¡Descarga El núcleo como orgánulo y más Apuntes en PDF de Biología Celular solo en Docsity!
TEMA 10. EL NÚCLEO
La presencia del núcleo es la característica principal de las células eucariotas. Para explicar su origen, se aplica la teoría endosimbiótica.
El núcleo es el orgánulo que dirige la célula, e interacciona con toda la célula, no solo con otros orgánulos que tenga alrededor.
Dentro del núcleo se encuentran:
- Cromatina: DNA condensado y proteínas.
- Nucléolo: donde se lleva a cabo la síntesis de rRNA y el acoplamiento de las subunidades ribosomales.
- Matriz nuclear.
- Nucleoplasma: iones, proteínas nucleares,…
- Envoltura nuclear: externa, interna, poro y lámina nuclear.
Normalmente hay un núcleo por célula (menos en eritrocitos de mamíferos que no hay, en hepatocitos que hay 2 ó 3 y en fibras musculares que hay hasta 100), de forma esférica (menos en leucocitos que está lobulado y en el musculo alargado), de unos 5 micrómetro y que representa un 10% del volumen celular.
Sus funciones principales son el almacenamiento de la información genética, la síntesis de RNA y el acoplamiento de las subunidades ribosomales (indirectamente permite la traducción).
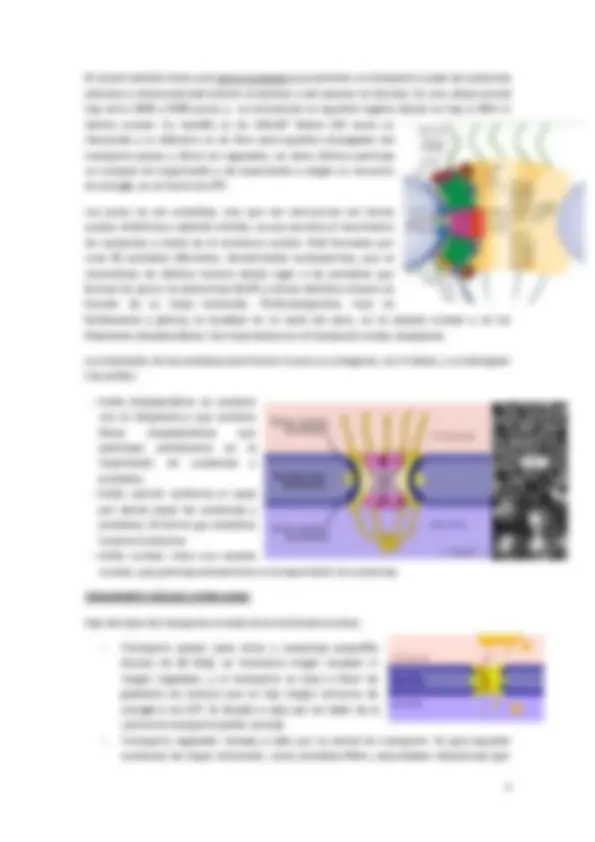
ENVOLTURA NUCLEAR.
Está formada por dos membranas, por lo que es un doble bicapa lipídica de permeabilidad selectiva que rodea al núcleo, lo delimita funcional y estructuralmente, ya que lo mantiene en una determinada región del citoplasma (aislada en cuanto a espacio), y lo comunica con el resto del citoplasma a través de los poros nucleares. Se continúa con el RE.
La membrana externa es aquella en contacto con el citosol, que se continúa con el RE y que tiene ribosomas unidos a su cara citosólica.
La membrana interna está en contacto con el Nucleoplasma, y contiene proteínas específicas que permiten la adhesión de la lámina nuclear, sólo por debajo de esta.
La lámina nuclear es un entramado fibroso (red formada por lámina proteínas A, B y C, de filamentos intermedios), que se encuentra unida a la membrana interna, y al mismo tiempo al DNA (también en forma de cromatina) solo por debajo. La lámina nuclear permite que la membrana nuclear se forma en vesícula cuando tienen lugar la división celular, de manera que la lámina nuclear se fosforila y pierde su conformación, quedando así libre el DNA durante la división celular, y además la membrana nuclear puede ahora invaginarse formando pequeñas vesículas. Cuando acaba la división celular, la lámina nuclear se desfosforila y todo vuelve a su conformación inicial.
En interfase, cuando la célula no se está dividiendo, es cuando podemos ver claramente el núcleo, ya que cuando la célula entra en división, la membrana nuclear se deforma. La fosforilación de la lámina nuclear permite que la membrana nuclear se vesiculice durante la división celular: al fosforilarse, se fragmenten las láminas nucleares y se forma pequeñas vesículas de membrana nuclear, dejando al DNA libre hasta que se acabe la división.
separado), e intervienen un receptor que regula el proceso; es activo porque requiere un gasto de energía en forma de GTP.
Todas las proteínas que realizan una función en el núcleo son fabricadas en el citoplasma, por tanto deberán de entrar en el núcleo, el cual se ha de encargar de aportar los elementos necesarios para la síntesis de la proteína: necesitamos los factores de transcripción que participan en la importación de otras proteínas. Por eso, desde el citosol se lleva a cabo una importación de proteínas hacia el núcleo, y desde este se lleva a cabo la exportación de tRNA, mRNA, rRNA y proteínas.
Hay varias diferencias en el transporte núcleo-citoplasma y otros transportes:
- Distintas proteínas transportadoras, que en este caso interaccionan con los poros.
- No se corta la secuencia señal: La NSL no se elimina una vez ha pasado la importación, ya que la proteína se ha sintetizado y madurado en el citosol, y por tanto eliminar la señal supondría eliminar una parte de la proteína que ya es funcional. Si está en el extremos es importante que se mantenga la señal para que pueda salir otra vez si fuera necesario (en este caso tendrá dos secuencias señal distintas), y si no ha de volver a salir no se corta porque, ya que el núcleo, durante la mitosis se deforma, cuando vuelva a formarse es necesario que todas las proteínas que debido a la deformación han quedado fuera, vuelvan a entrar.
IMPORTACIÓN Y EXPORTACIÓN DE PROTEÍNAS
IMPORTACIÓN DE PROTEÍNAS
Cuando una proteína es sintetizada en el citosol, es necesario que porte una secuencia que indique cuál es su destino final, donde va a ser funcional. Aquellas proteínas que se sintetizan en el citosol, lleva la señal NLS y son ricas que lisina y arginina. La importación de proteínas es un proceso mediado por receptor, y por tanto es selectivo; en el caso de importación estos receptores son la familia de las importinas α, β,...
La secuencia NLS ha de estar a la superficie de la proteína, ya que estas entran al núcleo replegadas; la secuencia es reconocida por el receptor y la transporta a través de los poros nucleares hasta el interior (...). Cuando la importina reconoce la NLS, la proteína y la importina se unen formando el P+I; este complejo se acercará al poro nuclear, de tal manera que las fibras citosólicas del poro nuclear, reconocerán la importina y permitirán el paso de la P+I. En el poro nuclear las nucleoporinas FG tienen dominios de aminoácidos que permiten la unión con la proteína. Así, la afinidad crece a medida que la proteína se introduce en el anillo central, hasta que el complejo P+I llega al Nucleoplasma. Esta entrada al núcleo se da sin gasto de energía.
Al interior del núcleo tenemos la RAN-GTP, una proteína unida a GTP (se ha unido a GTP por la acción de la RAN-GEF, que se encuentra en el interior del núcleo a intercambia su nucleótido de guanina para que la RAN-GDP pase a RAN-GTP y quede activada) que tiene mucha afinidad por las importinas, de forma que se une a la importina, provocando en ella un cambio conformacional que hace que se libere la proteína nuclear; el complejo importina +RAN GTP ha de salir del núcleo para que la importina introduzca otra proteína.
La importina y la RAN-GTP salen del núcleo interaccionando con las nucleoporinas, en el citoplasma está la RAN-GAP, y cuando sale la importina con la RAN-GTP la GAP hace que se hidrolice el GTP, y la importina se libera, pudiendo ahora entrar una nueva proteína dentro del
MODELO DE IMPROTACIÓN DE PROTEÍNAS A LA ENVOLTURA NUCLEAR
La membrana nuclear se continúa con el RE, donde se sintetizan la proteínas y se diferencian aquellas que han de llegar hasta la membrana nuclear externa. Aún no se conoce exactamente como una proteína de la membrana externa pasa hasta la membrana interna, pero si se sabe que se lleven a cabo los siguientes pasos:
- En la externa: cotraducción del RER y difusión de la proteína.
- En la interna: cotraducción del RER, reconocimiento gracias a las importinas αβ y cotraducción a través del poro nuclear.
De forma que una hipótesis es que para pasar de la externa a la interna, una NLS es reconocida por la importina, entrará por los poros con el proceso natural de importación, y al liberarse en el núcleo, quedará unida a la membrana interna.
EXPORTACIÓN DE PROTEÍNAS.
La exportación de proteínas es también un proceso importante para mantener en el núcleo solo aquello que ha de estar allí y además es necesario que salgan los componentes necesarios en el citosol para la síntesis de proteínas, al igual que aquellas proteínas que ya han llevado a cabo su función en el núcleo. Además está mediado por receptor, de forma que es selectivo y se necesita tanto el receptor, de la familia de las exportinas, como la secuencia señal NES en la proteína, que es la etiqueta que indica que la proteína ha de sacarse del núcleo; son secuencias ricas en Leu.
Ahora tenemos una proteína que tiene la secuencia NES, y para que interaccione con la exportina, la exportina ha de estar unida a la RAN-GTP (esta unión ha producido un cambio conformacional que permite a la exportina acoplar la proteína). Los tres van por los poros nucleares interaccionando con las nucleoporinas: pasan por la canasta nuclear, y luego por el anillo central.
Fuera, cuando se hidroliza el GTP por la acción de la RAN-GAP, se separan los tres elementos: la proteína queda en el citosol, la exportina vuelve al interior del núcleo, y la RAN unida a GDP también volverá dentro.
Por tanto las interacciones con las RAN GTP implican consecuencias distintas en caso de extraer proteína o de introducir proteínas.
EXPORTACIÓN DE RNA
Los tRNA y el rRNA utilizan los métodos de las proteínas para salir, ya que se unen a proteínas para salir, pero los mRNA usan orto método.
EXPORTACIÓN DE mRNA
Hay que saber que:
No interviene la RAN-GTP. No intervienen la importina y exportinas clásicas (kariophorinas), por lo que se ha complicado más la investigación. El TAP es un receptor de exportación que regula la salida de mRNA. Cuando el mRNA sale del núcleo, se produce una hidrólisis de ATP, aunque no se sabe cómo modula la afinidad. Cuando el mRNA ha salido del núcleo, el TAP se desprende y se vuelve a meter al núcleo. El mRNA es remodelado fuera del núcleo.
MATRIZ NUCLEAR
Es una red fibrilar proteica insoluble de la cual aún se desconocen muchos aspecto, como cuáles son las proteínas que la conforman o cuáles son sus funciones.
Se cree que sus funciones son estructura, organizar la cromatina y que es el lugar donde se pueden anclar las proteínas replicación y transcripción.
ORGÁNULOS SUGUNUCLEARES.
la replicación de DNA hay mucha síntesis de proteínas, por lo que se necesita muchos ribosomas, y por tanto muchos nucléolos activos.
Dentro del nucléolo podemos distinguir tres regiones:
- Centro fibrilar: transcripción de DNA que codifica los rRNA.
- Componente fibrilar denso: transcritos de rRNA, procesamiento y modificación.
- Componente glandular: ensamblaje de preribosomas.
SÍNTESIS DE RIBOSOMAS
Los ribosomas maduros son 80S, y están formados por una subunidad grande y otra pequeña. La subunidad grande es 60S y está formada por 3 tipos de rRNA, el 5S, el 28S y el 5.8S, sintetizados en el núcleo, y por unas 49 proteínas sintetizadas en el citosol. La subunidad pequeña es 40S y está formada por una rRNA 10S, sintetizado en el núcleo, y por 33 proteínas diferentes sintetizadas en el citoplasma.
Lo primero es la síntesis del precursor rRNA 45S en el nucléolo. Hay múltiples copias en tándem (unas 200 en humanos) y están repartidas en varios cromosomas, los acocéntricos NOR) y tienenuna alta frecuencia de transcripción (síntesis de RNAr muy alta).
Hay dos secuencias de transcripción y en medio DNA espaciador. Las zonas de transcripción son más gruesas porque tienen las RNA polimerasas: van leyendo el gen y al mismo tiempo van sintetizando un rRNA precursor, que es 45s; luego este se procesa, se corta y se modifica por las snoRNP, y una vez se ha modificado consigues tres subunidades ribosomales: una forma la pequeña, y dos de estas, otra que vienen de fuera y proteínas, forman la subunidad grande del ribosoma, y luego cada una de ellas sale por el poro nuclear por separado, para lo que necesitan en sus proteínas la secuencia de exportación.
El rRNA 5S se sintetiza en el Nucleoplasma, fuera del nucléolo, donde se hacen las copias en tándem.