¡Descarga Compartiments cel.lulars y más Apuntes en PDF de Biología Celular solo en Docsity!
8. COMPARTIMENTS CEL·LULARS
En una cèl·lula eucariota en un moment determinat es fan reaccions químiques contràries, com ara formació i degradació de proteïnes, tot alhora. Aquestes reaccions es fan en compartiments diferents Æ orgànuls envoltats per membranes.
8.1. ORGÀNULS DELIMITATS PER MEMBRANES
- Citosol : solució aquosa on hi ha tots aquests orgànuls. Hi ha molta activitat metabòlica.
- Nucli: S’hi fa la síntesi, la transcripció, etc... de DNA. El mat. genètic té forma de eucromatina o heterocromatina (que no es transcriu). Està envoltat per una doble membrana (embolcall nuclear). La mb nuclear té ribosomes enganxats. En els porus nuclears hi ha eucromatina. Hi ha una continüitat entre la mb externa del nucli i el RE.
- RER: Síntesi de proteïnes (més abundant que REL)
- REL: Síntesi de lípids i extreu molècules tòxiques.
- Aparell de Golgi: Estructura que s’organitza en dictiosomes (compartiments), en els quals hi ha tràfic de molècules. → Cara cis: més propera al reticle. És per on entren les proteïnes i els lípids. → Cara trans: per on surten les anteriors modificades; les deriva cap a on hagin d’anar mitjançant vesícules.
- Lisosomes: Degrada material d’endocitosi i els propis orgànuls de cèls: autofàgia.
- Endosomes: sacs irregulars que s’encarreguen de classificar el material d’endocitosi (si van a lisosomes per degradar-se o no).
- Mitocondris: sintetizar ATP. Divisió independent.
----microvellositats
- Cloroplasts (cèls. vegetals): Es divideixen per partició (mitocondris també) i fan síntesi d’ATP: el CO2 atmosfèric es fixa en forma de molècula de carboni orgànic i té lloc la síntesi d’ATP.
- Peroxisomes: orgànuls esfèrics relacionats amb l’oxidació de molècules (tòxiques). Aquesta energia s’escapa en forma de calor. També fan divisió independent. S’hi oxiden molts alcohols.
- La composició molecular de cada orgànul és diferent, segons la funció que tingui.
- Cada orgànul manté una posició bastant uniforme dins la cèl·lula determinada pel citoesquelet.
- Alguns dels orgànuls estan connectats i es passen material entre ells mitjançant tràfic de vesícules. Si el trajecte és curt, les vesícules fan difusió. Si es llarg, faran servir proteïnes motores del citoesquelet associades, com actina, microtúbuls...
8.1.1. Origen evolutiu
Els orgànuls de les cèl·lules tenen orígens diferents:
Nucli RE (A) Invaginació de la membrana d’una cèl·lula A.Golgi procariota primitiva Æ els orgànuls estan més connectats entre sí. Endosomes Lisosomes
Mitocondris (B) Van ser originats a partir d’endocitosis La cèl·lula procariota Cloroplasts primitiva va endocitar altres procariotes, i aleshores va permetre fer Peroxisomes fotosíntesis (cloroplasts) o sintetitzar ATP (mitocondris).
Un cop dins, els gens d’aquests aeròbics primitius van passar al nucli. Tenen divisió independent de la cèl·lula (ja que és una resta del que feien abans), però per créixer depenen de la cel·lula totalment. Degut a que els processos de endosimbiosi van ser diferents, no estan connectats entre ells ni amb els del primer grup.
que l’ambient sigui hidrofílic. Les proteïnes s’hauran de desplegar perquè els canals són més estrets.
- Transport per mitjà de vesícules Ens portarà proteïnes a AG, endosomes, lisosomes, membrana plasmàtica i exterior cel·lular. L’orgànul donador deixa anar petites vesícules (peces) de la membrana que tenen material i es fusionen amb l’orgànul de destí (acceptador). Les vesícules contenen lípids i proteïnes. Els mecanismes de reconeixement depenen de les proteïnes Snare.
- Com arriben les proteïnes al embolcall nuclear? Abans han hagut de passar per RE. Simplement faran difusió lateral des del reticle fins l’embolcall nuclear.
- Com saben a quin orgànul han d’anar? Segons la seva seqüència senyal.
8.1.3. Seqüència senyal Una part de la seqüència d’aa de la proteïna (seqüència senyal) és com l’etiqueta amb l’adreça; especifica el destí de la proteïna. Mateix orgànul Æmateixes seqüències senyal o amb característiques físico-químiques semblants:
- Hidrofòbics
- Retenció al lumenÆsempre iguals (KDEL)
- Mitocondris: positius alter- nats amb negatius
Les proteïnes que tenen funció al citoplasma no tenen seqüència senyal, ja que es troben en el seu lloc d’ús. Si la seq. senyal es troba en posició amino o carboxil (terminal) s’eliminen després d’haver fet la seva funció. Si es troben en posicions intermèdies la seqüència senyal formarà part de la proteïna madura; no s’eliminarà. Alguns cops també han de passar més d’un orgànul (perquè han d’anar a lloc diferents abans d’arribar al seu). Les seq. senyals no tenen per què ser úniques.
- Com s’ha demostrat que són les seqüencies senyal les que determinen la posició? Per experiments de manipulació genètica: traiem la seq. senyal d’una proteïna que s’ha de transportar i observarem que aquesta no surt del citosol.
8.2. TRANSPORT DE PROTEÏNES A TRAVÉS DE MEMBRANES
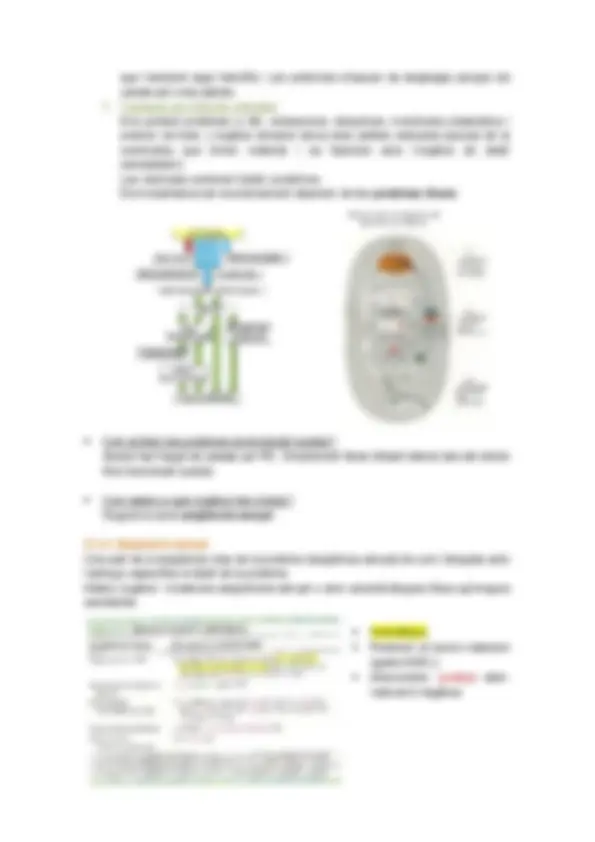
8.2.1. Entrada de proteïnes al nucli Els porus nuclears estan formats per unes estructures proteiques molt complexes que formen un canal aquós i funcionen com un diafragma. Tot i que els porus estiguin tancats, poden entrar molècules per difusió lliure (sempre i quan tinguin un ∅ inferior a 9nm). Totes les proteïnes que s’importen a dins del nucli tenen ∅ superior a 9 nm (histones, DNApolimerasa, etc...) per tant, han de fer obrir els porus nuclears per tal d’entrar dins del nucli, mitjançant la hidròlisi de GTP. Les proteïnes que s’importen a dins del nucli tenen una seqüència senyal NLS (senyal de localització nuclear). Els aa que codifiquen aquesta seqüencia senyal estan polaritzats (+).
Hi ha una proteïna anomenada importina citoplasmàtica que quan s’uneix a les proteïnes que tenen NLS formen un complexe proteïc que fa que s’obrin els porus nuclears. D’aquesta manera la proteïna que tingui una funció dins del nucli hi podrà entrar. Un cop la importina ha fet la seva funció, es desfà el complexe. La importina es recicla (s’expulsa al citosol mitjançant energia) i la proteïna+NLS fa la seva funció.
Î El senyal NLS és reconegut per la importina. Aquesta reconeix la proteïna a importar i van al forat del porus. Les fibrilles actuen com a rails.
Entra tot el complex i la importina es desenganxa (gastant GTP) i surt per importar un altre proteïna.
8.2.3. Importació de proteïnes a RE El RE serveix per portar proteïnes a altres llocs també. El transport de proteïnes és mitjançant vesícules a altres orgànuls (excepte al embolcall nuclear). Les proteïnes quan es sintetitzen al RE, tenen dues opcions:
-Es poden quedar integrades a la membrana Marcarà el lloc on aniran. O de membrana a membrana o de ca- -Cauen dins la cavitat (seran solubles) -vitat en cavitat.
L’inici de les proteïnes sempre és per ribosomes lliures al citosol. Uns missatgers es quedaran lliures o es quedaran a la membrana del reticle. Els ribosomes són sempre els mateixos. El que marca la diferència és la pròpia proteïna que s’està sintetitzant. Aquesta proteïna té una senyal: senyal d’importació al reticle (SRP). Les que el tinguin aniran a la membrana. Les que no, quedaran integrades en el citosol i no aniran a cap dels orgànuls.
Les proteïnes que portin el senyal aniran, quan comencin a ser formades, al reticle. Després, si han d’anar a altres llocs, tindran 2ons i 3ers senyals. Un cop el senyal hagi fet la funció, s’eliminarà, sempre que tingui la posició aminoterminal. Si tingués posició central (pocs casos) no s’eliminaria.
(1) La proteïna para de créixer, perquè el ribosoma no pot continuar ja que la seq. senyal s’ho impedeix. (2) SRP s’enganxa a la seq. senyal i condueix el complexe cap a la membrana del RE gràcies al receptor del SRP. (3) Un cop el complexe queda enganxat a la membrana, la proteïna retorna a créixer i passa per la proteïna translocadora cap al interior (cavitat). El SRP i el seu receptor es desenganxen i es reciclen. (4) El senyal es queda retingut a la membrana, la proteïna continua creixent cap a l’interior.
(^2 ) 4
5a 5b
1
(5) a) El ribosoma acaba de fer la seva funció. Quan la proteïna acaba de créixer, s’activa una senyal que avisa a la proteasa de que talli la proteïna. Aquesta queda lliure dintre de la cavitat i la seq. senyal es degrada. b) En la seqüència d’aa de la proteïna, hi ha unes “regions d’stop-transfer”. Quan la proteïna estigui creixent dintre de la cavitat, aquesta regió aturarà el procés, i l’invertirà, fent que la proteïna creixi cap a la banda citosòlica, i aquesta regió quedarà anclada en la membrana. Un cop hagi acabat, la senyal de la peptidasa tallarà la seqüència senyal. Quan una proteïna ha de travessar varies vegades la membrana, tindrà intercalades diverses seqüències d’stop/start-transfer.
8.3. BASES DE TRANSPORT VESICULAR
En molts casos, RE no és el destí final. El transport de compartiment a compartiment es fa per vesícules.
1- Ruta secretora biosintètica: comença al RE i passen cap a AG i d’aquí a membrana plasmàtica i exterior cel·lular. També pot ser que de AG vagi a endosomes i lisosomes. 2-Ruta endocítica: comença a membrana plasmàtica i exterior cel·lular i va a endosomes i d’aquí a lisosomes. 3-Ruta de recuperació: per a recuperar el que es propi de cada compartiment (normalment receptors). Com el que va d’endosomes primerases a membrana plasmàtica. També endosomes tardats a AG i d’aquí a RE.
Perquè tot aquest sistema funcioni cada vesícula porti les macromolècules adequades. S’aconsegueix amb proteïnes de recobriment i adaptadores , que tenen funció de seleccionar el que li convé a l’orgànul al qual va. També cal que la vesícula fusioni amb l’orgànul adequat; s’aconsegueix amb les Snare, que són proteïnes que es troben a la membrana.
8.3.2. Fusió de vesícules amb l’orgànul adequat Tipus Snare: V-Snare Æ a la mb de la vesícula (orgànul donador) T-Snare Æ a la mb de l’orgànul (orgànul diana)
Les v-Snare només interaccionen específica- ment.
Toxina botulisme: és mortal. Si algun menjar (com ara sardines) no està en bon estat, hi ha uns bacteris que generen toxines i generen gasos i per això les llaunes es bomben i fan sorollet al obrir-la. Aquesta toxina talla v-Snare i t-Snare (les vesícules no sabrien on fusionar-se). Al cos humà es produeix una petita paràlisi i es produeixen arrugues.
8.4. RUTA BIOSINTÈTICA SECRETORA
Aquesta és la ruta que fan les proteïnes i lípids per anar a diferents llocs.
8.4.1. RE: síntesi de proteïnes i lípids Síntesi de proteïnes Æ RE rugós Síntesi de lípids Æ RE llis
- Els lípids de gairebé totes les membranes es sintetitzen a les membranes del REL. Es fa bàsicament ensamblant “peces prefabricades”. Només es fa en la monocapa citosòlica perquè està mediada per uns enzims els centres actius dels quals es troben en aquesta capa.
Unes proteïnes agafen les cadenes hidrocarbonades i les porten a la monocapa. Aquesta monocapa creixerà més i, per equilibrar-les, intervindran les flipases per transportar lípids d’una membrana a l’altra. En un principi, aquestes flipases són inespecífiques, per tant, passaran qualsevol tipus de lípids (la composició de lípids estarà equilibrada). Més endavant, les flipases (al AG) seran específiques: passaran els fosfolípids determinats, i el que faran serà donar asimetria a la membrana lipídica.
8.4.2. RE: glucosilació de proteïnes Glucosilació : enganxar oligosacàrids a proteïnes. Hi ha dos orgànuls que ho fan: RE i AG. Les glucosilacions sempre es fan en la banda no citosòlica, d’orgànul a orgànul.
- Sempre es lliguen en el mateix aminoàcid [Asn]
- S’afegeixen en blocs (N-lligats) Formen 14 aminoàcids en total. Les proteïnes sempre porten els mateixos oligosacàrids (el sucre és el mateix). La diferenciació serà més endavant.
8.4.3. RE: control de qualitat Només es poden empaquetar en vesícules les proteïnes ben plegades, sinó les enviarien per proteïnes translocadores (chaperones) al citosol que degradaran a proteosomes. Les vesícules que surten es fusionen les unes amb les altres i formen un compartiment (Cluster) que fusionarà amb el AG.
8.4.4. Arribada i pas per AG
El cluster fusiona amb els primers compartiments de AG i aquest rep tot el que portaven. Una de les funcions de AG és reconèixer què prové del RE i retornar-li (mitjançant túbuls i vesícules)
- Les que siguin residents de la membrana del reticle tenen una senyal: KKXX
- Les residents de cavitat del reticle tenen una altra: KDEL (mai podrà anar a membrana plasmàtica). Seran reconegudes mitjançant receptors (per KDEL) i Cop I (per KKXX) El complexe Golgi té diferents regions:
Per exemple, les glàndules mamàries s’activen quan hi ha un part.
Es reconeixen perquè formen agregats a la CGN i s’empaqueten a vesícules de secreció. Una part de les cèl·lules, amb el pH àcid, formen senyals d’agregació. Aquestes parts amb el pH àcid fan que es formin els agregats. Fan créixer la membrana plasmàtica. En aquest tipus de secreció, el creixement es fa més ràpidament.
També hi ha la via de recuperació de membrana. Amb la endocitosi es torna a reduir.
8.4.7. Transport de proteïnes a lisosomes Sempre haurem de passar per endosomes. Es fa transport d’enzims hidrolítics (actúen en medi àcid). Aporten a l’interior dels orgànuls i a la seva membrana.
- Com reconeixen les trans Golgi les han d’anar a endosomes i lisosomes? Gràcies als enzims que estan a la cavitat de lisosomes: enzims lisosomals. Són glucoproteïnes, sintetitzades a RE. Arriben a Golgi i se les senyala com a proteïnes lisosomals fosforialnt les manoses. Aquest procés ens indica que és un enzim lisosomal. Avança des de la xarxa cis fins la trans, on les manoses fosforilades són reconegudes per receptors transmembranals, s’empaqueten i van cap a endosomes i lisosomes.
Tots els enzims lisosomals porten les manoses fosforilades. És un senyal que desvia la proteïna cap a lisosomes. El pH cada vegada és més àcid. El receptor es desenganxa de l’enzim. El receptor buit retorna a TGN. L’enzim lisosomal (que es troba a endosomes tardants) es desfosforila i arriba a lisosomes per maduració. Els diferents enzims lisosomals porten les manoses fosforilades. Qui fosforila les manoses és l’enzim fosfotransferasa. Aquest enzim es troba a la CGN. Reconeix la zona comú dels enzims lisosomals i s’hi enganxen, enganxant-se a la vegada un fosfat a les manoses.
Qualsevol proteïna sintetitzada a RE anirà a AG i d’aquí s’empaquetarà a vesícules de secreció per fer secreció constitutiva. Sempre serà així, a no ser que:
- La retinguem a RE (les KDEL)
- La retinguem a Golgi (senyals desconeguts)
- La desviem a fer secreció regulada (senyals d’agregació)
- Les desviem per anar a lisosomes (manoses fosforilades)
8.5. RUTES D’ENDOCITOSI
Tenim 3 mecanismes:
- Fagocitosi : entrada de partícules. És especialitzada.
- Pinocitosi : vesícules de pinocitosi
- Endocitosi per receptors: vesícules d’endocitosi mitjançant receptors La pinocitosi i l’endocitosi consisteixen en l’entrada de macromolècules, i la fan totes les cèl·lules. També es fan alhora a la mateixa vesícula.
8.5.1. Fagocitosi La fagocitosi només la fan algunes cèl·lules. Per exemple els neutròfils, que fagociten bacteris. Els macròfags també són cèl·lules de teixits que fagociten bacteris i restes cel·lulars. No és un mecanisme per adquirir aliments en animals (en organismes unicel·lulars la fagocitosi serveix per nodrir-se), sinó de defensa i renovació de cèl·lules.
- Què passa si el procés no funciona bé? Patiran hipercolesterolèmia familiar, que és una característica genètica relacionada amb el colesterol i es manifesta en grans i joves. Tenen infart de miocardi. Hereten mutacions al gen que codifica pel receptor de LDL, que queda fora. El colesterol queda a la sang, fa plaques arterioscleròtiques a venes i artèries i aprima el pas de la sang. Dificulta el pas de sang i provoca lesions.
8.5.4. Endosomes Endosomes: compartiment on va material de pinocitosi i fagocitosi per receptors. El pH és una mica àcid (6). Hi ha dos tipus:
- Primerencs Æ més a prop de la membrana plasmàtica
- Tardans Æ més propers al nucli
Primerencs: compartiments amb el que fusionen vesícules de pinocitosi i fagocitosi mediada per receptors.
- Distribueixen material, i careixen d’enzims hidrolítics. Surten vesícules que tornaran a la membrana plasmàtica (és un mecanisme de reciclatge i el fan servir, per exemple, receptors buits).
- Tot el que s’hagi de degradar anirà a primerencs, després a tardans i després a lisosomes.
- Via de vesícules que aniran a fer transcitosi. Es fa endocitosi per una banda i exocitosi per l’altra. No passa per lisosomes, perquè no s’ha de degradar, sinó transportar.
Per exemple: cèl·lules de l’epiteli intestinal, perquè els medis per on passen són diferents. També les inmunoglobulines arriben als nadons amb aquest mecanisme: reben els anticossos per la llet materna.
- Reserva de proteïnes de membrana. Per exemple, tenim pocs transportados de glucosa (glucosa permeases), perquè la [glu] és baixa. Les glucoses permeases estan amagades a endosomes primerencs. Quan la [glu] puja, la insulina s’enganxa amb receptors d’insulina i fa que els endosomes fusionin amb la membrana plasmàtica i haurà més transportadors i s’importarà més glucosa.
Els enzims de la TGN van a endosomes tardans.
Tardans: és on es troben allò que han de degradar amb els enzims hidrolítics. Els processos de degradació són poc actius perquè el pH no és gaire àcid (6) i els enzims no funcionen a ple rendiment, però tot i així, comença la degradació. Els receptors de manoses fosforilades es retornen a TGN. Tot el complex junt que
s’hagi de degradar passarà a lisosomes. Els endosomes tardans, a mida que van reciclant i es van acidificant es converteixen a lisosomes (és una maduració).
8.5.5. Lisosomes El lisosoma és l’orgànul on es fa la major part de digestió de material d’endocitosi, és a dir, on es degrada. Contenen enzims anomenats hidrolases àcides perquè funcionen bé amb pH 5 aproximadament. Aquests enzims poden ser nucleases, proteases... cada enzim està especialitzat en degradar un tipus determinat de complex. Per obtenir pH 5 hi ha les bombes de protons (H+) a les membranes dels lisosomes. Amb hidròlisi d’ATP porten protons a lisosomes i els acidifiquen. També trobem altres proteïnes de membranes, com ara de transport de subproductes degradats, com ara d’aa, monosacàrids, colesterol, fosfolípids... Tant lípids com proteïnes (glucolípids i glucoproteïnes) que constitueixen les membranes tenen molts glúcids enganxats (sempre a la banda no citosòlica). Fan funció de protecció, perquè els enzims de dins es “mengen” proteïnes i lípids de la membrana i s’ha de reparar contínuament. Si estan molt glucosilats, els enzims triguen més en degradar. Hi ha 3 vies d’obtenció de material a degradar:
1) Endocitosi: endosomes tardans Æ lisosomes 2) Fagocitosi: lisosomes (només a neutròfils o macròfags, que poden fagocitar). El fagosoma fusiona la seva membrana amb la del lisosoma. Aleshores, la partícula fagocitada es degradarà. 3) Autofàgia: lisosomes (a totes les cèl·lules). Es fagociten parts de la pròpia cèl·lula. Es forma una vesícula anomenada autofagosoma, que també fusionarà la seva membrana. Serveix per degradar parts velles de la pròpia cèl·lula (renovació). A vegades, les secretores fan massa vesícules secretores i amb l’autofàgia eliminen l’excés. També fan funció de nutrició en casos extrems degradant parts de la pròpia cèl·lula per produir ATP.