¡Descarga capitulo 4 guyton y hall y más Apuntes en PDF de Fisiología solo en Docsity!
CAPITULOP 4
TRANSPORTE DE SUSTANCIAS A TRAVÉS DE LAS MEMBRANAS CELULARES
La membrana celular consiste en una BICAPA lipídica con proteínas de transporte de la membrana celular- La bicapa lipídica no es miscible con el líquido extracelular ni con el líquido intracelular. Por tanto, constituye una barrera frente al movimiento de moléculas de agua y de sustancias insolubles entre los compartimientos del líquido extracelular e intracelular Las moléculas proteicas de la membrana tienen unas propiedades totalmente diferentes para transportar sustancias. Sus estructuras moleculares interrumpen la continuidad de la bicapa lipídica y constituyen una ruta alternativa a través de la membrana celular. Muchas de estas proteínas penetrantes pueden actuar como proteínas transportadoras. Proteínas diferentes actúan de una manera diferente. Algunas tienen espacios acuosos en todo el trayecto del interior de la molécula y permiten el movimiento libre de agua, así como de iones o moléculas seleccionados; estas proteínas se denominan proteínas de los canales. Otras, denominadas proteínas transportadoras, se unen a las moléculas o iones que se van a transportar y cambios conformacionales de las moléculas de la proteína desplazan después las sustancias a través de los intersticios de la proteína hasta el otro lado de la membrana. Las proteínas de los canales y las proteínas transportadoras habitualmente son selectivas para los tipos de moléculas o de iones que pueden atravesar la membrana. «Difusión» frente a «transporte activo» El transporte a través de la membrana celular, ya sea directamente a través de la bicapa lipídica o a través de las proteínas, se produce mediante uno de dos procesos básicos: difusión o transporte activo. Aunque hay muchas variaciones de estos mecanismos básicos, la difusión se refiere a un movimiento molecular aleatorio de las sustancias molécula a molécula, a través de espacios intermoleculares de la membrana o en combinación con una proteína transportadora. La energía que hace que se produzca la difusión es la energía del movimiento cinético normal de la materia. Por el contrario, el transporte activo se refiere al movimiento de iones o de otras sustancias a través de la membrana en combinación con una proteína transportadora de tal manera que la proteína transportadora hace que la sustancia se mueva contra un gradiente de energía, como desde un estado de baja concentración a un estado de alta concentración. Este movimiento precisa una fuente de energía adicional, además de la energía cinética.
Difusión: movimiento continuo de moléculas entre sí en los líquidos o los gases se denomina difusión Difusión a través de la membrana celular: La difusión a través de la membrana celular se divide en dos subtipos, denominados difusión simple y difusión facilitada. Difusión simple significa que el movimiento cinético de las moléculas o de los iones se produce a través de una abertura de la membrana o a través de espacios intermoleculares sin ninguna interacción con las proteínas transportadoras de la membrana. La velocidad de difusión viene determinada por la cantidad de sustancia disponible, la velocidad del movimiento cinético y el número y el tamaño de las aberturas de la membrana a través de las cuales se pueden mover las moléculas o los iones. La difusión facilitada precisa la interacción de una proteína transportadora. La proteína transportadora ayuda al paso de las moléculas o de los iones a través de la membrana mediante su unión química con estos y su desplazamiento a través de la membrana de esta manera. Se puede producir difusión simple a través de la membrana celular por dos rutas: 1) a través de los intersticios de la bicapa lipídica si la sustancia que difunde es liposoluble, y 2) a través de canales acuosos que penetran en todo el grosor de la bicapa a través de las grandes proteínas transportadoras Difusión a través de poros y canales proteicos: permeabilidad selectiva y «activación» de canales Las reconstrucciones tridimensionales computarizadas de los poros y canales proteicos han mostrado trayectos tubulares que se extienden desde el líquido extracelular hasta el intracelular. Por tanto, las sustancias se pueden mover mediante difusión simple directamente a lo largo de estos poros y canales desde un lado de la membrana hasta el otro. Los poros están compuestos por proteínas de membranas celulares integrales que forman tubos abiertos a través de la membrana y que están siempre abiertos. Sin embargo, el diámetro de un poro y sus cargas eléctricas proporcionan una selectividad que permite el paso de solo ciertas moléculas a su través. Por ejemplo, los poros proteicos denominados acuaporinas o canales de agua permiten el rápido paso de agua a través de las membranas celulares pero impiden el de otras moléculas. En las distintas células del cuerpo humano se han descubierto al menos 13 tipos diferentes de acuaporinas. Las acuaporinas tienen un poro estrecho que permite que las moléculas de agua se difundan a través de la membrana en una única fila. El poro es demasiado pequeño para permitir el paso de iones hidratados Los canales proteicos se distinguen por dos características importantes: 1) con frecuencia son permeables de manera selectiva a ciertas sustancias, y 2) muchos de los canales se pueden abrir o cerrar por compuertas que son reguladas por señales eléctricas (canales activados por el voltaje) o sustancias químicas que se unen a las proteínas de canales (canales activados por ligandos). Activación de los canales proteicos
- Activación por el voltaje. En el caso de activación por el voltaje, la conformación molecular de la compuerta o de sus enlaces químicos responde al potencial eléctrico que se establece a través de la membrana celular
- Activación química (por ligando). Las compuertas de algunos canales proteicos se abren por la unión de una sustancia química (un ligando) a la proteína, que produce un cambio conformacional o un cambio de los enlaces químicos de la molécula de la proteína que abre o cierra la compuerta. Uno de los casos más importantes de activación química es el efecto de la acetilcolina sobre el denominado canal de la acetilcolina. La acetilcolina abre la compuerta de este canal, dando lugar a la apertura de un poro de carga negativa de aproximadamente 0,65 nm de diámetro que permite que lo atraviesen moléculas sin carga o iones positivos menores de este diámetro. Esta compuerta es muy importante para la transmisión de las señales nerviosas desde una célula nerviosa a otra
Potencial de membrana en reposo de las neuronas El potencial de membrana en reposo de las fibras nerviosas grandes cuando no transmiten señales nerviosas es de aproximadamente –90 mV. Es decir, el potencial en el interior de la fibra es 90 mV más negativo que el potencial del líquido extracelular que está en el exterior de la misma. En los siguientes párrafos se explican las propiedades de transporte de la membrana en reposo de los nervios para el sodio y el potasio, así como los factores que determinan el nivel de este potencial en reposo. Transporte activo de los iones sodio y potasio a través de la membrana: la bomba sodio-potasio (Na+-K+) Como se señaló en el capítulo 4, todas las membranas celulares del cuerpo tienen una potente bomba Na+-K+ que transporta continuamente iones sodio hacia el exterior de la célula e iones potasio hacia el interior, como se señala en el lado izquierdo de la figura 5-4. Debe observarse que se trata de una bomba electrógena porque se bombean más cargas positivas hacia el exterior que hacia el interior (tres iones Na+ hacia el exterior por cada dos iones K+ hacia el interior), dejando un déficit neto de iones positivos en el interior; esto genera un potencial negativo en el interior de la membrana celular Potencial de acción de las neuronas Las señales nerviosas se transmiten mediante potenciales de acción que son cambios rápidos del potencial de membrana que se extienden rápidamente a lo largo de la membrana de la fibra nerviosa. Cada potencial de acción comienza con un cambio súbito desde el potencial de membrana negativo en reposo normal hasta un potencial positivo y termina con un cambio casi igual de rápido de nuevo hacia el potencial negativo. Para conducir una señal nerviosa el potencial de acción se desplaza a lo largo de la fibra nerviosa hasta que llega a su extremo. Las sucesivas fases del potencial de acción son las siguientes. Fase de reposo La fase de reposo es el potencial de membrana en reposo antes del comienzo del potencial de acción. Se dice que la membrana está «polarizada» durante esta fase debido al potencial de membrana negativo de –90 mV que está presente. Fase de despolarización En este momento la membrana se hace súbitamente muy permeable a los iones sodio, lo que permite que un gran número de iones sodio con carga positiva difunda hacia el interior del axón. El estado «polarizado» normal de –90 mV se neutraliza inmediatamente por la entrada de iones sodio cargados positivamente, y el potencial aumenta rápidamente en dirección positiva, un proceso denominado despolarización. En las fibras nerviosas grandes el gran exceso de iones sodio positivos que se mueven hacia el interior hace que el potencial de membrana realmente se «sobreexcite» más allá del nivel cero y que se haga algo positivo. En algunas fibras más pequeñas, así como en muchas neuronas del sistema nervioso central, el potencial simplemente se acerca al nivel cero y no hay sobreexcitación hacia el estado positivo. Fase de repolarización En un plazo de algunas diezmilésimas de segundo después de que la membrana se haya hecho muy permeable a los iones sodio, los canales de sodio comienzan a cerrarse y los canales de potasio se abren más de lo normal. De esta manera, la rápida difusión de los iones potasio hacia el exterior restablece el potencial de membrana en reposo negativo normal, que se denomina repolarización de la membrana. Para explicar más en detalle los factores que producen tanto la despolarización como la repolarización se describirán las características especiales de otros dos tipos de canales transportadores que atraviesan la membrana nerviosa: los canales de sodio y de potasio activados por el voltaje. Canales de sodio y potasio activados por el voltaje El actor necesario en la producción tanto de la despolarización como de la repolarización de la membrana nerviosa durante el potencial de acción es el canal de sodio activado por el voltaje. Un canal de potasio activado por el voltaje también tiene una función importante en el aumento de la rapidez de la
repolarización de la membrana. Estos dos canales activados por el voltaje tienen una función adicional a la de la bomba Na+-K+ y de los canales de fuga de K+. Activación del canal de sodio Cuando el potencial de membrana se hace menos negativo que durante el estado de reposo, aumentando desde –90 mV hacia cero, finalmente alcanza un voltaje (habitualmente algún punto entre –70 y – 50 mV) que produce un cambio conformacional súbito en la activación de la compuerta, que bascula totalmente hasta la posición de abierta. Durante este estado activado, los iones sodio pueden atravesar el canal, aumentando la permeabilidad de la membrana al sodio hasta 500 a 5.000 veces. Inactivación del canal de sodio La parte superior derecha de la figura 5-7 muestra un tercer estado del canal de sodio. El mismo aumento de voltaje que abre la compuerta de activación también cierra la compuerta de inactivación. Sin embargo, la compuerta de inactivación se cierra algunas diezmilésimas de segundo después de que se abra la compuerta de activación. Es decir, el cambio conformacional que hace bascular la compuerta de inactivación hacia el estado cerrado es un proceso algo más lento que el cambio conformacional que abre la compuerta de activación. Por tanto, después de que el canal de sodio haya permanecido abierto durante algunas diezmilésimas de segundo se cierra la compuerta de inactivación y los iones sodio ya no pueden pasar hacia el interior de la membrana. En este punto el potencial de membrana comienza a recuperarse de nuevo hacia el estado de membrana en reposo, lo que es el proceso de repolarización. Otra característica importante de la inactivación del canal de sodio es que la compuerta de inactivación no se abre de nuevo hasta que el potencial de membrana se normaliza o casi a valores de reposo. Por tanto, en general el canal de sodio no se puede abrir de nuevo sin que antes se repolarice la fibra nerviosa. Inicio del potencial de acción Hasta ahora hemos explicado la permeabilidad cambiante de la membrana al sodio y al potasio, así como la generación del potencial de acción, aunque no hemos explicado qué lo inicia. Un ciclo de retroalimentación positiva abre los canales de sodio Primero, siempre que no haya alteraciones de la membrana de la fibra nerviosa, no se produce ningún potencial de acción en el nervio normal. Sin embargo, si algún episodio produce una elevación suficiente del potencial de membrana desde –90 mV hacia el nivel cero, el propio aumento del voltaje hace que empiecen a abrirse muchos canales de sodio activados por el voltaje. Esta situación permite la entrada rápida de iones sodio, lo que produce una elevación adicional del potencial de membrana y abre aún más canales de sodio activados por el voltaje y permite que se produzca una mayor entrada de iones sodio hacia el interior de la fibra. Este proceso es un círculo vicioso de retroalimentación positiva que, una vez que la retroalimentación es lo suficientemente intensa, continúa hasta que se han activado (abierto) todos los canales de sodio activados por el voltaje. Posteriormente, en un plazo de otra fracción de milisegundo, el aumento del potencial de membrana produce cierre de los canales de sodio y apertura de los canales de potasio, y pronto finaliza el potencial de acción. Umbral para el inicio del potencial de acción No se producirá un potencial de acción hasta que el aumento inicial del potencial de membrana sea lo suficientemente grande como para dar origen al ciclo de retroalimentación positiva que se ha descrito en el párrafo anterior. Esto se produce cuando el número de iones sodio que entran en la fibra supera al número de iones potasio que salen. Habitualmente es necesario un aumento súbito del potencial de membrana de 15 a 30 mV. Por tanto, un aumento súbito del potencial de membrana en una fibra nerviosa grande desde –90 mV hasta aproximadamente –65 mV suele dar lugar a la aparición explosiva de un potencial de acción. Se dice que este nivel de –65 mV es el umbral para la estimulación.
Una vez que se ha originado un potencial de acción en cualquier punto de la membrana de una fibra normal, el proceso de despolarización viaja por toda la membrana si las condiciones son las adecuadas, o no viaja en absoluto si no lo son. Este principio se denomina principio del todo o nada y se aplica a todos los tejidos excitables normales. De manera ocasional, el potencial de acción alcanza un punto de la membrana en el que no genera un voltaje suficiente como para estimular la siguiente zona de la membrana. Cuando se produce esta situación se interrumpe la diseminación de la despolarización. Por tanto, para que se produzca la propagación continuada de un impulso, el cociente del potencial de acción respecto al umbral de excitación debe ser mayor de 1 en todo momento. Este requisito de «mayor de 1» se denomina factor de seguridad para la propagación. CAPÍTULO 6 Contracción del músculo esquelético Aproximadamente el 40% del cuerpo es músculo esquelético, y tal vez otro 10% es músculo liso y cardíaco. Algunos de los principios básicos de la contracción se aplican también a los diferentes tipos de músculos. En este capítulo se considera principalmente la función del músculo esquelético; las funciones especializadas del músculo liso se analizan en el capítulo 8 y el músculo cardíaco se analiza en el capítulo 9. Anatomía fisiológica del músculo esquelético Fibras del músculo esquelético La figura 6-1 ilustra la organización del músculo esquelético y muestra que todos los músculos esqueléticos están formados por numerosas fibras cuyo diámetro varía entre 10 y 80 μm. Cada una de estas fibras está formada por m. Cada una de estas fibras está formada por subunidades cada vez más pequeñas, que también se muestran en la figura 6-1 y que se describen en los párrafos siguientes En la mayoría de los músculos esqueléticos, las fibras se extienden a lo largo de toda la longitud del músculo. Todas las fibras, excepto alrededor de un 2%, habitualmente están inervadas por una sola terminación nerviosa localizada cerca del punto medio de la fibra
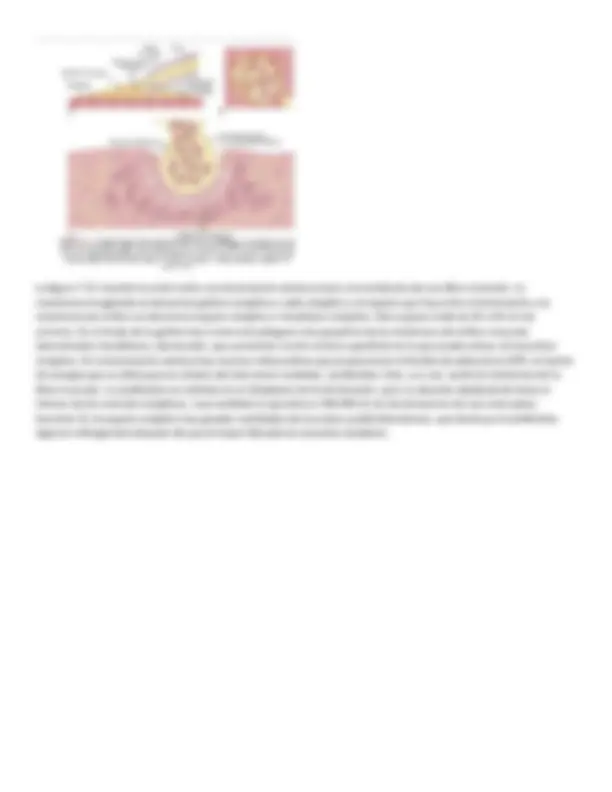
El sarcolema es una fina membrana que envuelve a una fibra musculoesquelética El sarcolema está formado por una membrana celular verdadera, denominada membrana plasmática, y una cubierta externa formada por una capa delgada de material polisacárido que contiene numerosas fibrillas delgadas de colágeno. En cada uno de los dos extremos de la fibra muscular la capa superficial del sarcolema se fusiona con una fibra tendinosa. Las fibras tendinosas a su vez se agrupan en haces para formar los tendones musculares, que después insertan los músculos en los huesos. Las miofibrillas están formadas por filamentos de actina y miosina Cada fibra muscular contiene varios cientos a varios miles de miofibrillas, que se representan en la vista en sección transversal de la figura 6-1C. Cada miofibrilla (v. fig. 6-1D y E) está formada por aproximadamente 1.500 filamentos de miosina y 3.000 filamentos de actina adyacentes entre sí, que son grandes moléculas proteicas polimerizadas responsables de la contracción muscular real. Estos filamentos se pueden ver en una imagen longitudinal en la microfotografía electrónica de la figura 62 y se representan esquemáticamente en la figura 6-1, partes E a L. Los filamentos gruesos de los diagramas son miosina y los filamentos delgados son actina. Obsérvese en la figura 6-1E que los filamentos de miosina y de actina se interdigitan parcialmente y de esta manera hacen que las miofibrillas tengan bandas claras y oscuras alternas, como se ilustra en la figura 6-2. Las bandas claras contienen solo filamentos de actina y se denominan bandas I porque son isótropas a la luz polarizada. Las bandas oscuras contienen filamentos de miosina, así como los extremos de los filamentos de actina en el punto en el que se superponen con la miosina, y se denominan bandas A porque son anisótropas a la luz polarizada. Obsérvense también las pequeñas proyecciones que se originan en los lados de los filamentos de miosina en la figura 6-1E y L, y que se denominan puentes cruzados. La interacción entre estos puentes cruzados y los filamentos de actina produce la contracción. La figura 6-1E también muestra que los extremos de los filamentos de actina están unidos al disco Z. Desde este disco estos filamentos se extienden en ambas direcciones para interdigitarse con los filamentos de miosina. El disco Z, que está formado por proteínas filamentosas distintas de los filamentos de actina y miosina, atraviesa las miofibrillas y también pasa desde unas miofibrillas a otras, uniéndolas entre sí a lo largo de toda la longitud de la fibra muscular. Por tanto, toda la fibra muscular tiene bandas claras y oscuras, al igual que las miofibrillas individuales. Estas bandas dan al músculo esquelético y cardíaco su aspecto estriado. La porción de la miofibrilla (o de la fibra muscular entera) que está entre dos discos Z sucesivos se denomina sarcómero. Cuando la fibra muscular está contraída, como se muestra en la parte inferior de la figura 6-5, la longitud del sarcómero es de aproximadamente 2 μm. Cada una de estas fibras está formada por m. Cuando el sarcómero tiene esta longitud, los filamentos de actina se superponen completamente con los filamentos de miosina y las puntas de los filamentos de actina están comenzando ya a superponerse entre sí. Como se expone más adelante, a esta longitud el músculo es capaz de generar su máxima fuerza de contracción. Las moléculas filamentosas de titina mantienen en su lugar los filamentos de miosina y actina La relación de yuxtaposición entre los filamentos de miosina y de actina se mantiene por medio de un gran número de moléculas filamentosas de una proteína denominada titina (fig. 6-3). Cada molécula de titina tiene un peso molecular de aproximadamente 3 millones, lo que hace que sea una de las mayores moléculas proteicas del cuerpo. Además, como es filamentosa, es muy elástica. Estas moléculas elásticas de titina actúan como armazón que mantiene en su posición los filamentos de miosina y de actina, de modo que funcione la maquinaria contráctil del sarcómero. Un extremo de la molécula de titina es elástico y está unido al disco Z; para actuar a modo de muelle y con una longitud que cambia según el sarcómero se contrae y se relaja. La otra parte de la molécula de titina la une al grueso filamento de miosina. La
Los filamentos de miosina están compuestos por múltiples moléculas de miosina Cada una de las moléculas de miosina, mostradas en la figura 6-6A, tiene un peso molecular de aproximadamente 480.000. La figura 6-6B muestra la organización de muchas moléculas para formar un filamento de miosina, así como la interacción de este filamento por un lado con los extremos de dos filamentos de actina La molécula de miosina (v. fig. 6-6A) está formada por seis cadenas polipeptídicas, dos cadenas pesadas, cada una de las cuales tiene un peso molecular de aproximadamente 200.000, y cuatro cadenas ligeras, que tienen un peso molecular de aproximadamente 20.000 cada una. Las dos cadenas pesadas se enrollan entre sí en espiral para formar una hélice doble, que se denomina cola de la molécula de miosina. Un extremo de cada una de estas cadenas se pliega bilateralmente para formar una estructura polipeptídica globular denominada cabeza de la miosina. Así, hay dos cabezas libres en un extremo de la molécula de miosina de doble hélice. Las cuatro cadenas ligeras también forman parte de la cabeza de la miosina, dos en cada cabeza. Estas cadenas ligeras ayudan a controlar la función de la cabeza durante la contracción muscular. El filamento de miosina está formado por 200 o más moléculas individuales de miosina. En la figura 6-6B se muestran la porción central de uno de estos filamentos y las colas de las moléculas de miosina agrupadas entre sí para formar el cuerpo del filamento, mientras que hay muchas cabezas de las moléculas por fuera de los lados del cuerpo. Además, parte del cuerpo de cada una de las moléculas de miosina se prolonga hacia la región lateral junto a la cabeza, formando de esta manera un brazo que separa la cabeza del cuerpo, como se muestra en la figura. Los brazos y las cabezas que protruyen se denominan en conjunto puentes cruzados. Cada puente cruzado es flexible en dos puntos denominados bisagras, una en el punto en el que el brazo sale del cuerpo del filamento de miosina y la otra en el punto en el que la cabeza se une al brazo. Los brazos articulados permiten que las cabezas se separen del cuerpo del filamento de miosina o que se aproximen a este. Las cabezas articuladas, a su vez, participan en el proceso real de contracción, como se analiza en las secciones siguientes. La longitud total de los filamentos de miosina es uniforme, casi exactamente 1,6 μm. Cada una de estas fibras está formada por m. Sin embargo, se debe tener en cuenta que no hay cabezas de puentes cruzados en el centro del filamento de miosina en una distancia de aproximadamente 0,2 μm. Cada una de estas fibras está formada por m, porque los brazos articulados se separan desde el centro. Ahora, para completar este cuadro, el filamento de miosina está enrollado de modo que cada par sucesivo de puentes cruzados está desplazado en sentido axial 120° con respecto al par previo. Esta torsión garantiza que los puentes cruzados se extiendan en todas las direcciones alrededor del filamento. Energética de la contracción muscular Generación de trabajo durante la contracción muscular Cuando un músculo se contrae contra una carga realiza un trabajo. Esto significa que se transfiere energía desde el músculo hasta la carga externa para levantar un objeto hasta una mayor altura o para superar la resistencia al movimiento. En términos matemáticos el trabajo se define mediante la siguiente ecuación: donde T es el trabajo generado, C es la carga y D es la distancia del movimiento que se opone a la carga. La energía necesaria para realizar el trabajo procede de las reacciones químicas de las células musculares durante la contracción, como se describe en las secciones siguientes. Tres fuentes de energía para la contracción muscular La mayor parte de la energía necesaria para la contracción muscular se utiliza para activar el mecanismo de cremallera mediante el cual los puentes cruzados tiran de los filamentos de actina, aunque son necesarias cantidades pequeñas para: 1) bombear iones calcio desde el sarcoplasma hacia el interior del retículo sarcoplásmico después de que haya finalizado la contracción, y 2) para bombear iones sodio y potasio a través de la membrana de la fibra muscular para mantener un entorno iónico adecuado para la propagación de los potenciales de acción de la fibra muscular. La contracción de ATP en la fibra muscular, de aproximadamente 4 milimolar, es suficiente para mantener la contracción completa durante solo 1 a 2 s como máximo. El ATP se escinde para formar ADP, que transfiere la energía de la molécula de ATP a la maquinaria contráctil de la fibra muscular. Después, como se describe en el capítulo 2, el ADP se vuelve a fosforilar para formar nuevo ATP en otra fracción de segundo, lo que permite que el músculo mantenga su contracción.
Hay varias fuentes de energía para esta nueva fosforilación. La primera fuente de energía que se utiliza para reconstituir el ATP es la sustancia fosfocreatina, que contiene un enlace fosfato de alta energía similar a los enlaces del ATP. El enlace fosfato de alta energía de la fosfocreatina tiene una cantidad ligeramente mayor de energía libre que la de cada uno de los enlaces del ATP, como se analiza con más detalle en los capítulos 68 y 73. Por tanto, la fosfocreatina se escinde inmediatamente y la energía que se libera produce el enlace de un nuevo ion fosfato al ADP para reconstituir el ATP. Sin embargo, la cantidad total de fosfocreatina en la fibra muscular también es pequeña, solo aproximadamente cinco veces mayor que la de ATP. Por tanto, la energía combinada del ATP y de la fosfocreatina almacenados en el músculo es capaz de producir una contracción muscular máxima durante solo 5 a 8 s. La segunda fuente importante de energía, que se utiliza para reconstituir tanto el ATP como la fosfocreatina, es la «glucólisis» del glucógeno que se ha almacenado previamente en las células musculares. La escisión enzimática rápida del glucógeno en ácido pirúvico y ácido láctico libera energía que se utiliza para convertir el ADP en ATP; después se puede utilizar directamente el ATP para aportar energía a la contracción muscular adicional y también para reconstituir los almacenes de fosfocreatina. La importancia de este mecanismo de glucólisis es doble. Primero, las reacciones glucolíticas se pueden producir incluso en ausencia de oxígeno, de modo que se puede mantener la contracción muscular durante muchos segundos y a veces hasta más de 1 min, aun cuando no se disponga de aporte de oxígeno desde la sangre. Segundo, la velocidad de formación de ATP por el proceso glucolítico es aproximadamente 2,5 veces más rápida que la formación de ATP en respuesta a la reacción de los nutrientes celulares con el oxígeno. Sin embargo, se acumulan tantos productos finales de la glucólisis en las células musculares que la glucólisis también pierde su capacidad de mantener una contracción muscular máxima después de aproximadamente 1 min. La tercera y última fuente de energía es el metabolismo oxidativo, lo que supone combinar oxígeno con los productos finales de la glucólisis y con otros diversos nutrientes celulares para liberar ATP. Más del 95% de toda la energía que utilizan los músculos para la contracción sostenida a largo plazo procede del metabolismo oxidativo. Los nutrientes que se consumen son hidratos de carbono, grasas y proteínas. Para una actividad muscular máxima a muy largo plazo (durante un período de muchas horas) la mayor parte de la energía procede con mucho de las grasas, aunque durante períodos de 2 a 4 h hasta la mitad de la energía puede proceder de los hidratos de carbono almacenados. Los mecanismos detallados de estos procesos energéticos se analizan en los capítulos 67 a 72. Además, en el capítulo 84 se expone la importancia de los diferentes mecanismos de liberación de energía durante la realización de diferentes deportes. Características de la contracción de todo el músculo Muchas características de la contracción muscular se pueden demostrar desencadenando espasmos musculares únicos. Esto se puede conseguir con la excitación eléctrica instantánea del nervio que inerva un músculo o haciendo pasar un estímulo eléctrico breve a través del propio músculo, dando lugar a una única contracción súbita que dura una fracción de segundo Las contracciones isométricas no acortan el músculo, mientras que las contracciones isotónicas lo acortan a una tensión constante Se dice que la contracción muscular es isométrica cuando el músculo no se acorta durante la contracción e isotónica cuando se acorta, pero la tensión del músculo permanece constante durante toda la contracción. En la figura 6-12 se muestran sistemas para registrar los dos tipos de contracción muscular
músculos que, como el sóleo, responden lentamente pero con una contracción prolongada están formados principalmente por fibras «lentas». Las diferencias entre estos dos tipos de fibras se describen en el siguiente apartado. Fibras lentas (tipo I, músculo rojo) Las características de las fibras lentas son las siguientes: 1. Las fibras son más pequeñas que las fibras rápidas. 2. Las fibras lentas están también inervadas por fibras nerviosas más pequeñas. 3. En comparación con las fibras rápidas, las fibras lentas tienen un sistema de vascularización más extenso y más capilares para aportar cantidades adicionales de oxígeno. 4. Las fibras lentas tienen números muy elevados de mitocondrias, también para mantener niveles elevados de metabolismo oxidativo. 5. Las fibras lentas contienen grandes cantidades de mioglobina, una proteína que contiene hierro y que es similar a la hemoglobina de los eritrocitos. La mioglobina se combina con el oxígeno y lo almacena hasta que sea necesario, lo cual acelera también notablemente el transporte de oxígeno hacia las mitocondrias. La mioglobina da al músculo lento un aspecto rojizo y el nombre de músculo rojo. Fibras rápidas (tipo II, músculo blanco) Las características de las fibras rápidas son: 1. Las fibras rápidas son grandes para obtener una gran fuerza de contracción. 2. Existe un retículo sarcoplásmico extenso para una liberación rápida de iones calcio al objeto de iniciar la contracción. 3. Están presentes grandes cantidades de enzimas glucolíticas para la liberación rápida de energía por el proceso glucolítico. 4. Las fibras rápidas tienen una vascularización menos extensa que las lentas, porque el metabolismo oxidativo tiene una importancia secundaria. 5. Las fibras rápidas tienen menos mitocondrias que las lentas, también porque el metabolismo oxidativo es secundario. Un déficit de mioglobina roja en el músculo rápido le da el nombre de músculo blanco. Mecánica de la contracción del músculo esquelético Unidad motora: todas las fibras musculares inervadas por una única fibra nerviosa Todas las motoneuronas que salen de la médula espinal inervan múltiples fibras nerviosas y el número de fibras inervadas depende del tipo de músculo. Todas las fibras musculares que son inervadas por una única fibra nerviosa se denominan unidad motora (fig. 6-14). En general, los músculos pequeños que reaccionan rápidamente y cuyo control debe ser exacto tienen más fibras nerviosas para menos fibras musculares (p. ej., tan solo dos o tres fibras musculares por cada unidad motora en algunos de los músculos laríngeos). Por el contrario, los músculos grandes que no precisan un control fino, como el músculo sóleo, pueden tener varios centenares de fibras musculares en una unidad motora. Una cifra promedio para todos los músculos del cuerpo es cuestionable, aunque una buena estimación sería de aproximadamente 80 a 100 fibras musculares por unidad motora. Las fibras musculares de todas las unidades motoras no están agrupadas entre sí en el músculo, sino que se superponen a otras unidades motoras en microfascículos de 3 a 15 fibras. Esta interdigitación permite que las unidades motoras separadas se contraigan cooperando entre sí y no como segmentos totalmente individuales Contracciones musculares de diferente fuerza: sumación de fuerzas Sumación significa la adición de los espasmos individuales para aumentar la intensidad de la contracción muscular global. La sumación se produce de dos maneras: 1) aumentando el número de unidades motoras que se contraen de manera simultánea, lo que se denomina sumación de fibras múltiples, y 2) aumentando la frecuencia de la contracción, lo que se denomina sumación de frecuencia y que puede producir tetanización Tono del músculo esquelético Incluso cuando los músculos están en reposo habitualmente hay una cierta cantidad de tensión, que se denomina tono muscular. Como las fibras normales del músculo esquelético no se contraen sin que ningún potencial de acción estimule las fibras, el tono del músculo esquelético se debe totalmente a impulsos nerviosos de baja frecuencia que proceden de
la médula espinal. Estos impulsos nerviosos, a su vez, están controlados en parte por señales que se transmiten desde el encéfalo a las motoneuronas adecuadas del asta anterior de la médula espinal y en parte por señales que se originan en los husos musculares que están localizados en el propio músculo. Hipertrofia y atrofia muscular El aumento de la masa total de un músculo se denomina hipertrofia muscular. Cuando la masa total disminuye, el proceso recibe el nombre de atrofia muscular. Prácticamente toda la hipertrofia muscular se debe a un aumento del número de filamentos de actina y miosina en cada fibra muscular, dando lugar a aumento de tamaño de las fibras musculares individuales; esta situación se denomina hipertrofia de las fibras. La hipertrofia aparece en un grado mucho mayor cuando el músculo está sometido a carga durante el proceso contráctil. Solo son necesarias unas pocas contracciones intensas cada día para producir una hipertrofia significativa en un plazo de 6 a 10 semanas. Se desconoce el mecanismo por el cual una contracción intensa produce hipertrofia. Sin embargo, se sabe que la velocidad de síntesis de las proteínas contráctiles del músculo es mucho mayor cuando se está produciendo la hipertrofia, lo que da lugar también a números cada vez mayores de filamentos tanto de actina como de miosina en las miofibrillas, aumentando con frecuencia hasta un 50%. A su vez, se ha observado que algunas de las miofibrillas se dividen en el interior del músculo que se está hipertrofiando para formar nuevas miofibrillas, aunque todavía no se sabe la importancia de este proceso en la hipertrofia muscular que se ve habitualmente. Junto con el aumento de tamaño de las miofibrillas, también se produce un aumento de los sistemas enzimáticos que proporcionan energía. Este aumento se aplica especialmente a las enzimas de la glucólisis, lo que permite el aporte rápido de energía durante la contracción muscular intensa a corto plazo. Cuando un músculo no se utiliza durante muchas semanas, la velocidad de degradación de las proteínas contráctiles es mucho más rápida que la velocidad de sustitución. Por tanto, se produce atrofia muscular. La ruta que parece importar en buena parte para la degradación proteica en un músculo que experimenta atrofia es la ruta de ubicuitina-proteasoma dependiente del ATP. Los proteasomas son grandes complejos proteicos que degradan las proteínas dañadas o innecesarias por proteólisis, una reacción química que rompe los enlaces peptídicos. La ubicuitina es una proteína reguladora que básicamente marca las células que serán diana para una degradación proteasómica Hiperplasia de las fibras musculares En situaciones poco frecuentes de generación extrema de fuerza muscular se ha observado que hay un aumento real del número de fibras musculares (aunque solo en un pequeño porcentaje), además del proceso de hipertrofia de las fibras. Este aumento del número de fibras se denomina hiperplasia de las fibras. Cuando aparece, el mecanismo es la división lineal de fibras que estaban previamente aumentadas de tamaño. CAPÍTULO 7 Excitación del músculo esquelético: transmisión neuromuscular y acoplamiento excitacióncontracción Transmisión de impulsos desde las terminaciones nerviosas a las fibras del músculo esquelético: la unión neuromuscular Las fibras del músculo esquelético están inervadas por fibras nerviosas mielinizadas grandes que se originan en las motoneuronas grandes de las astas anteriores de la médula espinal. Como se expone en el capítulo 6, todas las fibras nerviosas, después de entrar en el vientre muscular, normalmente se ramifican y estimulan entre tres y varios cientos de fibras musculares esqueléticas. Cada terminación nerviosa forma una unión, denominada unión neuromuscular, con la fibra muscular cerca de su punto medio. El potencial de acción que se inicia en la fibra muscular por la señal nerviosa viaja en ambas direcciones hacia los extremos de la fibra muscular. Con la excepción de aproximadamente el 2% de las fibras musculares, solo hay una unión de este tipo en cada fibra muscular. Anatomía fisiológica de la unión neuromuscular: la placa motora terminal La figura 7-1A y B muestra la unión neuromuscular que forma una gran fibra nerviosa mielinizada con una fibra muscular esquelética. La fibra nerviosa forma un complejo de terminaciones nerviosas ramificadas que se invaginan en la superficie de la fibra muscular, pero que permanecen fuera de la membrana plasmática. Toda la estructura se denomina placa motora terminal. Está cubierta por una o más células de Schwann que la aíslan de los líquidos circundantes.