¡Descarga Conceptos previos y más Apuntes en PDF de Biología solo en Docsity!
Review
The mitochondrial genome: structure, transcription, translation and
replication
Jan-Willem Taanman *
Department of Clinical Neurosciences, Royal Free Hospital School of Medicine, University of London, Rowland Hill Street, London NW3 2PF, UK Received 12 May 1998; received in revised form 9 July 1998; accepted 21 July 1998
Abstract
Mitochondria play a central role in cellular energy provision. The organelles contain their own genome with a modified genetic code. The mammalian mitochondrial genome is transmitted exclusively through the female germ line. The human mitochondrial DNA (mtDNA) is a double-stranded, circular molecule of 16 569 bp and contains 37 genes coding for two rRNAs, 22 tRNAs and 13 polypeptides. The mtDNA-encoded polypeptides are all subunits of enzyme complexes of the oxidative phosphorylation system. Mitochondria are not self-supporting entities but rely heavily for their functions on imported nuclear gene products. The basic mechanisms of mitochondrial gene expression have been solved. Cis-acting mtDNA sequences have been characterised by sequence comparisons, mapping studies and mutation analysis both in vitro and in patients harbouring mtDNA mutations. Characterisation of trans-acting factors has proven more difficult but several key enzymes involved in mtDNA replication, transcription and protein synthesis have now been biochemically identified and some have been cloned. These studies revealed that, although some factors may have an additional function elsewhere in the cell, most are unique to mitochondria. It is expected that cell cultures of patients with mitochondrial diseases will increasingly be used to address fundamental questions about mtDNA expression. fl 1999 Elsevier Science B.V. All rights reserved.
Keywords: Mitochondrial DNA; Replication; Transcription; Translation; Protein synthesis; Mitochondrial encephalomyopathy; (Human)
Contents
- Introduction.......................................................... 104
- Structure of mtDNA.................................................... 105
0005-2728 / 99 / $ ^ see front matter fl 1999 Elsevier Science B.V. All rights reserved. PII: S 0 0 0 5 - 2 7 2 8 ( 9 8 ) 0 0 1 6 1 - 3
Abbreviations: CSB, conserved sequence block; EST, expressed sequence tags; HMG, high mobility group; HSP, H-strand promoter; H-strand, heavy strand; IT, initiation of transcription site; LSP, L-strand promoter; L-strand, light strand; mtDNA, mitochondrial DNA; mtIF, mitochondrial initiation factor; mtEF, mitochondrial elongation factor; mtRNase P, mitochondrial ribonuclease P; mtSSB, mitochondrial single-stranded binding protein; mtTERM, mitochondrial transcription terminator; mtTFA, mitochondrial transcription factor A; OH , origin of H-strand replication; OL , origin of L-strand replication; RNase MRP, mitochondrial RNA processing endo- ribonuclease; TAS, termination-associated sequence
Biochimica et Biophysica Acta 1410 (1999) 103^
- Transcription of mtDNA................................................. 106 3.1. Initiation of transcription............................................. 106 3.2. Elongation and termination of transcription................................ 108 3.3. Processing of primary transcripts........................................ 109
- Mitochondrial protein synthesis............................................ 110 4.1. Mitochondrial ribosomes.............................................. 110 4.2. Initiation and elongation of translation................................... 111 4.3. The e°ect of tRNA point mutations on mtDNA expression.................... 112
- Replication of mtDNA.................................................. 113 5.1. Basic mechanism of mammalian mtDNA replication......................... 113 5.2. Initiation of H-strand synthesis......................................... 113 5.3. Initiation of L-strand synthesis......................................... 115 5.4. Trans-acting factors involved in elongation and maturation of progeny strands...... 115 5.5. A role for mtTFA in mtDNA maintenance................................ 116
- Prospects............................................................. 117
Acknowledgements......................................................... 119
References............................................................... 119
- Introduction
Mitochondria are the energy-transducing organ- elles of eukaryotic cells in which fuels to drive cellu- lar metabolism are converted into ATP through the process of oxidative phosphorylation. Mitochondria have a double membrane. The outer membrane sep- arates the mitochondrion from the cytosol. The inner membrane is invaginated to form the cristae which protrude into and de¢ne the matrix of the organelle. The ¢ve enzyme complexes of the oxidative phos- phorylation system [1] are embedded in the mito- chondrial inner membrane. Mitochondria contain their own genome, the mitochondrial DNA (mtDNA), which is located in the mitochondrial ma- trix. In mammalian cells, each organelle generally contains several identical copies of mtDNA [2^5]. Mitochondria are thought to have originated from incorporated K-purple bacteria [6]. During its evolu- tion into the present-day powerhouses of the eukary- otic cell, the endosymbiont transferred many of its essential genes to the nuclear chromosomes. Never- theless, the mitochondrion still carries hallmarks of its bacterial ancestor. For instance, mitochondria use an N-formylmethionyl-tRNA (fMet-tRNA) as initia- tor of protein synthesis [7,8]. For over 20 years, it has been recognised that, in
mammals, mtDNA is only transmitted through the female germ line [9^12]. In mammalian sperm cells, the copy number of mtDNA is low (50^75 [13]), whereas in mammalian oocytes the copy number is extremely high (v 105 [2,14]). Therefore, the mater- nal inheritance of mtDNA observed in early studies could simply have been the result of dilution of the paternal contribution beyond the detection limit of the restriction enzyme analysis on which these studies relied. In a more recent investigation, in which a more sensitive, PCR-based technique was used, low levels of paternal mtDNA were detected in interspe- ci¢c hybrids of the mice species Mus musculus and Mus spretus throughout development from pronu- cleus stage to neonates [15]. In intraspeci¢c o°spring of M. musculus, however, paternal mtDNA was only detected in the early pronucleus stage [15]. In the majority of mammals, including humans, sperm mi- tochondria are transferred to the oocyte during fer- tilisation [16], but detailed morphological studies in rodents [15,17] and cows [18] have indicated that sperm-derived mitochondria are lost early in embryo- genesis. The mechanism underlying the elimination of sperm-derived mitochondria is unknown but the leakage of paternal mtDNA in progenies of interspe- ci¢c mice crosses suggest that this process is species speci¢c. Although it is not clear to which extent the
mechanism, allowing translation of all codons with less than the 32 tRNA species required according to Crick's wobble hypothesis. This reduction in the number of tRNA species is achieved by the use of a single tRNA with U in the ¢rst anticodon (wobble) position to recognise all codons of a four-codon fam- ily [29,30]. Fungal mitochondria use a modi¢ed U in the wobble position to read two codon families with a purine in the third position of the codon [31,32]. This mechanism prevents misreading of two codon families with a pyrimidine in the third position and is considered to be conserved in vertebrate mitochon- dria. This would imply that 24 tRNA species are required to decode mtDNA, however, as mentioned above, in vertebrates AGR codons indicate a stop and the corresponding tRNA gene is absent. Fur- thermore, a single tRNA species, with a modi¢ed C in the ¢rst anticodon position followed by AU, func- tions for initiation (tRNA fMet^ ) as well as elongation (tRNA Met^ ) and pairs with codon AUA in addition to AUG [28,33]. Hence, the 22 tRNA species encoded by human mtDNA are su§cient to translate all 13 mitochondrial protein genes. In vertebrate cells that are metabolically active, a large proportion of the mtDNA duplexes contain a short three-stranded structure, called the displace- ment loop or D-loop [23], in which a short nucleic acid strand, complementary to the L-strand, displa- ces the H-strand. The D-loop region is bounded by the genes for tRNAPhe^ and tRNAPro^ [34] and has evolved as the major control site for mtDNA expres- sion, containing the leading-strand origin of replica- tion and the major promoters for transcription (Fig. 1). Mitochondria are not self-supporting entities in the cell. Replication and transcription depend upon trans-acting nuclear-encoded factors. Mitochondrial tRNAs are charged by imported aminoacyl-tRNA synthases and, in vertebrates, all mitochondrial ribo- somal proteins are coded and synthesised outside the organelle. Enzymes of the various catabolytic path- ways located in the mitochondria, as well as the com- ponents of the mitochondrial import machinery, are encoded by nuclear DNA. Even the enzyme com- plexes of the oxidative phosphorylation system are of hybrid genetic origin. All nuclear-encoded poly- peptides destined for mitochondria are synthesised on cytosolic ribosomes, usually with a cleavable, N-
terminal presequence for mitochondrial targeting and are subsequently imported into mitochondria (re- viewed in [35]).
- Transcription of mtDNA
3.1. Initiation of transcription
The basic mechanism of mitochondrial transcrip- tion has been solved in representative species of sev- eral phylogenetic groups (reviewed in [36^38]). Hu- man mtDNA transcription initiation sites and promoter regions have been determined using a vari- ety of techniques, including 5P-end mapping of pri- mary mitochondrial transcripts by S1 nuclease pro- tection experiments [39^41] and deletion [42^44], site- directed mutagenesis [45] and linker substitution analyses [46] of cloned mtDNA fragments in run- o° transcription assays. All available data are consistent with the conclusion that there are two major transcription initiation sites in the D-loop (ITH1 and ITL ) situated within 150 bp of one another (Fig. 2). A promoter element with a 15-bp consensus sequence motif, 5P-CANACC(G)CC(A)AAAGAYA, surrounds the transcription initiation sites (under- lined) and is critical for transcription [43,45]. H-strand transcription starts at nucleotide position 561 (ITH1 ; numbering according to [21]) located within the H-strand promoter (HSP) and immedi- ately adjacent to the tRNAPhe^ gene, whereas the L-strand transcription starts at nucleotide position 407 (ITL ) located within the L-strand promoter (LSP). Additional upstream enhancer elements are required for optimal transcription (Fig. 2). These el- ements, which were later shown to include binding sites for a transcription factor (mtTFA, see below), exhibit sequence similarity but only if one element is inverted relative to the other [43,45,47]. Despite the close proximity of the HSP and LSP, the initial in vitro transcription studies demonstrated that these elements are functionally independent [42,43,45,46]. This functional autonomy was later corroborated in patients with progressive external ophthalmoplegia, harbouring a large-scale deletion, including the HSP, in a sub-population of their mtDNA molecules. In situ hybridisation experiments revealed focal accumulations of deleted mtDNA
and L-strand transcripts with concomitant depletion of H-strand transcripts in muscle ¢bres of the pa- tients [48,49]. These results con¢rm the functional independence of the transcriptional promoters in vivo. A second putative initiation site for H-strand tran- scription is located around nucleotide position 638 (ITH2 ) in the tRNAPhe^ gene, immediately adjacent to the gene for 12S rRNA (Fig. 2). Its promoter region only shows limited similarity with the 15-bp consensus sequence and this site is used less fre- quently than IT (^) H1 for transcription of the H-strand [39^41,43]. Although there is a fairly detailed picture of the cis-acting elements involved in mtDNA transcrip- tion, knowledge of the trans-acting nuclear-encoded factors is still incomplete. Biophysical fractionation of human mitochondrial transcription extracts have revealed the requirement of at least two trans-acting proteins: a relatively non-selective core RNA po- lymerase and a dissociable transcription factor which confers promoter selectivity on the polymerase [47,50]. The core enzyme is expected to interact with the HSP and LSP as mutations in these do-
mains obliterate even the lowest level of transcription [46]. Mitochondrial RNA polymerases have not been puri¢ed to homogeneity. Nevertheless, human cDNA specifying mitochondrial RNA polymerase was re- cently identi¢ed by screening of an expressed se- quence tags (EST) database with the yeast sequence [51]. Interestingly, the C-terminal half of the pre- dicted polypeptide shares signi¢cant amino acid se- quence identity with the single subunit RNA polym- erases of T3, T7 and SP6 bacteriophages. The human dissociable transcription factor acting in concert with the core mitochondrial RNA polym- erase has been puri¢ed [52,53], its cDNA has been cloned and sequenced [54] and its gene has been characterised [55^57]. The factor, now termed mtTFA [58], is an abundant 25-kDa mitochondrial protein and is largely comprised of two high mobility group (HMG) domains separated by a 27-amino acid residue linker and followed by a 25-amino acid res- idue basic C-terminal tail. HMG domains are con- sidered to be involved in DNA binding and are found in a rather diverse family of proteins whose members have been implicated in processes such as transcription enhancement and chromatin packaging
Fig. 2. Schematic representation of the initiation of transcription and replication of human mtDNA. The genes encoding 12S rRNA, tRNA Phe^ and tRNAPro^ are indicated with boxes on the H- and L-strands. Transcription initiation sites and direction of synthesis are indicated by bent arrows, dotted lines represent synthesised RNA. In the D-loop region (the 1118-bp sequence between nucleotides 577 and 16028), two major transcription initiation sites are present. Transcription initiation site IT (^) H1 , encompassed by the H-strand promoter (HSP), directs the transcription of the H-strand, whereas transcription initiation site IT (^) L , encompassed by the L-strand pro- moter (LSP), directs the transcription of the L-strand. A second, minor transcription initiation site (ITH2 ) for H-strand transcription is located in the gene for tRNA Phe^ near the boundary with the 12S rRNA gene. Enhancer elements upstream of the HSP and LSP that are known to bind the mitochondrial transcription factor mtTFA are indicated. A short (RNA) transcript originating at IT (^) L serves as a primer for replication of the (leading) H-strand. Transitions from RNA to DNA occur within the dashed line, in the re- gion around the conserved sequence blocks (CSBs) I, II and III. O (^) H is the origin of H-strand synthesis. Short DNA strands that are part of the triplex D-loop structure terminate near the termination-associated sequence (TAS).
en further credence to the model [62]. Nevertheless, it is di§cult to imagine how two initiation events tak- ing place less than 100 bp apart can determine the fate of RNA synthesis at the distal end of the 16S rRNA gene, more than 2500 nucleotides down- stream. In addition to the dual H-strand transcription model, the di°erence in synthesis rate of rRNA and mRNA has been explained by an attenuation event at the border of the 16S rRNA and tRNALeuÖUURÜ^ genes (Fig. 1). The ¢rst indication of early termination of the polycistronic H-strand tran- script came from structural analysis of the 3P-ends of 16S rRNA molecules which revealed that a large proportion of the molecules have ragged 3P-termini that are coded for by the immediately adjacent gene for tRNALeuÖUURÜ^ [78,79]. This suggests that mature 16S rRNA species are generated by imprecise tran- script termination at the tRNALeuÖUURÜ^ gene as well as by accurate endonucleolytic cleavage of the longer precursor RNA. Later, a crude protein fraction was isolated from mitochondrial lysates of HeLa cells which, in DNase I footprinting studies, protected the region immediately downstream of the mtDNA region corresponding to in vivo produced 3P-ends of 16S rRNA molecules and promoted speci¢c termina- tion of transcription [80]. The footprint encompasses a conserved tridecamer sequence block within the tRNALeuÖUURÜ^ gene (nucleotide positions 3237^ of the human mtDNA; Fig. 1), which has been shown by in vitro deletion mutagenesis experiments to be essential and su§cient for directing termination [81]. The factor mediating attenuation of transcription has been termed mTERF [80] or mtTERM [82] and is known to induce bending of the DNA helix [83]. In vitro transcription studies have shown that mtTERM bound to its mtDNA target site functions bidirec- tionally and shows an even greater e§ciency of ter- mination in the reverse orientation relative to the promoter site [84]. Thus, in addition to an attenua- tion function for H-strand transcription, mtTERM may halt L-strand transcription at a site where no L-strand encoded genes are found downstream (see Fig. 1). Bound mtTERM probably stops elongation of transcription by constituting a physical barrier, rather than by a speci¢c interaction with the mito- chondrial RNA polymerase, because mtTERM also
mediates termination of transcription by heterolo- gous RNA polymerases [83]. Both Clayton's and Attardi's groups have shown that a polypeptide fraction of a mitochondrial lysate containing polypeptides with a molecular weight of around 34 kDa has the ability to produce the speci¢c DNase I footprint at the termination site associated with mtTERM function [82,85]. The cDNA of the predominant polypeptide from this fraction was re- cently cloned and sequenced [86]. The polypeptide contains two widely separated basic regions and three leucine zipper motifs which were shown to be necessary for its speci¢c DNA-binding capacity [86]. The footprint produced by the recombinant protein was similar but not identical to that produced by the 34-kDa polypeptide fraction. The recombinant pro- tein was also shown to be unable to promote tran- scription termination in an in vitro system [86]. These observations suggest that an additional component of the 34-kDa polypeptide fraction is required for the termination activity. This apparent complexity of mtTERM is not surprising, given that it should be able to modulate its activity in response to the cellular demand for mitochondrial rRNAs, on the one hand, and for mitochondrial tRNAs and mRNAs on the other. Interestingly, a heteroplasmic A to G transition in the middle of the mtTERM binding site (A3243G) is frequently found in patients with the mitochondrial encephalomyopathy MELAS (mitochondrial myopa- thy, encephalopathy, lactic acidosis and stroke-like episodes) [87] and in patients with maternally inher- ited adult onset diabetes [88]. In vitro studies have shown that this mutation dramatically reduces the a§nity of mtTERM for its binding site and causes a defect in transcript termination [82,83,89]. In con- trast, steady-state transcript levels upstream and downstream of the termination site were not a°ected by the A3243G transition in cultured cells [89,90] or in tissues of patients [49,91,92]. Thus, it seems un- likely that the in vitro observed defective attenuation of mitochondrial transcription is of pathological sig- ni¢cance.
3.3. Processing of primary transcripts
Once the RNA polymerase passes the 16S rRNA/ tRNALeuÖUURÜ^ boundary, H-strand transcription ap-
pears to be straightforward. As no intron sequences are present in vertebrate mtDNA and intergenetic sequences are minimal, processing of the long poly- cistronic H- and L-strand messengers is thought to be a relatively simple process, requiring only a few enzymes. Genes for tRNAs £ank the two rRNA genes and nearly every protein gene (Fig. 1). This unique genetic organisation has led to the proposal that the secondary structure of the tRNA sequences provide the punctuation marks in the reading of the mtDNA information [27]. Precise endonucleolytic ex- cision of the tRNAs from the nascent transcripts will concomitantly yield correctly processed rRNAs and, in most cases, correctly processed mRNAs [27,93]. In those cases in which the mRNA termini cannot be accounted for by tRNA excision (e.g. the messenger for subunit I of cytochrome c oxidase [27,93]; Fig. 1), the processing enzyme possibly recognises a sec- ondary structure which shares critical features with the typical cloverleaf structures of tRNAs. Maturation of mitochondrial tRNAs involves three enzymatic activities which were recently identi- ¢ed by Rossmanith and colleagues [94] in an in vitro HeLa cell mitochondrial tRNA processing system. Their experiments showed that cleavage at the 5 P-end precedes that at the 3P-end. The endonuclease responsible for 3P-end cleavage has not been charac- terised. Cleavage at the 5P-end is performed by a mitochondrial RNase P (mtRNase P). The enzyme containing fractions prepared by Rossmanith and colleagues [94] cut mitochondrial tRNA precursors at the correct 5P-end, but, unlike the preparations by others (see, e.g. [95]), do not cleave tRNATyr^ pre- cursors of Escherichia coli correctly. This suggests that previous preparations were contaminated with a cytosolic isoform of RNase P which is apparently able to accurately process bacterial tRNA precur- sors. Yeast mtRNase P has been characterised in de- tail. The enzyme of Saccharomyces cerevisiae is com- posed of a nuclear-encoded protein and a mtDNA- encoded RNA species [96,97]. The RNA moiety of the ribonucleoprotein complex is AU-rich and forms the catalytic core of the enzyme. Comparison of mtRNase P RNA from di°erent yeast species has revealed a remarkable variation in size from 490 to 140 nucleotides [98]. Maturation of the excised tRNAs is completed by addition of the sequence CCA to their 3P-end cata-
lysed by ATP(CTP):tRNA nucleotidyltransferase [94]. Mitochondrial mRNAs are polyadenylated by a mitochondrial poly(A) polymerase during or imme- diately after cleavage [99,100], whereas the 3P-ends of the two rRNAs are post-transcriptionally modi¢ed by the addition of only short adenyl stretches [78]. Mitochondrial messengers do not carry upstream polyadenylation signals as found in nuclear messen- gers.
- Mitochondrial protein synthesis
4.1. Mitochondrial ribosomes
Early indications of the prokaryotic origin of mi- tochondria came from observations, now more than three decades ago, that the spectrum of antibiotics inhibiting mitochondrial protein synthesis resembles that of prokaryotic systems (reviewed in [101]). Sub- sequent research con¢rmed that nearly all constitu- ents of the mitochondrial translation machinery are distinct from their cytosolic counterparts. Unique features of mitochondrial protein synthesis were ¢rst studied at the molecular level in mitochondrial ribo- somes. The mitochondrial ribosomes, or mitoribo- somes, are located in the matrix of the organelle. Steady-state rRNA levels in rat hepatocytes [102] suggest that there are 6 100 mitoribosomes per mi- tochondrion. The physical and chemical properties of mitoribosomes di°er considerably from their cyto- solic as well as their bacterial counterparts. Mamma- lian mitoribosomes have an unusually low RNA con- tent and, consequently, a low sedimentation coe§cient of V55S [103^106]. The V39S and V28S ribosomal subunits contain respectively the 16S and 12S rRNA species encoded by the mtDNA [103,104]. A 5S rRNA species, typically present in ribosomes, appears to be absent in mammalian mi- toribosomes [103,104] but a 23-bp region at the 3 P-end of the human 16S rRNA exhibits a 68% se- quence identity to a portion of the Bacillus subtilis 5S rRNA. The structural similarity of the region and its mapping position have led to the suggestion that this piece represents a truncated 5S rRNA which remained part of the large rRNA component [107]. The low RNA content of mammalian mitoribo-
Three mitochondrial elongation factors, mtEF-Tu, mtEF-Ts and mtEF-G, have been puri¢ed from bo- vine liver [123,124]. The cDNAs for all three factors have been cloned and sequenced from mammalian sources [125^129] and the gene for human mtEF- Tu has been mapped to chromosome 16q11.2 [129]. The in vitro characterisation of the puri¢ed factors and the cDNA sequence information have revealed striking similarities with the corresponding prokary- otic factors. Consequently, elongation of the nascent mitochondrial polypeptide is assumed to proceed in a similar fashion as in E. coli [130]. Di°erent from E. coli EF-Tu and EF-Ts, mammalian mtEF-Tu and mtEF-Ts form a tightly associated complex that, un- like the bacterial complex, cannot readily be dissoci- ated by guanidine nucleotides alone [123]. However, it was recently demonstrated that the mtEF-Tu-Ts complex will dissociate in the presence of GTP and charged tRNAs [131].
4.3. The e°ect of tRNA point mutations on mtDNA expression
Pathogenic mutations in mitochondrial genes are generally heteroplasmic, i.e. wild-type and mutant mtDNA co-exist in the same cell. The wild-type mtDNA allows the mutant mtDNA with the other- wise lethal base change to persist. Maternally inher- ited, mitochondrial encephalomyopathies are often associated with point mutations in mitochondrial tRNA genes [132]. Muscle biopsies from patients
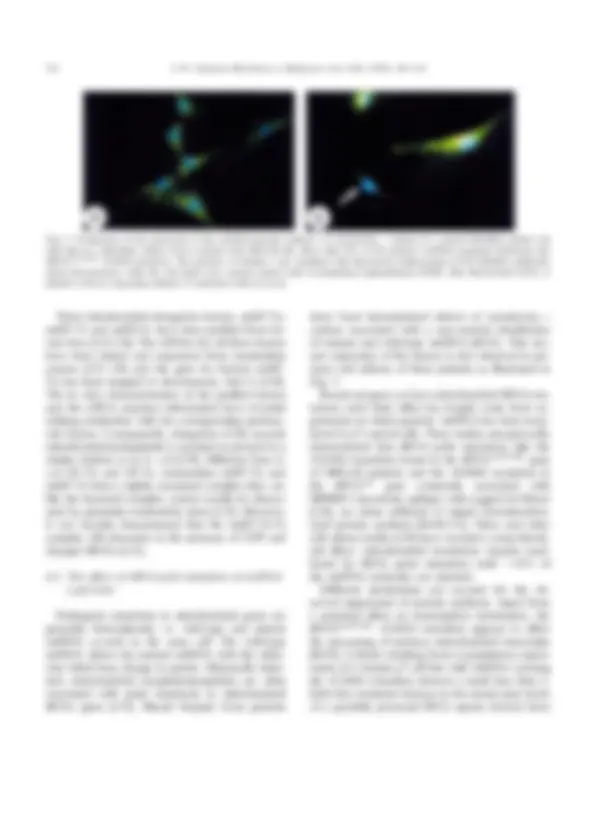
show focal histochemical defects of cytochrome c oxidase associated with a non-random distribution of mutant and wild-type mtDNA [49,91]. This mo- saic expression of the disease is also observed in pri- mary cell cultures of these patients as illustrated in Fig. 3. Recent progress on how mitochondrial tRNA mu- tations exert their e°ect has largely come from ex- periments in which patients' mtDNA has been trans- ferred to b^0 control cells. These studies unequivocally demonstrated that tRNA point mutations, like the A3243G transition found in the tRNALeuÖUURÜ^ gene of MELAS patients and the A8344G transition in the tRNALys^ gene commonly associated with MERRF (myoclonic epilepsy with ragged-red ¢bres) [134], are alone su§cient to impair intramitochon- drial protein synthesis [89,90,135]. These and other cell culture studies [136] have revealed a steep thresh- old e°ect: mitochondrial translation remains unaf- fected by tRNA point mutations until s 85% of the mtDNA molecules are mutated. Di°erent mechanisms can account for the ob- served impairment of protein synthesis. Apart from a potential e°ect on transcription termination, the tRNALeuÖUURÜ^ A3243G transition appears to a°ect the processing of primary mitochondrial transcripts [90,92]. Cybrids resulting from re-population experi- ments of a human b^0 cell line with mtDNA carrying the A3243G transition showed a small (less than 2- fold) but consistent increase in the steady-state levels of a partially processed RNA species derived from
Fig. 3. Comparison of the expression of the mtDNA-encoded subunit I of cytochrome c oxidase in a control ¢broblast culture (A) with that in a ¢broblast culture from a patient with MELAS (B). More than 95% of the patient's mtDNA molecules harboured the tRNA LeuÖUURÜ^ A3243G mutation. The presence of subunit I was visualised with £uorescein isothiocyanate (FITC)-labelled antibodies (green £uorescence), while the cell nuclei were counter-stained with 4,6-diamidino-2-phenylindole (DAPI; blue £uorescence) [133]. A patient's cell not expressing subunit I is indicated with an arrow.
transcription of the 16S rRNA+tRNA LeuÖUURÜ+ND genes [90,137], which are contiguous in the mtDNA (Fig. 1). Steady-state levels of mature 16S rRNA, tRNALeuÖUURÜ^ and ND1 mRNA are not a°ected by the mutation [90,137]. Although the steady-state level of the immature transcript is extremely low in the cybrids compared to the level of mature 16S rRNA, the level showed a strong inverse correlation with the rates of oxygen consumption of the cybrids (an indicator of mitochondrial oxidative phosphoryl- ation capacity [138,139]). These observations have led to the hypothesis [90,138] that the unprocessed transcript, which contains 16S rRNA, is incorpo- rated into ribosomes rendering them functionally de- ¢cient. If this results in stalling of the translation of polyribosomal mRNAs, then a small increase of un- processed transcript could interfere disproportionally with mitochondrial translation and explain the severe oxidative phosphorylation defects observed in the patients. Alternatively, point mutations in tRNA genes may induce a conformational change of the tRNA result- ing in a decreased stability of the molecule. A change in structure may also a°ect the identi¢cation by the cognate aminoacyl-tRNA synthase and lead to de- creased levels of aminoacylation or even mischarging of mutated tRNAs. In prokaryotes, the T 8 C region of the tRNA molecule is important for recognition and binding by EF-Tu and ribosomes, therefore, mu- tations in the T 8 C region of mitochondrial tRNAs are likely to a°ect their a§nity with mtEF-Tu and mitoribosomes. High resolution Northern (RNA) blot hybridisation experiments have indicated that these mechanisms are indeed likely to play a role in the aetiology of these diseases. In cell lines carrying either tRNALys^ A8344G [140,141], tRNALeuÖUURÜ C3256T [142], or tRNAAsn^ G5703A mutations [143], markedly reduced steady-state levels of the af- fected tRNA have been found, suggesting an in- creased susceptibility to nucleolytic digestion. More- over, a V40% reduction in aminoacylation of tRNALys^ has been demonstrated in cell lines with the tRNALys^ A8344G mutation [141]. In contrast, cell lines with the tRNALys^ C8356T mutation exhib- ited no signi¢cant decrease in tRNALys^ content [140] but this mutation, which is located in the T 8 C arm of the molecule, may interfere with the binding of mtEF-Tu or the mitoribosome [144].
- Replication of mtDNA
5.1. Basic mechanism of mammalian mtDNA replication
The fortuitously slow rate of mtDNA replication has facilitated the isolation and characterisation of in vivo replicative intermediates and has provided the now generally accepted model of the replication cycle of mammalian mtDNA. Early studies, which pre- dominantly relied on electron microscopic and cen- trifugal analysis of mtDNA molecules from cultured cells, indicated that mammalian mtDNA molecules replicated unidirectionally from two spatially and temporally distinct, strand-speci¢c origins [23]. The origin of H-strand replication (OH ) is located down- stream of the LSP in the D-loop region of the ge- nome, whereas the origin of L-strand replication (OL ) is at two-thirds of the genomic distance away from O (^) H with respect to the polarity of H-strand synthesis (Fig. 1). A round of replication begins at O (^) H with the synthesis of a daughter H-strand and continues along the parental L-strand to produce a full H-strand circle. Only after the replication fork has passed the second replication origin, O (^) L , is syn- thesis of the L-strand initiated which proceeds in a direction opposite to that of H-strand replication (reviewed in [145]).
5.2. Initiation of H-strand synthesis
Fine mapping of RNA and DNA species in the D- loop region of human and mouse mtDNA have sug- gested that short mitochondrial transcripts, originat- ing at IT (^) L , serve as primers for the initiation of syn- thesis of nascent H-strands (Fig. 2; [146,147]). Thus, replication of mammalian mtDNA appears to be in- timately linked with mitochondrial transcription. There are no known di°erences between the initia- tion of L-strand transcription and the initiation of RNA primer formation for mtDNA replication [36] and it is not clear which mechanism decides between transcript elongation or H-strand synthesis. Transi- tions from RNA to DNA synthesis take place at several distinct sites that collectively constitute OH in a region of three short, evolutionary conserved sequence blocks, named CSB I, II and III (Fig. 2; [34]).
ments and regulate the equilibrium between D-loop formation and H-strand replication.
5.3. Initiation of L-strand synthesis
The replication origin OL was ¢rst identi¢ed in mouse and later in human mtDNA by 5P-end map- ping of in vivo nascent L-strands [169,170]. The ori- gin is located in a non-coding region of V30 nucleo- tides and is £anked by ¢ve tRNA genes (Fig. 1). O (^) L is only activated when the parental H-strand is dis- placed as a single strand by the growing daughter H- strand. After strand displacement, O (^) L is thought to adopt a distinctive stem-loop structure [36]. In vitro run-o° replication studies of O (^) L have suggested that this con¢guration serves as the recognition structure for a mitochondrial DNA primase which provides a short RNA primer for L-strand synthesis [171,172]. However, the location of OL within a cluster of tRNA genes and the fact that the potential stem- loop structure might be absent in mtDNA of some vertebrate species [149] suggest that additional sec- ondary structures contribute to DNA primase recognition in vivo. RNA priming starts at the T-rich portion of the predicted OL -loop and the transition from RNA to DNA synthesis takes place at a speci¢c site near a critical GC-rich element at the base of the hairpin [171,172]. The mitochondrial DNA primase involved in priming of the L-strand synthesis has only been partly puri¢ed [173]. The physical properties of the mitochondrial enzyme are distinct from the major nuclear DNA primase. Mitochondrial DNA primase is associated with RNA. Treatment with ribonuclease A leads to rapid inactivation of primase activity and suggests a crucial role for the associated RNA. The predominant RNA moiety co-fractioning with primase activity is the nuclear gene product 5.8S rRNA [173]. However, as only crude fractions of mitochondrial DNA primase have been analysed, the co-fractionation of 5.8S rRNA might have been the result of an adventitious contamination.
5.4. Trans-acting factors involved in elongation and maturation of progeny strands
DNA polymerase Q is the only DNA polymerase present in mitochondria [174]. Disruption of the
DNA polymerase Q gene in S. cerevisiae has demon- strated that the enzyme is necessary for mtDNA syn- thesis but has no basic function outside the organelle [175]. DNA polymerase Q accounts for only a small fraction of the total cellular DNA polymerase activ- ity, yet its unique enzymatic properties, its resistance to aphidicolin and its sensitivity to dideoxynucleoside triphosphates make it relatively easy to identify the mitochondrial DNA polymerase [176]. The enzyme has been isolated from humans [177] and several other organisms (reviewed in [149]). DNA polymer- ase Q appears to be prone to proteolytic degradation and its subunit structure is still a matter of debate. Most recent data suggest that the enzyme is a heter- odimer in vertebrates comprised of a 125^140-kDa polypeptide and a 35^54-kDa polypeptide (see, e.g. [178]). In addition to its 5PC 3 P polymerase activity, DNA polymerase Q has an intrinsic 3PC 5 P exonu- clease activity which is highly mispair-speci¢c and ensures faithful replication of mtDNA [176]. In re- cent years, cDNA sequences specifying the major subunit have been identi¢ed from a number of verte- brate species, including human [179^181]. Sequence comparisons with the polypeptide from S. cerevisiae have shown that the major subunit is evolutionarily conserved and have allowed identi¢cation of both the polymerase and the exonuclease proof-reading domain. Recent kinetic studies with the recombi- nantly expressed, 140-kDa human subunit showed that the major subunit is a relatively poor polymer- ase compared to other DNA polymerases, suggesting that this core subunit may require accessory factors to increase the enzymatic rate [182]. Nevertheless, the kinetic parameters indicated that the major subunit could replicate the mitochondrial genome in a phys- iologically relevant time frame [182]. Replicative intermediates of mtDNA feature ex- tensive single-stranded DNA regions [145]. A mito- chondrial single-stranded binding protein (mtSSB) has been isolated and cloned from human [183] and several other species (reviewed in [149]). Mutation analysis in S. cerevisiae has demonstrated that mtSSB is required for maintenance of mtDNA [184]. Binding of mtSSB to the exposed single- stranded mtDNA is thought to maintain the integrity of these regions and to stimulate the overall rate of DNA synthesis by DNA polymerase Q in vivo (see, e.g. [185,186]). The mitochondrial protein is distinct
from the nuclear single-stranded binding protein but resembles the E. coli single-stranded binding protein in structure as well as in DNA-binding properties [187,188]. The protein interacts with single-stranded DNA as a homotetramer [187^189]. The crystal structure of human mtSSB suggests that single- stranded DNA wraps around the tetrameric complex through electropositive channels guided by £exible loops [189]. Several additional enzymatic activities are consid- ered to be essential for mtDNA replication but only a few supplementary enzymes of the mitochondrial replication machinery have been identi¢ed. Helicases catalyse the unwinding of duplex DNA by disrupting the hydrogen bonds that hold the two strands togeth- er to provide single-stranded templates for DNA pol- ymerases [190]. An ATP-dependent helicase has been identi¢ed and partly puri¢ed from mitochondria of bovine brain [191] and sea urchin eggs [192]. The enzyme shows biochemical similarities with Rep heli- case of E. coli and shares its 3PC 5 P polarity of un- winding with respect to the single-stranded portion of the partial duplex DNA. This direction would place the mitochondrial helicase on the template strand ahead of DNA polymerase Q during mtDNA replication. Type I topoisomerases catalyse the relaxation of supercoiled DNA, while type II topoisomerases are responsible for the introduction of supercoils. Topo- isomerases change the topology of DNA by transi- ently breaking the backbone bonds, using a mecha- nism involving either single strand (type I) or double strand (type II) cleavage [193]. Mitochondrially asso- ciated, type I topoisomerases have been identi¢ed from various sources, including human leukemia cells [194] and human platelets [195]. The mitochon- drial enzyme is immunologically related to its nuclear counterpart [196,197] and yeast mitochondrially as- sociated type I topoisomerase activity is abolished when the gene that encodes the nuclear type I top- oisomerase is disrupted [198]. Despite the apparent immunological and genetic relationship between the nuclear and mitochondrial type I topoisomerases, the enzymes can be distinguished from each another by a di°erential response to chemical reagents and inhib- itors, both in yeast and humans [195,197]. Mitochondrially associated type II topoisomerases have been identi¢ed in Dictyostelium discoideum
[199], rat liver [200] and human leukemia cells [194]. Further support for the existence of a distinct mitochondrial type II topoisomerase comes from ¢ndings with cell cultures grown in the presence of cipro£oxacin. This antibiotic inhibits prokaryotic type II topoisomerases much more than the eukary- otic nuclear enzyme. Treatment of cultured cells with cipro£oxacin causes double-stranded breaks in mtDNA and results in loss of the mitochondrial ge- nome [201]. These observations suggest the presence of a bacterial-like type II topoisomerase in mitochon- dria.
5.5. A role for mtTFA in mtDNA maintenance
In addition to its role in transcription and, conse- quently, primer formation for H-strand synthesis, mtTFA appears to have a function in maintenance of mtDNA. Mitochondria do not contain histones [202,203]. Yeast mtTFA [204] contains a tandem of HMG DNA-binding motifs, similar to mtTFA of vertebrates, but yeast mtTFA lacks the basic C-ter- minal region shown to be important for promoter selection in vertebrates [60]. The yeast homologue is not considered to play a key role in initiation of transcription [37] but is thought to have a histone- like function in coating the entire mitochondrial ge- nome [66,202]. Likewise, binding of vertebrate mtTFA is not restricted to the regions upstream of the HSP and LSP. The factor is inherently £exible in its recognition of DNA sequences [52,65,71] and a comparative in organello and in vitro footprinting study indicated that human mtTFA binds at regu- larly spaced intervals throughout a 500-bp region encompassing O (^) H and the two major promoters [61]. Binding is largely excluded from CSB II and III but mtTFA shows a strong interaction with CSB I [61,65]. As some of the intervals of mtTFA binding align with prominent 5P-ends of nascent H- strands, it has been suggested that the phased bind- ing of the protein functionally organises the D-loop and facilitates access of other trans-acting factors in- volved in the transition from RNA to DNA synthesis [37,61,65]. There exists a direct correlation between mtTFA and mtDNA steady-state levels. Heterozygous mtTFA knockout mice exhibit reduced mtDNA copy numbers and homozygous knockout embryos
post-mortem. No mtDNA mutations have been documented in these infants and pedigree analysis is consistent with autosomal inheritance of the trait. A nuclear genomic involvement has been con¢rmed in two families by mtDNA transfer techniques [218,133]. The progressive loss of mtDNA in tissues is also observed in cell cultures of some of the pa- tients [218,133]. In the depleting cultures, a fraction of the cells still contains apparently normal levels of mtDNA but mtDNA replication is more or less di- minished in all cells (Fig. 4). Taken together, these observations strongly suggest a replication arrest due to a de¢ciency of a trans-acting, nuclear-encoded fac- tor. Patients with mtDNA depletion display decreased levels of mtTFA in tissue [206,207]. However, the
fact that remnant mtDNA is still transcribed in cell cultures of the patients [133,219] suggests that the decrease in mtTFA levels is probably secondary to depletion of mtDNA. Other candidate genes, listed in Table 1, are currently being evaluated but, to date, no molecular defects underlying the mtDNA deple- tion have been reported. A de¢ciency of one of the key enzymes of mtDNA replication is likely to be lethal early in embryonic development [205]. The ap- parent tissue-speci¢c and neonatal expression of symptoms indicates a less drastic defect and points to the de¢ciency of a factor involved in regulation of mtDNA copy numbers. Regulatory factors are gen- erally present at very low cellular concentrations and only at certain stages of development, making it ex- ceedingly di§cult to characterise these factors by tra-
Fig. 4. Comparison of total mtDNA (A, B) and recently replicated mtDNA (C, D) levels in a control myoblasts culture (A, C) with that in a myoblast culture from a patient exhibiting mtDNA depletion (B, D). In (A) and (B), SYTO 17 (Molecular Probes) was used to visualise mtDNA; this cell-permeant, red £uorescent dye reveals the characteristic mitochondrial structures in the cell if mtDNA is present. In (C) and (D) replicating mtDNA was visualised by the incorporation of 5-bromo-2P-deoxyuridine (BrdU) and subsequent detection with an FITC-labelled antibody (green £uorescence). Nuclei were stained £uorescent blue with DAPI. Cells were either cul- tured for 1.5 h in medium containing 2 Wg/ml of DAPI, followed by culturing for 30 min in 62.5 nmol of SYTO 17, 2 Wg/ml of DAPI, 10 mM of HEPESWNaOH (pH 7.4), 150 mM of NaCl, 2 mM of CaCl 2 and 2 mM of MgCl 2 (A, B), or cells were cultured for 16 h in medium containing 15 WM of BrdU, 10 Wg/ml of aphidicolin followed by immunological detection of BrdU (Detection kit I, Boehringer Mannheim) and counter staining with DAPI (C, D). Arrows indicate a patient's cell lacking mtDNA or a patient's cell de- void of mtDNA replication.
ditional means. It may, however, be feasible to iden- tify the de¢cient factor in complementation studies of mtDNA depleting cell cultures with a human cDNA expression library. This approach is facilitated by the fact that the patient's cells, because of impairment of the oxidative phosphorylation complexes, become auxotrophic for uridine and pyruvate [218,133], which provides selectable markers to identify com- plementing cDNAs. One can anticipate that future progress in the ¢eld of replication, transcription and translation of hu- man mtDNA will increasingly come from detailed case studies as exempli¢ed above for mtDNA deple- tion. This research will not only enrich science but will also permit the development of diagnostic tools as well as new pathogenic insights to minimise the devastating consequences of a defective expression of the mitochondrial genome.
Acknowledgements
I thank Prof. A.H.V. Schapira and Dr J.C. Blake for helpful discussion. The review was written in con- juncture with studies supported by The Wellcome Trust, Grant 048410.
References
[1] Y. Hate¢, Annu. Rev. Biochem. 54 (1985) 1015^1069. [2] G.S. Michaels, W.W. Hauswirth, P.J. Laipis, Dev. Biol. 94 (1982) 246^251. [3] R.C. Shuster, A.J. Rubenstein, D.C. Wallace, Biochem. Bio- phys. Res. Commun. 155 (1988) 1360^1365. [4] E.D. Robin, R. Wong, J. Cell. Physiol. 136 (1988) 507^513. [5] R.J. Wiesner, J.C. Ru´ egg, I. Morano, Biochem. Biophys. Res. Commun. 183 (1992) 553^559. [6] M.W. Gray, Int. Rev. Cytol. 141 (1992) 233^357. [7] J.B. Galper, J.E. Darnell, Biochem. Biophys. Res. Commun. 34 (1969) 205^214. [8] J.L. Epler, L.R. Shugart, W.E. Barnett, Biochemistry 9 (1970) 3575^3579. [9] C.A. Hutchison, J.E. Newbold, S.S. Potter, M.H. Edgell, Nature 251 (1974) 536^538. [10] A.M. Kroon, W.M. Vos, H. Bakker, Biochim. Biophys. Acta 519 (1978) 269^273. [11] J.-I. Hayashi, H. Yonekawa, O. Gotoh, J. Watanabe, Y. Tagashira, Biochem. Biophys. Res. Commun. 83 (1978) 1032^1038.
[12] R.E. Giles, H. Blanc, H.M. Cann, D.C. Wallace, Proc. Natl. Acad. Sci. USA 77 (1980) 6715^6719. [13] N.B. Hecht, H. Liem, K.C. Kleene, R.J. Distel, S.M. Ho, Dev. Biol. 102 (1984) 452^461. [14] L. Piko®, L. Matsumoto, Dev. Biol. 49 (1976) 1^10. [15] H. Kaneda, J.-I. Hayashi, S. Takahama, C. Taya, K. Fischer Lindahl, H. Yonekawa, Proc. Natl. Acad. Sci. USA 92 (1995) 4542^4546. [16] F. Ankel-Simons, J.M. Cummins, Proc. Natl. Acad. Sci. USA 93 (1996) 13859^13863. [17] J. Hiraoka, Y. Hirao, Gamete Res. 19 (1988) 369^380. [18] P. Sutovsky, C.S. Navara, G. Schatten, Biol. Reprod. 55 (1996) 1195^1205. [19] G. Manfredi, D. Thyagarajan, L.C. Papadopoulou, F. Pal- lotti, E.A. Schon, Am. J. Hum. Genet. 61 (1997) 953^960. [20] http://megasun.bch.umontreal.ca/ [21] S. Anderson, A.T. Bankier, B.G. Barrell, M.H.L. De Bruijn, A.R. Coulson, J. Drouin, I.C. Eperon, D.P. Nierlich, B.A. Roe, F. Sanger, P.H. Schreier, A.J.H. Smith, R. Staden, I.G. Young, Nature 290 (1981) 457^465. [22] D.R. Wolstenholme, Int. Rev. Cytol. 141 (1992) 173^216. [23] H. Kasamatsu, J. Vinograd, Annu. Rev. Biochem. 43 (1974) 695^719. [24] I.G. Macreadie, C.E. Novitski, R.J. Maxwell, U. John, B.G. Ooi, G.L. McMullen, H.B. Likins, A.W. Linnane, P. Nagley, Nucleic Acids Res. 11 (1983) 4435^4451. [25] A. Chomyn, P. Mariottini, M.W.J. Cleeter, C.I. Ragan, A. Matsuno-Yagi, Y. Hati¢, R.F. Doolittle, G. Attardi, Nature 314 (1985) 592^597. [26] A. Chomyn, M.W.J. Cleeter, C.I. Ragan, M. Riley, R.F. Doolittle, G. Attardi, Science 234 (1986) 614^618. [27] D. Ojala, J. Montoya, G. Attardi, Nature 290 (1981) 470^
[28] S. Osawa, T.H. Jukes, K. Watanabe, A. Muto, Microbiol. Rev. 56 (1992) 229^264. [29] B.G. Barrel, S. Anderson, A.T. Bankier, M.H.L. De Bruijn, E. Chen, A.R. Coulson, J. Drouin, I.C. Eperon, D.P. Nier- lich, B.A. Roe, F. Sanger, P.H. Schreier, A.J.H. Smith, R. Staden, I.G. Young, Proc. Natl. Acad. Sci. USA 77 (1980) 3164^3166. [30] S.G. Bonitz, R. Berlani, G. Coruzzi, M. Li, G. Macino, F.G. Nobrega, M.P. Nobrega, B.E. Thalenfeld, A. Tzagolo°, Proc. Natl. Acad. Sci. USA 77 (1980) 3167^3170. [31] J.E. Heckman, J. Sarno°, B. Alzner-De Weerd, S. Yin, U.L. RajBhandary, Proc. Natl. Acad. Sci. USA 77 (1980) 3159^
[32] R.P. Martin, A.-P. Sibler, C.W. Gehrke, K. Kuo, C.G. Ed- monds, J.A. McCloskey, G. Dirheimer, Biochemistry 29 (1990) 956^959. [33] J. Moriya, T. Yokogawa, K. Wakita, T. Ueda, K. Nishika- wa, P.F. Crain, T. Hashizume, S.C. Pomerantz, J.A. McCloskey, G. Kawai, N. Hayashi, S. Yokoyama, K. Wa- tanabe, Biochemistry 33 (1994) 2234^2239. [34] M.W. Walberg, D.A. Clayton, Nucleic Acids Res. 9 (1981) 5411^5421. [35] K.R. Ryan, R.E. Jensen, Cell 83 (1995) 517^519.
[93] J. Montoya, D. Ojala, G. Attardi, Nature 290 (1981) 465^
[94] W. Rossmanith, A. Tullo, T. Potuschak, R. Karwan, E. Sbisa©, J. Biol. Chem. 270 (1995) 12885^12891. [95] C.-J. Doersen, C. Guerrier-Takada, S. Altman, G. Attardi, J. Biol. Chem. 260 (1985) 5942^5949. [96] D.L. Miller, N.C. Martin, Cell 34 (1983) 911^917. [97] Y.L. Dang, N.C. Martin, J. Biol. Chem. 268 (1993) 19791^
[98] C.A. Wise, N.C. Martin, J. Biol. Chem. 266 (1991) 19154^
[99] K.M. Rose, H.P. Morris, S.T. Jacob, Biochemistry 14 (1975) 1025^1032. [100] F. Amalric, C. Merkel, R. Gelfand, G. Attardi, J. Mol. Biol. 118 (1978) 1^25. [101] P. Borst, L.A. Grivell, FEBS Lett. 13 (1971) 73^88. [102] P. Cantatore, Z. Flagella, F. Fracasso, A.M.S. Lezza, M.N. Gadaleta, A. De Montalvo, FEBS Lett. 213 (1987) 144^
[103] G. Attardi, D. Ojala, Nature New Biol. 229 (1971) 133^137. [104] A. Brega, C. Vesco, Nature New Biol. 229 (1971) 137^139. [105] M.G. Hamilton, T.W. O'Brien, Biochemistry 13 (1974) 5400^5403. [106] A. Cahill, D.L. Baio, C.C. Cunningham, Anal. Biochem. 232 (1995) 47^55. [107] D.P. Nierlich, Mol. Cell. Biol. 2 (1982) 207^209. [108] D.E. Matthews, R.A. Hessler, N.D. Denslow, J.S. Ed- wards, T.W. O'Brien, J. Biol. Chem. 257 (1982) 8788^8794. [109] H.-R. Graak, B. Wittmann-Liebold, Biochem. J. 329 (1998) 433^448. [110] K. Grohmann, F. Amairic, S. Crews, G. Attardi, Nucleic Acids Res. 5 (1978) 637^651. [111] C. Van den Bogert, H. De Vries, M. Holtrop, P. Muus, H.L. Dekker, M.J.M. Van Galen, P.A. Bolhuis, J.-W. Taanman, Biochim. Biophys. Acta 1144 (1993) 177^183. [112] H.L. Garstka, M. Fa´cke, J. Ramos Escribano, R.J. Wies- ner, Biochem. Biophys. Res. Commun. 200 (1994) 619^626. [113] H.-X. Liao, L.L. Spremulli, J. Biol. Chem. 264 (1989) 7518^7522. [114] E. Schmitt, J.M. Guillon, T. Meinnel, Y. Mechulam, F. Dardel, S. Blanquet, Biochimie 78 (1994) 543^554. [115] J.W.B. Hershey, Annu. Rev. Biochem. 61 (1991) 717^755. [116] N.D. Denslow, G.S. Michaels, J. Montoya, G. Attardi, T.W. O'Brien, J. Biol. Chem. 264 (1989) 8328^8338. [117] H.-X. Liao, L.L. Spremulli, J. Biol. Chem. 265 (1990) 11761^11765. [118] H.-X. Liao, L.L. Spremulli, J. Biol. Chem. 266 (1990) 13618^13622. [119] H.-X. Liao, L.L. Spremulli, J. Biol. Chem. 266 (1991) 20714^20719. [120] J. Ma, M.A. Farwell, W.A. Burkhart, L.L. Spremulli, Bio- chim. Biophys. Acta 1261 (1995) 321^324. [121] J. Ma, L.L. Spremulli, J. Biol. Chem. 270 (1995) 1859^
[122] J. Ma, L.L. Spremulli, J. Biol. Chem. 271 (1996) 5805^
[123] C.J. Schwartzbach, L.L. Spremulli, J. Biol. Chem. 264 (1989) 19125^19131. [124] H.K.J. Chung, L.L. Spremulli, J. Biol. Chem. 265 (1990) 21000^21004. [125] C. Barker, A. Makris, C. Patriotis, S.E. Bear, P.N. Tsichlis, Nucleic Acids Res. 21 (1993) 2641^2647. [126] H. Xin, V. Woriax, W. Burkhart, L.L. Spremulli, J. Biol. Chem. 270 (1995) 17243^17249. [127] J. Wells, F. Henkler, M. Leversha, R. Koshy, FEBS Lett. 358 (1995) 119^125. [128] V.L. Woriax, W. Burkhart, L.L. Spremulli, Biochim. Bio- phys. Acta 1264 (1995) 347^356. [129] M. Ling, F. Merante, H.-S. Chen, C. Du°, A.M.V. Dun- can, B.H. Robinson, Gene 197 (1997) 325^336. [130] K.H. Nierhaus, Nature 379 (1996) 491^492. [131] V.L. Woriax, J.M. Bullard, L. Ma, T. Yokogawa, L.L. Spremulli, Biochim. Biophys. Acta 1352 (1997) 91^101. [132] http://in¢nity.gen.emory.edu/mitomap.html/ [133] J.-W. Taanman, A.G. Bodnar, J.M. Cooper, A.A.M. Mor- ris, P.T. Clayton, J.V. Leonard, A.H.V. Schapira, Hum. Mol. Genet. 6 (1997) 935^942. [134] J.M. Sho°ner, M.T. Lott, A.M.S. Lezza, P. Seibel, S.W. Ballinger, D.C. Wallace, Cell 61 (1990) 931^937. [135] A. Chomyn, G. Meola, N. Bresolin, S.T. Lai, G. Scarlato, G. Attardi, Mol. Cell. Biol. 11 (1991) 2236^2244. [136] L. Boulet, G. Karpati, E.A. Shoubridge, Am. J. Hum. Gen- et. 51 (1992) 1187^1200. [137] Y. Koga, M. Davidson, E.A. Schon, M.P. King, Nucleic Acids Res. 21 (1993) 657^662. [138] E.A. Schon, Y. Koga, M. Davidson, C.T. Moraes, M.P. King, Biochim. Biophys. Acta 1101 (1992) 206^209. [139] Y. Koga, M. Davidson, E.A. Schon, M.P. King, Muscle Nerve 3 (Suppl.) (1995) S119^S123. [140] J.P. Masucci, M. Davidson, Y. Koga, E.A. Schon, M.P. King, Mol. Cell. Biol. 15 (1995) 2872^2881. [141] J.A. Enriquez, A. Chomyn, G. Attardi, Nature Genet. 10 (1995) 47^55. [142] H. Hao, C.T. Moraes, J. Biol. Chem. 271 (1996) 2347^
[143] H. Hao, C.T. Moraes, Mol. Cell. Biol. 17 (1997) 6831^
[144] J.P. Masucci, E.A. Schon, M.P. King, Mol. Cell. Biochem. 174 (1997) 215^219. [145] D.A. Clayton, Cell 28 (1982) 693^705. [146] D.D. Chang, D.A. Clayton, Proc. Natl. Acad. Sci. USA 82 (1985) 351^355. [147] D.D. Chang, W.W. Hauswirth, D.A. Clayton, EMBO J. 4 (1985) 1559^1567. [148] B. Xu, D.A. Clayton, EMBO J. 15 (1996) 3135^3143. [149] G.S. Shadel, D.A. Clayton, Annu. Rev. Biochem. 66 (1997) 409^435. [150] D.D. Chang, D.A. Clayton, Science 235 (1987) 1178^1184. [151] D.D. Chang, D.A. Clayton, EMBO J. 6 (1987) 409^417. [152] D.Y. Lee, D.A. Clayton, Genes Dev. 11 (1997) 582^592. [153] M.E. Schmitt, D.A. Clayton, Mol. Cell. Biol. 13 (1993) 7935^7941.
[154] S. Chu, R.H. Archer, J.M. Zengel, L. Lindahl, Proc. Natl. Acad. Sci. USA 91 (1994) 659^663. [155] Z. Lygerou, C. Allmang, D. Tollervey, B. Se®raphin, Science 272 (1996) 268^270. [156] T. Kiss, W. Filipowicz, Cell 70 (1992) 11^16. [157] J.N. Topper, J.L. Bennett, D.A. Clayton, Cell 70 (1992) 16^
[158] K. Li, C.S. Smagula, W.J. Parsons, J.A. Richardson, M. Gonzalez, H.K. Hagler, R.S. Williams, J. Cell. Biol. 124 (1994) 871^882. [159] M.E. Schmitt, D.A. Clayton, Genes Dev. 6 (1992) 1975^
[160] J.L. Paluh, D.A. Clayton, EMBO J. 15 (1996) 4723^4733. [161] J. Co™ te®, A. Ruiz-Carrillo, Science 261 (1993) 765^769. [162] W.O. Cummings, T.C. King, J.A. Holden, R.L. Low, J. Biol. Chem. 262 (1987) 2005^2015. [163] E. Prats, M. Noe´l, J. Le®tourneau, V. Tiranti, J. Vaque®, R. Debo® n, M. Zeviani, L. Cornudella, A. Ruiz-Carrillo, DNA Cell Biol. 16 (1997) 1111^1122. [164] M. Gerschenson, K.L. Houmiel, R.L. Low, Nucleic Acids Res. 23 (1995) 88^97. [165] H.P. Zassenhaus, T.J. Hofmann, R. Uthayashanker, R.D. Vincent, M. Zona, Nucleic Acids Res. 16 (1988) 3283^3296. [166] J.N. Doda, C.T. Wright, D.A. Clayton, Proc. Natl. Acad. Sci. USA 78 (1981) 6116^6120. [167] M. Roberti, C. Musicco, P. Loguercio Polosa, F. Milella, M.N. Gadaleta, P. Cantatore, Biochem. Biophys. Res. Commun. 243 (1998) 36^40. [168] C.S. Madsen, S.C. Ghivizzani, W.W. Hauswirth, Mol. Cell. Biol. 13 (1993) 2162^2171. [169] P.A. Martens, D.A. Clayton, J. Mol. Biol. 135 (1979) 327^
[170] D.P. Tapper, D.A. Clayton, J. Biol. Chem. 256 (1981) 5109^5115. [171] J.E. Hixson, T.W. Wong, D.A. Clayton, J. Biol. Chem. 261 (1986) 2384^2390. [172] T.W. Wong, D.A. Clayton, Cell 42 (1985) 951^958. [173] T.W. Wong, D.A. Clayton, Cell 45 (1986) 817^825. [174] A. Bolden, G.P. Noy, A. Weissbach, J. Biol. Chem. 252 (1977) 3351^3356. [175] F. Foury, J. Biol. Chem. 264 (1989) 20552^20560. [176] T.S.-F. Wang, Annu. Rev. Biochem. 60 (1991) 513^552. [177] H. Gray, T.W. Wong, J. Biol. Chem. 267 (1992) 5835^5841. [178] M.W. Olsen, Y. Wang, R.H. Elder, L.S. Kaguni, J. Biol. Chem. 270 (1995) 28932^28937. [179] F. Ye, J.A. Carrodeguas, D.F. Bogenhagen, Nucleic Acids Res. 24 (1996) 1481^1488. [180] P.A. Ropp, W.C. Copeland, Genomics 36 (1996) 449^458. [181] N. Lecrenier, P. Van der Bruggen, F. Foury, Gene 185 (1997) 147^152. [182] S.W. Graves, A.A. Johnson, K.A. Johnson, Biochemistry 37 (1998) 6050^6058. [183] V. Tiranti, M. Rocchi, S. DiDonato, M. Zeviani, Gene 126 (1993) 219^225. [184] E. Van Dyck, F. Foury, B. Stillman, S.J. Brill, EMBO J. 11 (1992) 3421^3430.
[185] G.D. Hoke, P.A. Pavco, B.J. Ledwith, G.C. Van Tuyle, Arch. Biochem. Biophys. 282 (1990) 116^124. [186] A.J. Williams, L.S. Kaguni, J. Biol. Chem. 270 (1995) 860^
[187] G. Webster, J. Genschel, U. Curth, C. Urbanke, C. Kang, R. Hilgenfeld, FEBS Lett. 411 (1997) 313^316. [188] U. Curth, C. Urbanke, J. Greipel, H. Gerberding, V. Ti- ranti, M. Zeviani, Eur. J. Biochem. 221 (1994) 435^443. [189] C. Yang, U. Curth, C. Urbanke, C. Kang, Nature Struct. Biol. 4 (1997) 153^157. [190] S.W. Matson, K.A. Kaiser-Rogers, Annu. Rev. Biochem. 59 (1990) 289^329. [191] G.L. Hehman, W.W. Hauswirth, Proc. Natl. Acad. Sci. USA 89 (1992) 8562^8566. [192] M. Roberti, C. Musicco, P. Loguercio Polosa, M.N. Gada- leta, P. Cantatore, Biochem. Biophys. Res. Commun. 219 (1996) 134^139. [193] J.C. Wang, Annu. Rev. Biochem. 65 (1996) 635^692. [194] F.J. Castora, G.M. Lazarus, D. Kunes, Biochem. Biophys. Res. Commun. 130 (1985) 854^866. [195] M.J. Kosovsky, G. Soslau, Biochim. Biophys. Acta 1078 (1991) 56^62. [196] M.J. Kosovsky, G. Soslau, Biochim. Biophys. Acta 1164 (1993) 101^107. [197] A. Tua, J. Wang, V. Kulpa, C.M. Wernette, Biochimie 79 (1997) 341^350. [198] J. Wang, K. Kearney, M. Derby, C.M. Wernette, Biochem. Biophys. Res. Commun. 214 (1995) 723^729. [199] K. Komori, K. Kuroe, K. Yanagisawa, Y. Tanaka, Bio- chim. Biophys. Acta 1352 (1997) 63^72. [200] F.J. Castora, M.V. Simpson, J. Biol. Chem. 254 (1979) 11193^11195. [201] J.W. Lawrence, D.C. Claire, V. Weissig, T.C. Rowe, Mol. Pharmacol. 50 (1996) 1178^1188. [202] F. Caron, C. Jacq, J. Rouvie©re-Yaniv, Proc. Natl. Acad. Sci. USA 76 (1979) 4265^4269. [203] L. DeFrancesco, G. Attardi, Nucleic Acids Res. 9 (1981) 6017^6030. [204] J.F.X. Di•ey, B. Stillman, Proc. Natl. Acad. Sci. USA 88 (1991) 7864^7868. [205] N.-G. Larsson, J. Wang, H. Wilhelmsson, A. Oldfors, P. Rustin, M. Lewandoski, G.S. Barsh, D.A. Clayton, Nature Genet. 18 (1998) 231^236. [206] N.-G. Larsson, A. Oldfors, E. Holme, D.A. Clayton, Bio- chem. Biophys. Res. Commun. 200 (1994) 1374^1381. [207] J. Poulton, K. Morten, C. Freeman-Emmerson, C. Potter, C. Sewry, V. Dubowitz, H. Kidd, J. Stephenson, W. White- house, F.J. Hansen, M. Parisi, G. Brown, Hum. Mol. Gen- et. 3 (1994) 1763^1769. [208] A.F. Davis, P.A. Ropp, D.A. Clayton, W.C. Copeland, Nucleic Acids Res. 24 (1996) 2753^2759. [209] R.A. Schultz, S.J. Swoap, L.D. McDaniel, B. Zhang, E.C. Koon, D.J. Garry, K. Li, R.S. Williams, J. Biol. Chem. 273 (1998) 3447^3451. [210] N.-G. Larsson, J.D. Garman, A. Oldfors, G.S. Barsh, D.A. Clayton, Nature Genet. 13 (1996) 296^302.