¡Descarga Compartimentación celular y más Apuntes en PDF de Biología Celular solo en Docsity!
TEMA 1. COMPARTIMENTACIÓN CELULAR.
La compartimentación de la célula eucariota: principales compartimentos de la célula
animal.
La compartimentación de la célula eucariota:
Que la célula tenga distintos compartimentos quiere decir que es una célula especializada , y que para las distintas funciones que realiza utiliza distintos compartimentos, los cuales están regulados por membranas. El citoplasma es todo el contenido celular que se encuentra entre la membrana plasmática y el núcleo. El citosol es el contenido del citoplasma con exclusión de todos los orgánulos limitados por membrana, y los elementos del citoesqueleto. Observamos que el número aproximado por célula de vesículas del ER rugoso es solamente uno, y esto es así porque el interior de las cisternas del retículo está comunicado entre sí formando un solo compartimento. En número aproximado por célula de vesículas del ER liso y de Golgi no pone nada porque forman, junto con el rugoso, un único compartimento.
Membranas celulares:
Las membranas celulares son estructuras que posibilitan que existan diferentes compartimentos celulares, con
diferentes funciones. Por ejemplo, para la célula dos compartimentos pueden ser el ambiente extracelular y el
ambiente intracelular, siendo la membrana celular la estructura que regula el paso de sustancias entre estos dos
compartimentos.
Definiremos membrana celular como una delgada lámina de una doble capa de moléculas lipídicas y proteínas
asociadas, que rodea a todas las células y a muchos orgánulos de las células eucariotas.
Tipos de membranas celulares:
Membrana plasmática (también llamada a veces membrana celular a secas).
Membranas endocelulares.
M. externa e interna nuclear.
M. externa e interna mitocondrial.
M. externa e interna plastidial.
M. del peroxisoma.
M. del retículo endoplasmático, del aparato de Golgi, gránulos de secreción, lisosomas y vacuolas
(presentes solamente en las células vegetales).
M. del endosoma.
Las membranas no pueden observarse a microscopía óptica, sino electrónica La bicapa lipídica o tricapa de la membrana constituye líneas paralelas (que nunca se juntan), pero forma únicamente una sola membrana plasmática.
Las colas de los fosfolípidos pueden ser de diferentes longitudes (normalmente de 14 a 24 átomos de carbono) y
normalmente una de ellas presenta uno o más dobles enlaces cis (es decir, es insaturada ) mientras que la otra
normalmente no tiene dobles enlaces (es decir, es saturada ). Tal y como se aprecia en la figura, cada doble
enlaces cis genera una curvatura suave en la cadena.
Es la naturaleza anfipática de las moléculas lipídicas lo que causa la formación espontánea de bicapas lipídicas en
solución acuosa. Cuando las moléculas anfipáticas se encuentran rodeadas por todas partes por un ambiente
acuoso, tienden a agregarse de tal manera que sus colas hidrofóbicas se escondan en el interior y sus cabezas
hidrofílicas queden expuestas al agua.
Estas diferencias de longitud y de grado de
saturación entre las colas son importantes
porque afectan a la capacidad de las
moléculas de fosfolípidos a empaquetarse
una contra otra y, por lo tanto, afectan a la
fluidez de la membrana.
Composición química: el colesterol.
Otro determinante de la fluidez de una membrana es el colesterol. Las membranas plasmáticas de los eucariotas
contienen cantidades relativamente elevadas de colesterol. Además de regular la fluidez, se cree que el colesterol
aumenta la estabilidad mecánica de la bicapa. Las moléculas de colesterol se orientan en la bicapa con su grupo
hidroxilo próximo a las cabezas polares de las moléculas de fosfolípidos; sus anillos esteroides planos y rígidos
interactúan – y en parte inmovilizan – con una parte de los grupos apolares de la cola hidrocarbonada, dejando el
resto de la cadena más flexible. Aunque de esta manera el colesterol tiende a hacer menos fluidas las bicapas
lipídicas, a las altas concentraciones en que se presenta en la mayoría de las membranas plasmáticas de los
eucariotas también ejerce efecto de impedir que las cadenas hidrocarbonadas se junten y cristalicen.
Cuando la cantidad de colesterol es sobrante en nuestro organismo, este se deposita en las membranas de las
células que forman los vasos sanguíneos, impidiendo por tanto la correcta dilatación y contracción del mismo y
disminuyendo pues el riego sanguíneo, esto puede provocar demencia senil. Es pues importantísima la dieta que
tomemos.
En un entorno acuoso los fosfolípidos forman espontáneamente
bicapas. Además, estas bicapas lipídicas tienden a cerrarse sobre
sí mismas formando compartimentos herméticos y eliminando
así los bordes libres en los que las colas hidrofóbicas podrían
estar en contacto con el agua. Por esta misma razón, los
compartimentos formados por bicapas lipídicas tienden a
cerrarse de nuevo después de haber sido rotos.
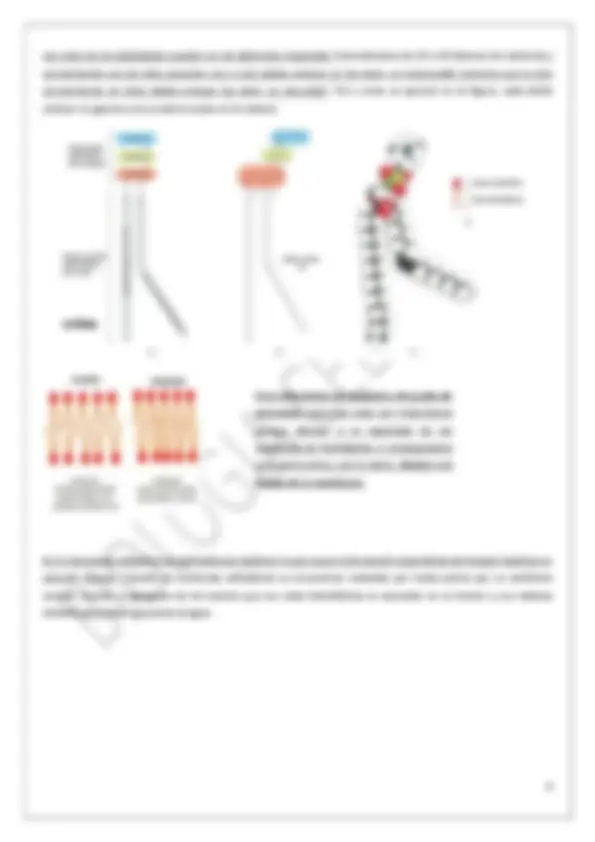
Un fosfolípido mide 3 nm, y como la bicapa lipídica son capas formadas por dos fosfolípidos en unidad pues ésta mide 6 nm. Colesterol interactuando con fosfolípidos
Tipos de proteínas de membrana:
Las células pueden limitar la movilidad de las proteínas de membrana:
Las proteínas integrales son iguales que las periféricas pero no se encuentran en la cara externa, solo en la interna , y no se encuentran asociadas a otras proteínas de membrana sino a la bicapa lipídica. Se pueden separar de la misma rompiendo la bicapa con detergentes o solventes orgánicos. La existencia de esta unión estrecha entre las células debido a su estructura permite que las proteínas que están en la parte apical de la célula no pasen a la basolateral y, tal y como dice la imagen, se favorece de este modo que la célula se polarice estructural y funcionalmente.
Además de la unión estrecha a otra célula o unión hermética, existen otros
mecanismos para la limitación de la movilidad de las proteínas:
- Unidas a proteínas internas.
- Unidas a proteínas externas.
- Unidas a proteínas de otras células.
- Por uniones herméticas (de las cuales ya hemos hablado arriba un
poco).
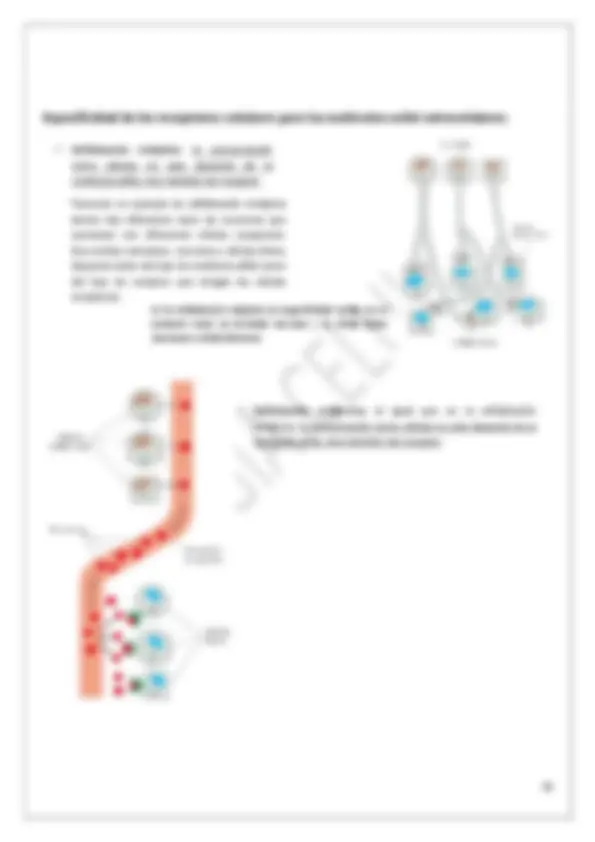
Proteínas receptoras de membrana:
Receptores acoplados a canales iónicos: Receptores acoplados a proteínas G: Receptores acoplados a enzimas:
Funciones de las membranas:
Aíslan y relacionan a la célula con el exterior, o al orgánulo con el citosol.
Separan medios con distinta composición química. El citosol con el exterior, o el orgánulo con el
citosol.
Son semipermeables y permiten el transporte de sustancias de un lado a otro de la membrana.
Actúan de soporte no aleatorio de enzimas.
La membrana plasmática presenta uniones intercelulares.
La membrana plasmática permite el reconocimiento celular y molecular mediante receptores.
Hay receptores acoplados a canales iónicos. Estos receptores permiten la entrada de los iones si se les une un sustrato específico. Mientras el receptor no se una al ligando los tres permanecen separados. Si llega la sustancia que activa al receptor, este se une a la proteína G activándola, y por consiguiente esta última activa a la enzima. La unión de la molécula señal con su correspondiente receptor va a llevar consigo la catálisis de una reacción enzimática.
Transporte a través de membrana:
Difusión simple (transporte inespecífico): transporte de sustancias como H 2 O, O 2 , CO 2 ,
esteroides, moléculas liposolubles, urea, glicerina, etc… a favor del gradiente electroquímico y sin
gasto de energía.
Transporte específico: de iones y de pequeñas moléculas. Proteínas canales (proteínas poro): transportan iones a favor del gradiente electroquímico y
sin gasto de energía.
Proteínas transportadoras: transportan iones y pequeñas moléculas bien por transporte
pasivo , a favor del gradiente y sin gasto de energía, bien por transporte activo , es decir, en contra
del gradiente y con gasto de energía (ej. Bomba de Na+/K*).
Algunas sustancias como el H 2 O, el O 2 , CO 2 , esteroides, vitaminas liposolubles, urea, glicerina, alcoholes de pequeño peso molecular, atraviesan la membrana celular por difusión simple , disolviéndose en la capa de fosfolípidos, a favor de gradiente de concentración y sin gasto de energía. El movimiento del agua a través de la membrana semipermeable genera sobre la membrana una presión osmótica.
Todas las membranas de la célula
son semipermeables.
La célula tiene que garantizar las diferencias de concentración de estas moléculas
El transporte puede ser bien por transporte inespecífico :
difusión , bien por transporte específico : transporte pasivo
(o mediado por canal o mediado por transportador), o
bien por transporte activo mediado por transportador.
Transporte mediado por proteínas transportadoras: la unión del ligando con su receptor hace que éste
último experimente un cambio conformacional y pueda transportar el ligando al interior o al exterior
celular.
Tipos de transportadores: Uniporte: El transportador transporta solo una molécula a favor del gradiente de concentración. Simporte: tipo de transporte por el cual se transporta una molécula en contra del gradiente acoplando en este trasiego el desplazamiento de uno o más iones diferentes a favor del suyo. Antiporte: tipo de transporte por el cual el transportador transporta dos moléculas a favor de su gradiente, siendo ambos gradientes contrapuestos.
Transporte activo por ión cotransportado: Regulación de canales iónicos en la unión neuromuscular: Transporte de glucosa en contra del gradiente acoplado al transporte de iones sodio a favor del gradiente electroquímico (SIMPORTE) El transportador de transporte activo por ión cotransportado solo se encuentra en la membrana apical de las células. En el lumen del intestino se encuentra la glucosa de todos los polisacáridos que hemos ingerido y estos pasan a las células, para luego ir al torrente sanguíneo y de ahí a todo el cuerpo, por este tipo de transporte, después pasan al torrente sanguíneo por transporte pasivo (a favor del gradiente).
UNIÓN NEUROMUSCULAR INACTIVA
UNIÓN NEUROMUSCULAR ACTIVA
Transporte mediado por proteínas poro o canal : la característica fundamental de este grupo es que
NO se requiere un cambio conformacional de las proteínas para que se efectúe el transporte. Las
proteínas crean un conducto hidrofílico que atraviesa la membrana, conducto que puede ser atravesado
por solutos polares o iónicos. Estas proteínas pueden cambiar de conformación y abrirse o cerrarse, pero
una vez abiertos los solutos transportados atraviesan el canal por un proceso de difusión, como si
estuviesen libres en disolución.
Mecanismos de regulación de la apertura de los canales iónicos: La célula siempre tiene más cargas positivas fuera que dentro, entre el interior y el exterior de la célula existe pues una diferencia de potencial de 120 mV. Los canales regulados por voltaje abren y cierran sus compuertas en respuesta a un cambio en la diferencia de potencial. Los canales regulados mecánicamente se abren o se cierran en respuesta a la fuerza mecánica (receptores de presión) aplicada sobre la membrana plasmática. En primer lugar no hay trasmisión del impuso nervioso, sino reposo. Existen vesículas en el interior del final del axón que llevan neurotransmisores. El canal de calcio regulado por voltaje hace cambiar la polaridad de la membrana, liberándose pues los neurotransmisores. Estos neurotransmisores activan los canales sodio, que hacen que entre sodio al interior celular (que puede ser una neurona) y esto hace que el retículo endoplasmático libere calcio al interior celular provocando la contracción muscular. El grado de selectividad de estas moléculas es muy variable: hay poros que pueden ser muy poco selectivos, permitiendo el paso de cualquier soluto suficientemente pequeño para atravesar su interior, mientras que en otros casos permiten el paso sólo de un soluto determinado.
TEMA 2. PRINCIPIOS GENERALES DE LA
COMUNICACIÓN CELULAR.
Señalización intercelular:
Las células tienen que comunicarse entre sí para formar y mantener un organismo pluricelular.
Esta comunicación se realiza a través de:
Uniones gap: median la comunicación intercelular permitiendo el
paso de iones inorgánicos y de otras pequeñas moléculas
hidrosolubles desde el citoplasma de una célula al citoplasma de
otra, acoplando así las células tanto eléctrica como
metabólicamente. Este acoplamiento celular tiene importantes
implicaciones funcionales.
Contacto directo célula – célula : tiene lugar
entre una célula señalizadora y una célula
diana contigua. La célula diana puede ser del
mismo tipo que la célula señalizadora, o
diferente.
Molécula señal extracelular: la comunicación
tiene lugar entre células distantes por
moléculas señal extracelulares. Las producen
células señalizadoras y actúan sobre células
diana, iguales o diferentes, activando en estas
una o más vías intracelulares de señalización.
Formas de comunicación intercelular mediante moléculas señal extracelulares:
Señalización autocrina: es la que establece una célula consigo misma.
Muchas células en crecimiento como las células del embrión o las células
cancerosas producen factores de crecimiento y los receptores para esos
mismos factores de crecimiento y así perpetuar su proliferación,
controlada en el caso del embrión y descontrolada en el caso del cáncer.
Señalización paracrina: una célula envía
señales a otras células, iguales o diferentes, de
su entorno. Un tipo especial de señalización
paracrina es la que utiliza neurotransmisores,
moléculas señal en la sinapsis entre neuronas
o entre neuronas y células efectoras.
Señalización endocrina: la molécula
señal (hormona) es segregada por
una célula endocrina, liberada a la
sangre y transportada hasta las
células diana que puede estar a una
cierta distancia.
Receptores de superficie e intracelulares:
Un receptor es cualquier proteína que se une de forma específica a una molécula señal (ligando), lo que inicia una
respuesta en la célula. Los hay que se encuentran en la superficie celular, o dentro de la célula.
Las moléculas señal extracelulares se comportan de manera diferente según sean hidrófilas o hidrófobas.
Hidrófilas: tienen receptores de superficie.
Hidrófobas: requieren de proteínas transportadoras y un receptor intracelular, citosólico o nuclear.
Respuestas señalizadoras:
Las señales lentas producen cambios en la expresión de genes, lo que induce a la síntesis de nuevas proteínas.
Respuesta de una célula diana a diferentes moléculas señal extracelulares:
Cada tipo celular posee un conjunto de receptores de superficie e intracelulares para distintos tipos de moléculas
señal (representadas como A, B, C, D, E, F, G). Estas moléculas actúan de forma combinada y regulan el
comportamiento de la célula.
Diferentes tipos celulares responden de forma diferente a la misma molécula señal:
Las señales rápidas producen cambios en proteínas de la célula, alterando su función. Por ejemplo, fosforilación de proteínas.