente después de una comida, la glucosa se convierte en glucógeno tanto en el hígado como en el músculo, un proceso conocido como glucogénesis. La concentración tisular de glucógeno es mayor en el hígado que en el músculo, pero debido a las masas relativas del músculo y del hígado, la mayoría del glucógeno del organismo se almacena en el músculo (tabla 12.1).
Tabla 12.
Distribución tisular de las reservas de energía de
hidratos de carbono (adulto de 70 kg)
La glucogenólisis y la gluconeogénesis hepática son
necesarias para el mantenimiento de la concentración
normal de glucosa en sangre
El glucógeno hepático se degrada gradualmente entre las comidas por la vía de la glucogenólisis , liberando glucosa para mantener su concentración en sangre. Sin embargo, las reservas totales de glucógeno en el hígado apenas son suficientes para mantener una concentración de glucosa en sangre en un ayuno de 12 h. Durante el sueño, cuando no estamos comiendo, hay un desplazamiento gradual de la glucogenólisis a una síntesis de novo de la glucosa, también a través de una vía hepática conocida como gluconeogénesis (fig. 12.1). La gluconeogénesis es esencial para la supervivencia en el ayuno o la inanición, cuando las reservas de glucógeno son mínimas. El hígado utiliza los aminoácidos de las proteínas musculares como precursores primarios de la glucosa, pero también utiliza lactato de la glucólisis y el glicerol del catabolismo de la grasa. Los ácidos grasos, movilizados desde las reservas de triglicéridos del tejido adiposo, proporcionan la energía para la gluconeogénesis.
Estructura del glucógeno
El glucógeno, un glucano sumamente ramificado,
constituye la forma de almacenamiento de la glucosa en
los tejidos
El glucógeno es un polisacárido ramificado de glucosa. Contiene solo dos tipos de enlaces glucosídicos, cadenas de residuos de glucosa con enlaces α1→4 con ramificaciones α1→6 espaciadas cada 4-6 residuos, aproximadamente, a lo largo de la cadena α1→4 (fig. 12.2). El glucógeno está estrechamente relacionado con el almidón, el polisacárido de almacenamiento de las plantas, pero el almidón consta de una mezcla de amilosa y amilopectina. El componente amilosa contiene solo cadenas α1→4 lineales; el componente amilopectina es una estructura más parecida al glucógeno, pero con menos ramificaciones α1→6, aproximadamente 1 por cada 12 residuos de glucosa con enlaces α1→4. La estructura macroscópica del glucógeno es de naturaleza dendrítica, expandiéndose desde una secuencia nuclear ligada a un residuo de tirosina en la proteína glucogenina y desarrollándose hasta una estructura final que se asemeja a una coliflor. Las enzimas del metabolismo del glucógeno están ligadas a la superficie de la partícula de glucógeno; numerosas moléculas terminales de glucosa en la superficie de la molécula proporcionan un acceso fácil para la liberación rápida de la glucosa a partir del polímero de glucógeno.
FIG. 12.2 Esquema de la estructura del glucógeno. La figura muestra las cadenas α1→4 y un punto de ramificación α1→6. El glucógeno se almacena como gránulos en el citoplasma del hígado y del músculo.
tejido adiposo como fuente de glicerol para la biosíntesis de triglicéridos. La vía de la glucogénesis a partir de la glucosa (fig. 12.3A) consta de cuatro pasos:
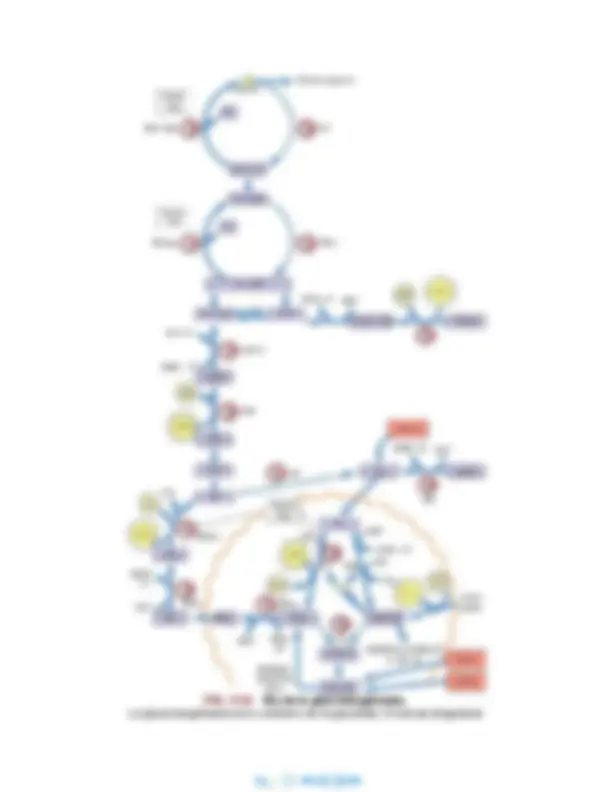
▪ Conversión de Glc-6-P en glucosa-1-fosfato (Glc-1-P) por la fosfoglucomutasa. ▪ Activación de la Glc-1-P para formar el azúcar nucleotídico, uridina difosfato (UDP)-glucosa mediante la enzima UDP- glucosa pirofosforilasa. ▪ Transferencia de glucosa desde la UDP-Glc al glucógeno en un enlace α1→4 mediante la glucógeno sintasa, un miembro de la clase de enzimas conocida como glucosil transferasas. ▪ Cuando la longitud de la cadena α1→4 es mayor de 8 residuos, la enzima ramificante del glucógeno , una transglucosilasa, transfiere parte de los azúcares de los enlaces α1→4 a una rama α1→6, estableciendo el estadio para el alargamiento continuo de ambas cadenas en α1→4 hasta que, a su vez, se hacen lo suficientemente largas para la transferencia por la enzima ramificante.
Vía de la glucogenólisis hepática
La glucógeno fosforilasa hepática permite una liberación
rápida de glucosa a la sangre durante la fase posterior a
la absorción
Lo mismo que ocurre con la mayoría de las vías metabólicas, se requieren enzimas distintas, a veces en compartimentos subcelulares separados, para las vías directas e inversas. La vía de la glucogenólisis (v. fig. 12.3B) comienza con la eliminación de los abundantes residuos externos de glucosa unidos mediante enlaces α1→4 en el glucógeno. Esto se realiza, no mediante una hidrolasa, sino por la glucógeno fosforilasa, una enzima que utiliza el fosfato citosólico y libera glucosa del glucógeno en forma de Glc-1-P. La Glc-1-P se isomeriza por la fosfoglucomutasa a Glc-6-P, situándola al inicio de la vía glucolítica; la reacción de la fosforilasa tiene como efecto evitar la necesidad de ATP en las reacciones de la hexocinasa o de la glucocinasa. En el hígado, la glucosa se libera a partir de la Glc-6-P por la glucosa-6-fosfatasa (Glc-6-Pasa) y la glucosa se incorpora a la sangre mediante el transportador GLUT-2. El paso limitante y regulador en la glucogenólisis está catalizado por la fosforilasa, la primera enzima de la vía. La fosforilasa es específica para los enlaces glucosídicos α1→4; no puede escindir los enlaces α1→6. Además, esta gran enzima no puede aproximarse eficazmente a los residuos ramificados de glucosa. Así, como se muestra en la figura 12.3B, la fosforilasa escinde los residuos externos de glucosa hasta que las ramas tienen una longitud de 3 o 4 residuos, y a continuación, la enzima desramificante, que tiene actividad transglucosilasa y también glucosidasa, mueve un segmento corto de residuos de glucosa unidos a la rama α1→6 al extremo de una cadena adyacente α1→4, dejando un único residuo de glucosa en el punto de ramificación. A continuación, esta glucosa es eliminada por la actividad exo-1,6-glucosidasa de la enzima desramificante permitiendo a la glucógeno fosforilasa proceder con la degradación de
la cadena α1→4 extendida hasta que se aproxima a otro punto de ramificación, estableciendo el estadio de repetición de las reacciones transglucosilasa y glucosidasa. Aproximadamente el 90% de la glucosa se libera del glucógeno como Glc-1-P y, el resto, derivado de los residuos ramificados α1→6, se libera como glucosa libre.
Conceptos clínicos
Enfermedad de von Gierke: glucogenosis por déficit
de Glc-6-Pasa
Un bebé de sexo femenino se encontraba siempre de mal humor, irritable, sudorosa y letárgica, y pedía alimento con frecuencia. La exploración física indicaba abdomen voluminoso secundario a hepatomegalia. La glucosa sanguínea, medida 1 hora después de la alimentación, era de 3,5 mmol/l (70 mg/dl); valor normal <5 mmol/l (100 mg/dl). Después de 4 h, cuando la niña estaba irritable y sudorosa, la frecuencia cardíaca había aumentado (pulso = 110) y la glucosa sanguínea había disminuido a 2 mmol/l (40 mg/dl). Estos síntomas se corrigieron al alimentarla. Una biopsia hepática mostraba un depósito masivo de partículas de glucógeno en el citosol hepático.
Comentario
Esta niña no puede movilizar el glucógeno. Dada la gravedad de la hipoglucemia, lo más probable es que la mutación sea en la Glc-6- Pasa hepática, necesaria para la producción de glucosa mediante la glucogenólisis y la gluconeogénesis. El tratamiento consiste en alimentación frecuente con hidratos de carbono de digestión lenta (p. ej., almidón sin cocer) y alimentación por goteo nasogástrico durante la noche.
baja en hidratos de carbono (v. cap. 31). La glucogenólisis hepática también se activa como respuesta al estrés agudo y crónico. El estrés puede ser de varios tipos:
▪ Fisiológico (p. ej., como respuesta al aumento de la utilización de la glucosa en sangre durante el ejercicio físico). ▪ Patológico (p. ej., como resultado de la pérdida de sangre [shock]). ▪ Psicológico (p. ej., como respuesta a amenazas agudas o crónicas).
Independientemente de su origen, el estrés agudo desencadena una activación de la glucogenólisis a través de la acción de la adrenalina, una hormona catecolamínica, liberada desde la médula suprarrenal. Durante el ejercicio prolongado o estresante, tanto el glucagón como la adrenalina contribuyen a la estimulación de la glucogenólisis y al mantenimiento de la concentración de glucosa sanguínea. El aumento de las concentraciones sanguíneas de cortisol, una hormona esteroidea corticosuprarrenal, también induce glucogenólisis. Las concentraciones del glucocorticoide cortisol varían en el plasma a lo largo del día, pero pueden estar crónicamente elevadas en situaciones continuas de estrés, incluido el estrés psicológico y ambiental (p. ej., frío). El glucagón sirve como modelo general para el mecanismo de acción de las hormonas que actúan mediante receptores de superficie celular. El cortisol, que actúa a nivel de la expresión génica, se comenta en los capítulos 23 y 25.
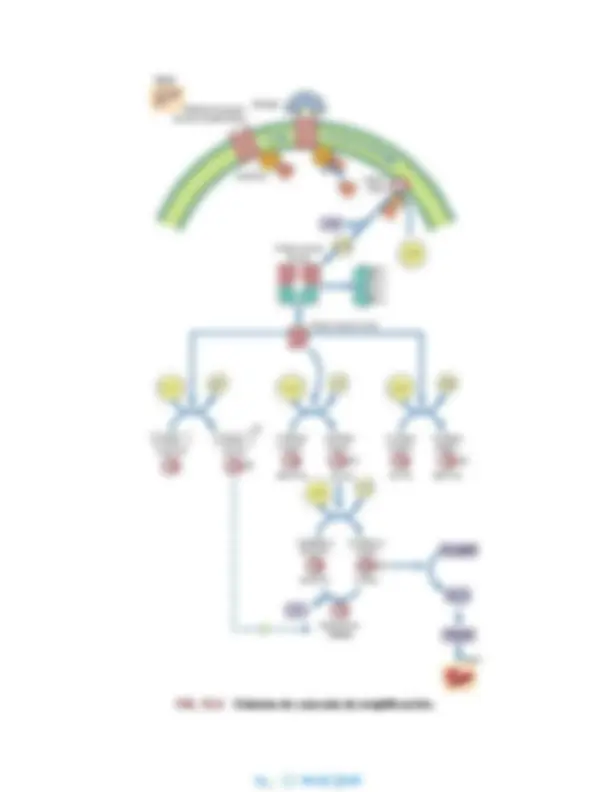
Mecanismo de acción del glucagón
El glucagón activa la glucogenólisis durante el período
posterior a la absorción
El glucagón se une a un receptor de la membrana plasmática del hepatocito e inicia una cascada de reacciones que conducen a la movilización del glucógeno hepático (fig. 12.4) durante el estado posterior a la absorción. En el lado interno de la membrana plasmática hay una clase de proteínas de transducción de señal conocidas como proteínas G que fijan guanosina trifosfato (GTP) y guanosina difosfato (GDP), nucleótidos análogos a ATP y ADP. El GDP está unido en situación de reposo. La unión del glucagón al receptor de la membrana plasmática estimula el intercambio de GDP por GTP en la proteína G y, a continuación, la proteína G sufre un cambio conformacional que da lugar a la disociación de su subunidad α, que después se une y activa la enzima de membrana plasmática, adenilato ciclasa. Esta enzima convierte el ATP citoplasmático en 3’,5’-AMP cíclico (AMPc), un mensajero soluble que se describe como el «segundo mensajero» de la acción del glucagón (y de otras hormonas). El AMP cíclico se une a la enzima citoplasmática proteína cinasa A (PKA) provocando la disociación de las subunidades inhibidoras (reguladoras) de las subunidades catalíticas de la enzima heterodimérica, liberando la inhibición de la PKA, que a continuación fosforila residuos de serina y treonina en proteínas diana y enzimas.
Movilización del glucógeno hepático por el glucagón. Una cascada de reacciones amplifica la respuesta hepática a la fijación del glucagón a su receptor de membrana plasmática. El AMPc se conoce como el segundo mensajero de la acción del glucagón. La proteína cinasa A activa indirectamente la fosforilasa a través de la fosforilasa cinasa e inactiva directamente la glucógeno sintasa. C, subunidades catalíticas; R, subunidades reguladoras (inhibidoras).
Conceptos clínicos
Enfermedad de McArdle: una glucogenosis que reduce la
capacidad para el ejercicio
Un hombre de 30 años consultó a su médico por mialgias crónicas en brazos y piernas, y calambres durante el ejercicio. El paciente refería que siempre había tenido debilidad muscular y, por esta razón, nunca participó en deportes en la escuela, pero el problema no se agravó hasta que recientemente se inscribió en un programa de ejercicio para mejorar su salud. Asimismo, notó que el dolor generalmente desaparecía tras 15-30 minutos, y a continuación podía continuar su ejercicio sin molestias. Su concentración de glucosa sanguínea era normal durante el ejercicio, pero la creatina cinasa sérica (isoforma MM del músculo esquelético) estaba elevada, sugiriendo una lesión muscular. La glucosa en sangre disminuyó ligeramente durante 15 minutos de ejercicio, pero de forma inesperada también disminuyó el lactato sanguíneo, en vez de aumentar, incluso cuando estaba teniendo calambres musculares. Una biopsia indicó un nivel inusualmente elevado de glucógeno en el músculo, sugiriendo una glucogenosis (enfermedad por almacenamiento de glucógeno).
Comentario
Este paciente tiene la enfermedad de McArdle, una deficiencia infrecuente de la actividad fosforilasa en el músculo. La deficiencia enzimática real debe confirmarse mediante un análisis enzimático, ya que otras mutaciones también pueden afectar al metabolismo del glucógeno en el músculo. En los períodos iniciales de ejercicio intenso, el músculo obtiene la mayor parte de la energía mediante el
metabolismo de la glucosa, procedente del glucógeno. Durante los calambres, que normalmente tienen lugar durante la deficiencia de oxígeno, gran parte del piruvato producido por la glucólisis se excreta a la sangre como lactato, lo que lleva a un aumento de la concentración de lactato sanguíneo. Sin embargo, en este caso el paciente padecía calambres y no excretaba lactato, lo que sugiere insuficiencia para movilizar el glucógeno muscular para producir glucosa. Su recuperación tras 15-30 minutos es resultado de la activación de la glucogenólisis hepática mediada por la adrenalina, que suministra glucosa a la sangre, superando la deficiencia de glucogenólisis muscular. El tratamiento de la enfermedad de McArdle habitualmente supone evitar el ejercicio o, cuando es necesario, consumir hidratos de carbono antes del mismo. Por otra parte, la enfermedad cursa sin incidentes.
Conceptos avanzados
Proteínas G
Las proteínas G son proteínas de la membrana plasmática que fijan nucleótidos de guanosina y que están implicadas en la transducción de señales de una amplia variedad de hormonas (fig. 12.4; v. también cap. 25). En algunos casos estimulan (Gs); en otros casos, inhiben (Gi) las proteína cinasas y la fosforilación de proteínas. Las proteínas G están estrechamente asociadas con los receptores hormonales en las membranas plasmáticas y constan de subunidades α, β y γ. La subunidad Gα fija GDP en reposo. Tras la unión con la hormona
(fijación), el receptor recluta las proteínas G, estimulando el intercambio de GDP por GTP en la subunidad Gα. La unión de GTP
da lugar a la liberación de las subunidades β y γ. A continuación, la subunidad α queda libre para unirse a la adenilato ciclasa y activarla. La respuesta hormonal se amplifica tras la unión con el receptor, porque un único receptor puede activar numerosas subunidades α. La respuesta hormonal también se extingue en los receptores y las proteínas G mediante dos mecanismos:
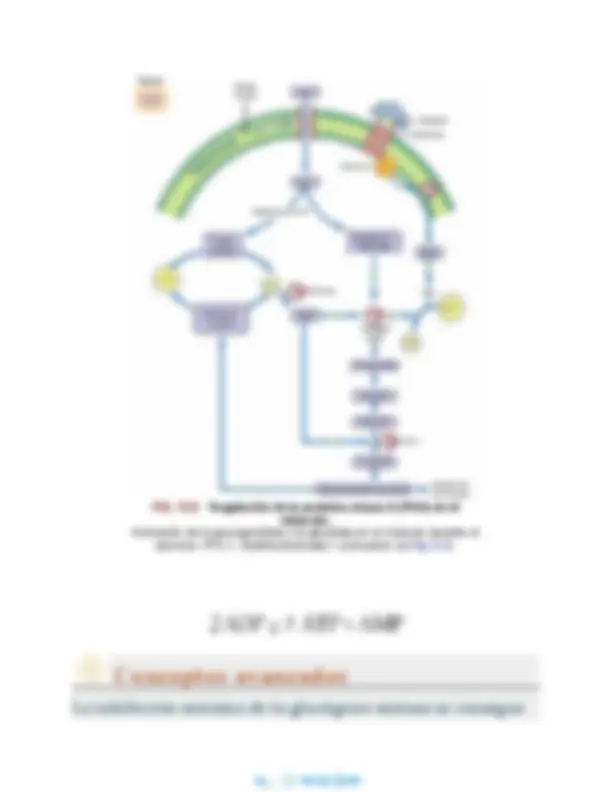
cinasa por la PKA, que después fosforila y activa muchas moléculas de la glucógeno fosforilasa. El efecto neto de estos pasos secuenciales, que se inician con la activación de muchas moléculas de adenilato ciclasa por proteínas G, es un sistema de «cascada de amplificación», similar a una serie de amplificadores de un equipo de radio o de estéreo, dando lugar a un aumento enorme en la potencia de la señal segundos después de la unión del glucagón a la membrana plasmática del hepatocito. La fosforilasa b (la forma inactiva y no fosforilada de la fosforilasa) normalmente es inhibida por el ATP y la glucosa en el hígado, pero al fosforilarse se convierte en su forma activa, la fosforilasa a (fig. 12.4), que activa la glucogenólisis y produce Glc y Glc-1-P, que son convertidas en Glc-6-P e hidrolizadas a glucosa que se exporta a la sangre.
La glucogenólisis y la glucogénesis están
contrarreguladas por la proteína cinasa A, que activa la
fosforilasa e inhibe la glucógeno sintasa
La glucogenólisis y la glucogénesis son vías opuestas. Teóricamente, la Glc-1-P producida por la fosforilasa podría activarse rápidamente a UDP-glucosa y reincorporarse al glucógeno. Para evitar este ciclo desaprovechado o fútil, la PKA también fosforila la glucógeno sintasa, inactivando la enzima en este caso. Así, la PKA activa la fosforilasa (glucogenólisis) e inactiva la glucógeno sintasa (glucogénesis) de una manera coordinada. Otras vías biosintéticas hepáticas (como la síntesis de proteínas, colesterol, ácidos grasos y triglicéridos, así como la glucólisis) también están reguladas por la fosforilación de enzimas reguladoras cruciales, limitando por lo general las reacciones de biosíntesis y dirigiendo el metabolismo del hígado en respuesta al glucagón hacia el suministro de glucosa a la sangre para mantener las funciones corporales vitales (v. cap. 31). Para equilibrar la cascada de fenómenos que amplifican la respuesta glucogenolítica al glucagón, existen varios mecanismos redundantes para asegurar un final rápido de la respuesta hormonal (tabla 12.3). Además de la lenta actividad GTPasa de la subunidad Gα, también
existe una fosfodiesterasa en la célula que hidroliza el AMPc a AMP,
permitiendo la reasociación de las subunidades inhibidoras y las catalíticas de la PKA, disminuyendo su actividad proteína cinasa. También hay fosfatasas de fosfoproteínas que eliminan los grupos fosfato de las formas activas fosforiladas de la fosforilasa cinasa y de la fosforilasa. Otro objetivo de la PKA es el inhibidor-1, una proteína inhibidora de la fosfatasa de fosfoproteínas, que se activa mediante fosforilación. El inhibidor-1 fosforilado inhibe las fosfatasas de fosfoproteínas citoplásmicas, que de lo contrario revertirían la fosforilación de las enzimas y desactivarían la respuesta al glucagón (v. fig. 12.4). La disminución de la concentración de AMPc y de la actividad de la PKA también da lugar a una disminución de la fosforilación del inhibidor-1, permitiendo un aumento de actividad de las fosfatasas de fosfoproteínas. Así, numerosos mecanismos actúan concertadamente para asegurar que la glucogenólisis hepática disminuya con rapidez como respuesta al aumento de glucosa sanguínea y a la disminución de la concentración sanguínea de glucagón después de una comida.
Tabla 12.
Varios mecanismos están implicados en la finalización de la respuesta hormonal al glucagón
- Hidrólisis del GTP en la subunidad Gα
- Hidrólisis del AMPc por la fosfodiesterasa
- Actividad proteína fosfatasa
Existen varias enfermedades genéticas autosómicas recesivas que afectan al metabolismo del glucógeno (tabla 12.4). Estas enfermedades, conocidas como glucogenosis (enfermedades que afectan al almacenamiento del glucógeno), se caracterizan por la acumulación de gránulos de glucógeno en los tejidos que, finalmente, afectan a la función tisular. De forma predecible, las glucogenosis que afectan al metabolismo hepático se caracterizan por hipoglucemia en ayunas y pueden poner en peligro la vida, mientras que los defectos en el metabolismo del glucógeno muscular se caracterizan por una fatiga muscular rápida durante el ejercicio.