¡Descarga Cuestionario Citología y más Apuntes en PDF de Citología e Histología Vegetal y Animal solo en Docsity!
PREGUNTAS
1. Analogías y diferencias entre células eucarióticas y procarióticas. ANALOGÍAS: Código genético común (ADN) que transcriben en ARN y traducen en proteínas. Proteínas y ARN para catálisis química. ATP moneda energética Delimitan su territorio mediante una membrana de fosfolípidos: membrana plasmática. Ésta las separa y comunica con el exterior, controla los movimientos celulares y mantiene el potencial eléctrico de la célula. Algunas células como bacterias y células vegetales poseen una pared celular que la rodea. Contienen un medio hidrosalino, el citoplasma, que forma la mayor parte del volumen celular y en el que están inmersos los orgánulos celulares. DIFERENCIAS: EUCARIOTAS: Citoplasma se encuentra compartimentalizado (endomembranas): Separación Núcleo/Citoplasma. Orgánulos membranosos (Mitocondrias, plastos), Flagelos complejos, Ribosomas 80S. ADN asociado a proteínas histonas. PROCARIOTAS: Compartimento único (escasa organización estructural) ni Mitocondrias ni Plastos, Flagelos simples, Ribosomas 70S. Dos billones de años más evolucionando. Variabilidad adaptativa al ambiente. Distintas estrategias para la obtención de la energía: Existen especies de bacterias que pueden utilizar como alimento prácticamente cualquier tipo de molécula orgánica, incluidos azúcares, aminoácidos, grasas, hidratos de carbono, polipéptidos y polisacáridos. Algunas son capaces incluso de obtener los átomos de carbono del CO2 y 1os átomos de nitrógeno del N2. A pesar de su simplicidad relativa, las bacterias han sobrevivido durante más tiempo que cualquier otro organismo y todavía constituyen el tipo de células más abundante de la Tierra. 2. ¿Qué se entiende por organismos: Organotrofos, Fototrofos, Litotrofos, Aerobios y Anaerobios? Clasificación nutricional básica La clasificación nutricional de un organismo se realiza en base a tres criterios importantes: el origen del carbono, la fuente de energía y los donadores de electrones. Fuente del carbono. Se refiere a la fuente del carbono usada por el organismo para su crecimiento y desarrollo. Un organismo se denomina HETERÓTROFO si usa compuestos orgánicos y AUTÓTROFO si su fuente del carbono es el dióxido de carbono (CO 2 ). Donador de electrones Se refiere a los compuestos donadores de electrones que se utilizarán en la biosíntesis (por ejemplo, en forma de NADH o NADPH). Un organismo se denomina ORGANOTROFO cuando utiliza compuestos orgánicos como fuente de electrones, mientras que se denomina LITOTROFO cuando utiliza compuestos inorgánicos. Los organismos organotrofos son a menudo también heterótrofos, y así usan compuestos orgánicos como fuente de electrones y de carbono al mismo tiempo (animales, hongos, bacterias parásitas intestinales). De forma similar, los organismos litotrofos son a menudo también autótrofos, con fuentes inorgánicas de electrones y dióxido de carbono como fuente inorgánica del carbono. (Bacterias) Fuente de energía. Se refiere al método empleado por el organismo para producir ATP, que se requiere para aprovisionar de combustible los caminos anabólicos de biosíntesis de los componentes de la célula.
Un organismo es FOTÓTROFO cuando utiliza luz como fuente de energía (plantas algas, muchas bacterias). Proporcionan O2 a la atmósfera. Un organismo es QUIMIOTROFO cuando obtiene la energía de reacciones con compuestos químicos. Se denominan AEROBIOS o aeróbicos a los organismos que necesitan del oxígeno diatómico para vivir o poder desarrollarse. Los organismos ANAEROBIOS o anaeróbicos son los que no utilizan oxígeno (O2) en su metabolismo (condiciones de atmósfera primitiva), más exactamente que el aceptor final de electrones es otra sustancia diferente del oxígeno. Si el aceptor de electrones es una molécula orgánica (piruvato, acetaldehído, etc.) se trata de metabolismo fermentativo; si el aceptor final es una molécula inorgánica distinta del oxígeno (sulfato, carbonato, etc.) se trata de respiración anaeróbica.
3. ¿Qué son las ribozimas y qué papel jugaron en el origen de la vida primitiva? Son moléculas de ARN con capacidad catalítica y, al tiempo, con la información necesaria para la replicación; podrían haber catalizado una síntesis primitiva, no codificada de cortas proteínas. Una ribozima es una molécula de ARN con capacidad catalítica. El término ribozima deriva de la combinación en zima de ácido ribo nucleico. El sustrato de esta enzima es, con frecuencia, una molécula de ARN_._ Se han descrito ribozimas en virus, en procariotas y en eucariotas. Una molécula de RNA tiene dos características especiales: transporta información codificada en su secuencia de nucleótidos que puede transmitir mediante el proceso de replicación y tiene una estructura plegada única que determina la manera en que interactuará con otras moléculas y responderá a las condiciones ambientales. Estos dos rasgos (uno de información y otro de función) son las dos propiedades esenciales que se necesitan para la evolución. Las ribozimas actúan como catalizadores, es decir, aceleran reacciones de forma específica. Al igual que las enzimas proteicas, poseen un centro activo que se une específicamente a un sustrato y que facilita su conversión en un producto. Las ribozimas además de actuar como patrón para su propia replicación, pueden catalizar la rotura y formación de enlaces covalentes, incluyendo uniones entre nucleótidos. Pueden por ejemplo cortar y eliminar secuencias determinadas de nucleótidos y reunir los extremos cortados (proceso de automaduración por corte y empalme o "self splicing"). Ciertas actividades catalíticas pudieron tener una importancia capital en el caldo primordial. Consideremos una molécula de RNA que catalice el proceso de polimerización utilizando cualquier molécula de RNA como patrón (actividad ribozima demostrada in vitro). Ésta molécula catalítica puede actuar sobre copias de sí misma o promover la replicación de otras moléculas de RNA distintas. Estas pueden realizar a su vez funciones de ribozima. Así si los efectos beneficiosos son recíprocos diferentes tipos de moléculas de RNA, especializados en diferentes actividades, pueden desarrollar un sistema cooperativo que se replique con eficacia. La hipótesis del mundo de ARN sobre el origen de la vida está basada en la función catalítica que pueden desarrollar las moléculas de RNA en forma de ribozimas. Supone que el ARN fue la primera forma de vida en la Tierra, desarrollando posteriormente una membrana celular a su alrededor y convirtiéndose así en la primera célula procariota. Hay ribozimas implicadas en diversas e importantes reacciones celulares; destacan entre ellas el procesamiento o maduración del ARN y la síntesis de proteínas (una de las moléculas de ARNr componentes del ribosoma cataliza la reacción de transpeptidación, o unión de los aminoácidos).
metabolismo energético de protistas sin mitocondrias. Pero no el origen de los plastidios.
7. Explica el significado de los organismos eucariotas actuales que carecen de mitocondrias y/o plastos. Siguiendo las teorías endosimbiontes, Las células eucariotas contienen en principio mitocondrias, orgánulos que habrían adquirido por endosimbiosis de ciertas bacterias primitivas, lo que les dota de la capacidad de desarrollar un metabolismo aerobio. Sin embargo, por ejemplo, en algunos eucariotas del reino protistas las mitocondrias han desaparecido secundariamente en el curso de la evolución, en general derivando a otros orgánulos, como los hidrogenosomas. Por lo tanto, la existencia de células sin ellos se explica con la teoría de que sí que tuvieron plastos o mitocondrias anteriormente pero al evolucionar los perdieron. Esto puede ocurrir por consecuencia de regresiones tales como hidrogenosomas o mitosomas, o por adaptaciones a ambientes pobres en O2. 8. Fluidez de los lípidos de membrana: diferencias entre los datos obtenidos con bicapas artificiales y en estudios fisiológicos. Bicapas artificiales Hay dos tipos de bicapas lipídicas artificiales que han resultado muy útiles para los estudios experimentales: los liposomas (vesículas esféricas) y membranas negras (bicapas planas formadas a través de un agujero situado en la separación de dos compartimentos acuosos). Estos estudios demuestran que las moléculas de fosfolípido de las bicapas artificiales raramente migran de un lado a otro de la monocapa. Este proceso denominado flip-flop se produce menos de una vez al mes en cualquier molécula lipídica. Por otro lado, las moléculas lipídicas intercambian fácilmente su lugar con el de las moléculas vecinas dentro de una monocapa. Ello da lugar a una rápida difusión lateral (una molécula lipídica promedio difunde la longitud de una gran célula bacteriana en un segundo). Además estos estudios indican que las moléculas lipídicas giran con gran rapidez alrededor de sus ejes longitudinales y que sus cadenas hidrocarbonadas son flexibles. Estudios con bicapas artificiales solo con liposomas y membranas "negras" solo tienen en cuenta los lípidos y no sus interacciones con otros componentes (por ej. Colesterol). Estudios fisiológicos Unos estudios similares han sido realizados con moléculas lipídicas marcadas en membranas biológicas aisladas y en células enteras relativamente sencillas como micoplasmas, bacterias y glóbulos rojos anucleados. En general, los resultados de estos estudios son los mismos que se obtienen con bicapas artificiales y demuestran que el componente lipídico de las membranas biológicas es un líquido bidimensional en el que las moléculas constituyentes son libres de moverse lateralmente: al igual que las bicapas sintéticas, las moléculas de fosfolípido se hallan restringidas a su propia monocapa. Este confinamiento crea un problema para su síntesis. Los fosfolípidos son sintetizados sólo en una de las monocapas de la membrana, la cual se sintetiza principalmente en la monocapa citosólica de la membrana del retículo endoplasmático; si ninguna de estas moléculas recientemente formadas pudiera migrar hacia la otra mitad de la bicapa lipídica, no se podría formar más bicapa. El problema se soluciona gracias a un tipo esencial de enzimas unidas al RE llamadas translocadoras de fosfolípidos, que catalizan un rápido flip-flop de fosfolípidos específicos desde la monocapa donde han sido sintetizados hasta la monocapa opuesta. ⇒ Flippasas: de hoja externa a interna ⇒ Floppasas: de hoja interna a externa ⇒ Revoltijasas o Scramblasas: bidireccional pero solo a favor de gradiente.
Factores en fluidez Colas no polares de FL agregan debido a su hidrofobicidad y a fuerzas de Van der Waals: largas y saturadas (menor fluidez), cortas (mayor fluidez), insaturadas (mayor fluidez) Colesterol presente en todas las membranas biológicas: A altas concentraciones la interacción del anillo esteroídico con colas lipídicas inmoviliza las colas (menor fluidez), a bajas concentraciones la interacción cesa (mayor fluidez). En general el colesterol genera menor fluidez pero su interposición entre las cadenas hidrocarbonadas impide sus interacciones aumentando su fluidez. A pesar de su fluidez las bicapas organizan distintos dominios con composiciones distintas (rafts) Esfingomielina crea dominios con los lípidos adyacentes a los que acumula. In vivo, todos los FL difunden libremente hasta 0,5 microm pero no mayores distancias. Regiones ricas en proteínas (1microm) alternan con regiones ricas en lípidos donde estos difunden libremente (cómo se organizan, mantienen estas separaciones??) Difícil extrapolar los resultados de las bicapas artificiales a la situación in vivo.
9. Definir temperatura de transición señalando como contribuyen a ella los distintos lípidos de membrana. La composición de la membrana determina su temperatura de transición. Ésta es la T a la que una bicapa lipídica pasa de sol a gel. Se dice que una membrana tiene una Tt baja si es más difícil de "congelar" osea, si tiene FL de cadenas hidrocarbonadas cortas y/o insaturadas y es, por tanto, fluida. Organismos dependientes de la T ambiente y de sus fluctuaciones ajustan la composición lipídica de sus membranas para mantenerlas fluidas: cuando cae la T incorporan a ellas lípidos insaturados y evitan, de ese modo, el descenso de la fluidez. Contribución de los distintos lípidos de membrana: Aumentan su Tt: Lípidos de colas apolares largas y colesterol Disminuyen su Tt: Lípidos de colas apolares cortas como la de Fosfatidil Colina. Cuatro grupos de fosfolípidos predominan en la membrana plasmática de muchas células de mamíferos: fosfatidilcolina, esfingomielina, fosfatidilserina y fosfatidiletanolamina. Sólo la fosfatidilserina tiene una carga neta negativa, el resto son eléctricamente neutras a pH fisiológico, presentando una carga negativa y otra positiva. La fluidez de membrana depende tanto de su composición como de la temperatura. Una bicapa sintética producida a partir de un único tipo de fosfolípido, pasa de un estado líquido a un estado cristalino rígido (o gel) en un punto de congelación característico. Este cambio de estado recibe el nombre de transición de fase y la temperatura la que se produce es más baja (es decir, la membrana resulta más difícil de congelar) si las cadenas hidrocarbonadas son cortas o tienen dobles enlaces cis. Una menor longitud de la cadena reduce la tendencia de las colas hidrocarbonadas a interaccionar entre sí, y los dobles enlaces cis producen pliegues en las cadenas hidrocarbonadas que dificultan su empaquetamiento, de forma que las membranas permanecen fluidas a temperaturas más bajas. Pero la bicapa de muchas membranas celulares contiene habitualmente colesterol y glucolípidos. Las membranas eucariotas contienen cantidades especialmente elevadas de colesterol, hasta una proporción de más de una molécula de colesterol por cada molécula de fosfolípido. Las moléculas de colesterol refuerzan el carácter de barrera permeable de la bicapa lipídica. Se orientan en la bicapa con sus grupos hidroxilo próximos a las cabezas polares de las moléculas de fosfolípidos; sus anillos esteroides, planos y rígidos, interactúan con (y en parte inmovilizan) las regiones de las cadenas hidrocarbonadas que son más cercanas a los grupos polares de la cabeza, dejando el
12. Movimientos de las proteínas en las bicapas lipídicas. Como ocurre con los lípidos de membrana, las proteínas no tienen movimiento de flip- flop, no saltan a través de la bicapa sino que difunden rotacionalmente alrededor de un eje perpendicular al plano de la bicapa. Además, muchas proteínas de membrana son capaces de desplazarse lateralmente por la membrana mediante difusión lateral (más restringida de lo que se pensaba). La primera evidencia de que algunas proteínas de la membrana plasmática son móviles en el plano de la membrana fue obtenida en 1970 mediante un experimento en el que se fusionaron artificialmente células de ratón con células humanas, produciendo células híbridas (heterocariontes). Para distinguir las proteínas de membrana plasmática de ratón de las humanas se utilizaron dos anticuerpos con distinto marcaje. Aunque al principio las proteínas de ratón y las humanas estaban confinadas a su propia mitad del heterocarionte recién formado, las dos clases de proteínas difundieron y se mezclaron ocupando, a la media hora, toda la superficie del heterocarionte.
La velocidad de difusión lateral de proteínas de membrana puede medirse utilizando la técnica de recuperación de fluorescencia después de fotoblanqueamiento, cuantificación FRAP. Habitualmente este método comporta el marcaje de la proteína de superficie que se pretende estudiar, con un ligando específico fluorescente, como un anticuerpo fluorescente. Luego, el ligando fluorescente es blanqueado en una pequeña área mediante un rayo láser, y se mide el tiempo que tardan las proteínas de membrana adyacentes que transportan moléculas de anticuerpo fluorescente no blanqueadas, hasta difundir en el área blanqueada. A partir de estas mediciones se pueden calcular los coeficientes de difusión de la proteína de superficie celular que había sido marcada. Y también se puede medir por cuantificación FLIP (perdida de fluorescencia con fotoblanqueamiento continuado). En esta técnica láser blanquean constantemente una pequeña área de la membrana, las moléculas fluorescentes adyacentes migran allí pero son de nuevo blanqueadas. Con cualquiera de las técnicas utilizadas es necesario añadir que los niveles de difusión son muy distintos para distintas proteínas y para distintas células; probablemente porque en cada tipo celular, las interferencias de unas proteínas con otras son diferentes. Por otro lado Muchas células tienen sistemas que les permiten limitar sus proteínas de membrana en dominios específicos de la bicapa lipídica continua. El modelo de mosaico fluido es una sobre-simplificación. Células confinan moléculas específicas en áreas concretas de la membrana plasmática: ⇒ Células polarizadas (célula epitelial) ⇒ Bacteriorodopsina en grandes cristales bidimensionales donde proteínas están relativamente inmóviles. Células epiteliales polarizadas (intestino, túbulos renales): Enzimas y proteínas en el borde apical y otras proteínas en las zonas laterales. En el dibujo proteína A y proteína B. Las uniones estrechas impiden la libre difusión. Proteínas A y B pueden difundir lateralmente en sus dominios respectivos pero no pueden entrar en el otro. La composición lipídica de estos dos dominios de membrana es diferente lo cual demuestra que las células epiteliales pueden evitar la difusión de moléculas lipídicas y proteicas entre los dominios.
Movimiento de Rafts lipídicos conlleva movimiento de las proteínas que se encuentren en ellos (en bloque).
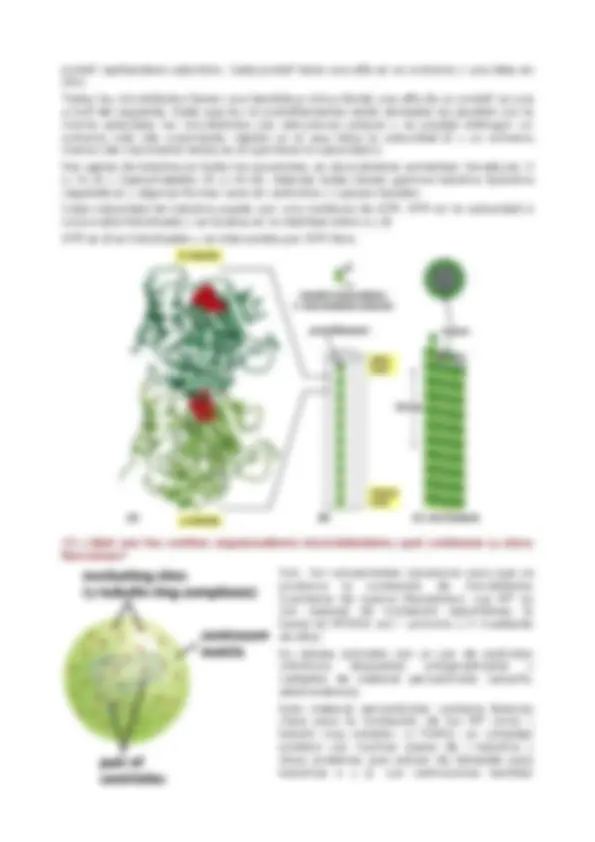
13. Esqueleto cortical de la membrana plasmática eritrocitaria: composición, organización, regulación. El córtex de los eritrocitos es una estructura relativamente simple y regular. Son pequeños y tienen una forma aplanada bicóncava distintiva, Su componente principal del córtex es la espectrina, una molécula cilíndrica larga, fina, flexible de acerca de 100 micrometros de longitud. Forma una malla que da soporte a la membrana plasmática y mantiene la forma celular. La malla de espectrina está conectada a la membrana con proteínas de unión que la unen a proteínas transmembrana (banda 3, glicoforinas). La espectrina es un heterodímero formado por dos subunidades estructuralmente similares. Los heterodímeros se asocian cabeza con cabeza, formando tetrámeros de 200 nm de largo. Las colas de 5 o 6 tetrámeros se enlazan entre sí mediante su unión a filamentos cortos de actina y a otras proteínas citoesqueléticas (banda 4.1 y ankirina) formando un "complejo de unión". El resultado final es una red deformable que se extiende por toda la superficie citoplásmica de la membrana. Este citoesqueleto basado en la espectrina permite al eritrocito resistir el estrés que sufre su membrana a medida que es empujado a través de los estrechos capilares. Ratones y humanos que poseen anomalías genéticas en la espectrina presentan anemia y sus glóbulos rojos son esféricos (en lugar de cóncavos) y anormalmente frágiles. La severidad de la anemia se incrementa con el grado de deficiencia de la espectrina. Otras patologías eritrocitarias
son: Esferocitosis hereditaria, eliptosis hereditaria, acantocitosis, esferoacantocitosis, estomatocitosis. Estan relacionadas también con otras proteínas implicadas además de la espectrina como la ankirina, glicoforina, proteína 4.2 y proteína banda 3. Además de la espectrina, la actina (otra molécula cilíndrica) también se une a las proteínas transmembrana de la MP eritrocitaria (glicoforinas y banda 3) Existen dos complejos implicados en dichas uniones: COMPLEJO ANKIRINA La ankirina es la proteína principalmente responsable de sujetar el citoesqueleto de la espectrina a la membrana plasmática del eritrocito. Se trata de una proteína intracelular de unión que se une tanto a la espectrina como al dominio citoplasmático de la proteína transmembrana Banda 3. También participa la proteína 4.2. La ankirina conecta algunas moléculas de la proteína Banda 3 a la espectrina, uniendo así la red de espectrina a la membrana, también reduce notablemente la velocidad de difusión de las moléculas de banda 3 en la bicapa lipídica. COMPLEJO 4.1R El citoesqueleto de espectrina también se une a la membrana a través de otro mecanismo que depende de la proteína 4.1. Esta proteína, que une a espectrina y actina, también se une al dominio citoplásmico de la proteína Banda 3 y a la Glucoforina, la otra proteína transmembrana mayoritaria en eritrocitos. Los filamentos de actina en el córtex del eritrocito son muy cortos y tan solo actúan como elementos de entrecruzamiento entre los tetrámeros de espectrina En este complejo participan otras proteínas: Demantina (proteína 4.9) adducina, tropomodulina, tropomiosina, p55: P55 y demantina estabilizan los vértices de la red. Adducina tropomiosina, y tropomodulina estabilizan las interacciones espectrina-actina.
14. Características de la G-actina y de los filamentos que la forman.
NUCLEACIÓN: Cuando se añade K+ y Mg 2+ a la actina monomérica en presencia de ATP se produce la fase lenta (lag) mientras se nuclean los nuevos filamentos. Se forman oligómeros cortos de 3 subunidades. La fase lag es debida a la misma barrera cinética para la nucleación que existe para la polimerización de la tubulina. La velocidad de nucleación es proporcional al cubo de la concentración de actina por lo que la estructura de nucleación para la polimeración espontánea de la actina pura es un trímero de moléculas de actina. Este periodo puede ser eliminado añadiendo un F-Act corto a la solución de G-actina. ELONGACIÓN: Oligómeros cortos actuan de nucleadores y se produce un aumento rápido de la longitud de F-Actina (fase de polimerización rápida).La velocidad con la cual un filamento se alarga es proporcional a la concentración de la subunidad libre indicando que el filamento se alarga mediante adición de moléculas de actina, de una en una.
La velocidad de polimerización es distinta en los dos extremos del filamento de actina: el extremo más (barbado) polimeriza unas 10 veces más rápidamente que el extremo menos (puntiagudo). La concentración crítica para la polimerización de la actina (concentración del monómero de actina libre en la cual la proporción de actina polimerizada detiene su incremento) está alrededor de 0 ,2 micromolar. Esta concentración es mucho más baja que la concentración de actina no polimerizada de la célula, por lo que la célula ha desarrollado mecanismos especiales para impedir que la mayoría de esa actina monomérica se ensamble formando filamentos. Rápidamente después de la polimerización, el fosfato terminar del ATP unido a la molécula de actina es hidrolizado, quedando ADP atrapado en el polímero. La hidrólisis de ATP produce cambios conformacionales en la molécula de actina. Ésta tiene forma de almeja, y el ATP se encuentra situado en la charnela situada entre las dos mitades; como la concha de una almeja, puede abrirse y cerrarse. Cuando la actina polimeriza, la concha está totalmente cerrada por interacciones entre aminoácidos de las dos valvas de la almeja, y la zona posterior de la siguiente subunidad del polímero. La hidrólisis del ATP se desencadena por el cierre de la concha mientras cada molécula de actina se incorpora al filamento y deja el ADP incorporado dentro STEADY-STATE PHASE: Monómeros de G-actina se intercambian con subunidades de los extremos del F-act sin que ello represente un cambio neto de la masa total del filamento. En este momento se produce el triturado de subunidades: El nivel de ensamblaje (pero no el de desensamblaje) depende de la concentración de G-actina/ATP. En (+) se necesita menos concentración de G-act para polimerizar F-act que en (-) (10 veces más lento polimerizando). Por lo tanto a medida se va elongando en (+), la [G-act] ira descendiendo hasta alcanzar su concentración crítica. En (-) en cambio, el nivel de ensamblaje es bajo y similar al de desensamblaje, por debajo de su Cc el F-act no crece en (-) pero sigue haciéndolo en (+). Por lo tanto en la condición steady (equilibrio), (+) crecerá y (-) disminuirá, fenómeno que se denomina Triturado de subunidades. No se pierde longitud total del filamento, pero se intercambian subunidades y el filamento se va desplazando hacia la dirección del extremo (+).
Proteínas capping bloquean el ensamblaje/desensamblaje de sus extremos: CapZ, Tropomodulina Cap Z: 2 subunidades intimamente relacionadas que se unen rápidamente y con gran afinidad los extremos (+) recien formados. Para evitar el bloqueo total del crecimiento del F-actina otros reguladores controlan CapZ: ⇒ PI (4,5) difosfato asociado a la MP inhibe la activación de Cap Z. ⇒ Proteínas reguladoras que se unen a (+) permiten el crecimiento de F-Act pero impiden la unión de Cap Z, que no lo permite. Tropomodulina: En células que necesitan una fuerte estabilización de F-Act (citoesqueleto cortical asociado a MP, sarcómeras del músculo esquelético). En los dos ejemplos TM actúa con Tropomiosina TPM que se alinea a lo largo del F-Act para estabilizarlo. TM se une a TPM y a (-) y contribuye a la estabilización. Otras proteínas que hacen capping con F-Act Gelsolina: se une a (+) bajo control de altas [Ca++] y separa filamentos de Act entre sí. La unión de Ca++ produce un cambio conformacional en Gelsolina que le permite unirse a F-Act insertándose entre las subunidades de la doble hélice y rompiendo el F- Act. Gel queda unida a (+) y crea (-) donde se desensambla el filamento.
Proteínas que hacen crosslinking con Act convierten una solución de F-Act en un gel, mientras Gelsolina, en presencia de Ca, se une a F-Actina y lo revierte a sol. (Gel-sol ina). Mecanismos de ensamblaje de los filamentos de actina: Proteínas nucleadoras: Familia de las Forminas: ensamblan grandes filamentos de Act (fibras stess, anillo contractil citocinesis) Complejo Arp 2/3: ensambla redes ramificadas.
17. La nucleación de F-actina se realiza in vivo con el concurso de "proteínas nucleadoras". Indica mecanismo de funcionamiento, moléculas implicadas y regulación. FORMINAS: Familia muy diversa (7 clases distintas, algunas con funciones por determinar) presentes en todos los eucariotas, con dos dominios adyacentes comunes FH1 y FH (Formin Homology Domain). FH1 se comporta como un punto de unión de los complejos P-G-Act/ATP (FH1 es rico en prolinas que unen Profilina), comportándose como una fuente de subunidades para añadir al F-act con el concurso de FH2. Capacidad de nucleación de las forminas: Dos dominios FH2 se asocian en un complejo en forma de donut que une 2 subunidades de G-Act con el (+) hacia los dominios FH2. El F-Act puede crecer por (+) mientras permanezca unido a FH2. FH2 oscila entre las dos G-Act dejando que una adicione nuevas subunidades, uniéndose entonces a ella y dejando la otra libre para una nueva adición. Forminas no solo son nucleadoras de Act sino que uniéndose a (+) impiden la unión de Cap Z que impide la elongación del filamento. Regulación de la actividad de Forminas Rho, proteína anclada a membrana en cara citosólica, inactiva está unida a GDP. Se activa por unión a GTP y une las forminas FH1 y FH2 activándolas.
Forma inactiva, plegada de Wasp no puede unirse al COO de Arp 2/3 y activarlo. Cuando se activa Cdc 42, se une GTP.y sale GDP por Ras GTP activada, Cdc se une a Wasp y lo activa, haciendolo accesible a Arp 2/3. Wasp tiene además un punto adyacente a su COO capaz de unir G-Act, facilitando la nucleación.
18. Organización de las moléculas de la familia de las miosinas (I, II, V). Todas las proteínas motoras de los filamentos de actina que han sido identificadas pertenecen a la familia de las miosinas. Fueron aisladas originalmente gracias a su capacidad para hidrolizar el ATP a ADP+Pi cuando se unen a la actina. Esta actividad motora también se puede observar con un ensayo in vitro de motilidad: moléculas de miosina adheridas a un cubre que se coloca hacia abajo sobre un porta formando una cámara por donde se hace fluir una solución de F-Act marcada. En presencia de ATP, las cabezas de miosina caminan hacia el (+) del F-Act, pero como están adheridas al cubre el movimiento de las cabezas lo que provoca es el deslizamiento de F-Act hacia (-), demostrando que S1 es suficiente para el movimiento del filamento. Todas las miosinas tienen un dominio tipo S1 como el de la miosina II que es responsable de su actividad motora. Las colas de las miosinas no contribuyen a la motilidad pero determinan qué cargo será desplazado, de manera que las colas serán distintas según el cargo que hayan de transportar. Tres grandes familias (II, I, V)que sobre el mismo principio (E ATP en movimiento) utilizan mecanismos específicos para realizar diferentes tareas: Miosina II: Implicadas en deslizamiento sobre actina. Numerosos miembros reflejan su implicación en distintos tipos de contracción en células musculares y no musculares. Todas ellas tienen dos cabezas y una larga cola, parecida a una varilla: Dos cadenas pesadas se entrelazan a través de sus dominios de cola en hélice a formando un sobreenrollamiento que da lugar a un dímero estable formado por dos cabezas y una única cola parecida a una varilla. Cada cabeza presenta tanto la actividad ATPasa como la motora. Una proteína del tipo miosina II está formada por dos cadenas pesadas idénticas, cada una de las cuales está complejada a un par de cadenas ligeras. S1: cabeza y cuello (cadenas ligeras) contiene la actividad ATPasa de la molécula y el F-actin binding site. La actividad ATPasica de S
aumenta en presencia de F-Act o es una actividad dependiente de tal presencia. S2 y LMM son la cola. La cabeza elongada de la miosina se une a un extremo de la hélice a del cuello. Las dos cadenas ligeras se disponen en la base de la cabeza, envolviendo el cuello y confiriéndole rigidez. La función más importante de la cola es la de permitir la polimerización de las moléculas formando filamentos bipolares. Esta polimerización es crucial para el desarrollo de la función de la miosina II, es decir, la de mover grupos de filamentos de actina opuestos, tal y como sucede durante la contracción muscular. Miosinas I: (8 en humanos) moléculas grandes con un número variable de L unidas al cuello. Las únicas cuyas H no se asocian a través de sus colas: una sola cabeza. Tamaño y diversidad sugieren su participación en muchas funciones. Miosinas V: (3 en humanos) 2H que resultan en dos cabezas, cuellos largos, 6L por cada H. Las colas dimerizan y tienen un dominio que une específicamente orgánulos para ser transportados.
19. Explicar someramente la base de la contracción de las sarcómeras de las células musculares esqueléticas. En ese músculo hay contracciones rápidas y repetitivas, a largas distancias y generando fuerzas capaces de mover grandes cargas. Sus células son cilíndricas, largas y multinucleadas. Cada una tiene muchas miofibrillas. Miofibrillas son los elementos contráctiles de la célula muscular. Son estructuras cilíndricas de unos 1 - 2 micrometros de diámetro que a menudo son tan largas como una célula muscular. Cada una está formada por una cadena de minúsculas unidades contráctiles o sarcómeras. En ellas se pueden ver una serie de anchas bandas claras y oscuras. Cada una está separada de la siguiente sarcómera por la banda Z (en el centro de la banda clara) Sarcómeras 2 micrometros de largo, se acortan un 70% en contracción, Filamentos delgados de actina se ensamblan con sus (+) en el disco Z (los dos sets de F-Act tienen orientaciones opuestas. Filamentos gruesos, bipolares de miosina II con las cabezas de cada mitad orientadas en sentido opuesto. Contracción muscular es el resultado de las interacciones que realiza una única molécula de miosina II con un F-Act multiplicadas por los cientos de moléculas que hay en un filamento grueso. Durante estas interacciones la hidrólisis de ATP está acoplada al movimiento de las cabezas de miosina hacia el disco Z (extremo + de la F-ACT) Como el filamento grueso es bipolar, la acción de las cabezas de miosina sucede en direcciones opuestas y va empujando el F-Act hacia el centro del filamento grueso y, por tanto, de la sarcómera.