¡Descarga Las mitocondrias y más Apuntes en PDF de Biología Celular solo en Docsity!
TEMA 9. MITOCONDRIAS.
Las mitocondrias ocupan una parte sustancial del volumen citoplasmático de las células eucariotas, y han sido esenciales para la evolución de los animales pluricelulares. Sin las mitocondrias las células animales actuales dependerían de la glucolisis anaeróbica para formar ATP. Sin embargo, cuadno la glucosa es convertida en piruvato a través de la glucolisis, sólo se libera una pequeña fracción del total de la energía libre potencialmente disponible de la glucosa. En las mitocondrias el metabolismo de los azúcares está integrado: el piruvato es importado dentro de la mitocondria y oxidado por el oxígeno molecular (O2) a CO2 y H2O. La energía liberada es almacenada de una forma tan eficiente que por cada molécula de glucosa oxidada se producen aproximadamente 30 moléculas de ATP. Por el contrario, solo dos moléculas de ATP son producidas a partir de la glucolisis.
Las mitocondrias suelen describirse como cilindros alargados y rígidos, de un diámetro comprendido entre 0,5 y 1 micrómetro y parecidos a bacterias. Sin embargo, la microcinematografía revela que las mitocondrias son orgánulos claramente móviles y plásticos, que cambian constantemente de forma. hay mitocondrias con crestas tubulares, otras longitudinales y otras transversales.
Incluso se pueden fusionar unas con otras y volverse a separar (fisión).
Por tanto las mitocondrias son unos orgánulos presentes en todas las células eucariotas, necesarios para la síntesis de ATP. El número de mitocondrias en una célula es variable, las que necesitan más aporte energético tendrán más mitocondrias, y al revés.
En general son orgánulos plásticos y dinámicos, pero en algunas células son fijas, como en espermatozoides y en los músculos.
- Mitocondrias fijas: mitocondrias del músculo esquelético, mitocondrias del músculo cardíaco y en espermatozoides. Cuadno las mitocondrias se mantienen en una cadena fija pueden proporcionar ATP
directamente a lugar de poco consumo anormalmente elevado de ATP. Por ejemplo, existen un gran número de mitocondrias empaquetadas entre las miofibrillas adyacentes en las células del músculo cardiaco, o densamente agrupadas alrededor del flagelo de espermatozoides.
- Mitocondrias móviles: cuando se desplazan dentro del citoplasma, las mitocondrias aparecen a menudo asociadas a los microtúbulos, lo que quizás determina la orientación y la distribución típica de las mitocondrias en los distintos tipos celulares. Así, las mitocondrias móviles forman largos filamentos o cadenas móviles cuando se mueven por el citoplasma, de manera que estas cadenas aparecen asociadas a los microtúbulos.
ESTRUCTURA.
La mitocondria presenta una membrana externa y una membrana interna que define dos compartimentos internos: cada mitocondria está limitada por dos membranas altamente especializadas, que desempeñan un papel crucial en las actividades de la mitocondria. Ambas membranas definen dos compartimentos: la matriz mitocondrial y un espacio intermembranoso. Cada compartimento tiene una colección determinada de proteínas.
La membrana externa , cuya composición es similar a la membrana plasmática (50% lípidos, 50% proteínas), y que contiene numerosas copias de una proteína de transporte en forma de lámina Beta, la porina , que forma grandes canales acuosos a través de la bicapa lipídica, de forma que esta membrana externa es permeable a todas las moléculas con un peso molecular menos de 5000 daltons, incluidas pequeñas proteínas. Estas moléculas pueden entonces penetrar en el espacio intermembranoso, pero la mayoría no puede atravesar la membrana interna. Otras proteínas de esta membrana son las enzimas implicadas en la síntesis mitocondrial de lípidos y las enzimas que transforman en la matriz los substratos lipídicos en forma metabolizables. En ella se lleva a cabo por tanto la síntesis de fosfolípidos y la desaturación y
elongación de los ácidos grasos.
El espacio intermembranoso contiene varias enzimas que utilizan la salida de ATP de la matriz para fosforilar otros nucleótidos. Su composición es muy similar a la del citosol en cuanto a moléculas de menos de 6000 daltons. Tienen muchas enzimas; destaca la adenilquinasa, una enzima muy importante, ya que cataliza la formación del ADPA a partir de
interna es altamente impermeable, hecho que provoca que no haya una libre circulación de moléculas a través de la membrana. Tanto el AMP con el Pi o el ADP provienen del citosol, que entran a través de una proteína. Pero esta solo permite el paso del ADP. Por eso la enzima adenilquinasa es útil para sintetizar el paso de AMP y Pi a ADP.
La matriz mitocondrial (semufluida) contiene una mezcla altamente concentrada de cientos de enzimas, incluyendo las necesarias para la oxidación del piruvato y los ácidos grasos y para el ciclo del ácido cítrico. También contiene diversas copias idénticas del genoma mitocondrial, ribosomas de mitocondrias (70S), tRNA, rRNA y varias enzimas necesarias para la expresión de los genes mitocondriales (enzimas que participan en la replicación, la transcripción y la traducción).
En cuanto al genoma mitocondrial : consiste en una molécula de DNA circular y pequeño (entre 15000 y 20000 pares de bases), con diversas copias en la matriz mitocondrial(en espermatozoides humanos unos pocos cientos de copias, en el ovocito unas 200000 copias; normalmente unas 1000 y 10000 copias por célula, con 2-10 moléculas de DNA por mitocondria). Corresponde a un 1% del DNA total en mamíferos.
Codifica para: unas 13 subunidades polipeptídicas de proteínas de la membrana interna (forman parte de complejos multienzimáticos del sistema OXPHOS), 2 rRNA (para las dos subunidades de 12S y de 16S del rRNA de la matriz mitocondrial) y 22 tRNA (requeridos para la síntesis proteica en la matriz mitocondrial).
Es DNA sin intrones. Además, en las mitocondrias existe un número más pequeño de tRNA que está en el citosol, lo que limita la cantidad de proteínas que se puedan sintetizar. Además, una gran cantidad de proteínas mitocondriales están codificadas por DNA del núcleo, que se sintetizan en los ribosomas citoplasmáticos y que son importadas posteriormente por la mitocondria. El código genético es diferente al universal, y el mitocondrial de diferentes especies también puede ser diferente.
La herencia es citoplasmática y materna, ya que la cola del espermatozoide, que es donde se encuentran las mitocondrias, no entra dentro del óvulo. Por tanto, el DNA de las mitocondrias es totalmente procedente del óvulo. Este genoma es útil en estudios porque la cantidad es mayor, no ha sufrido recombinación meiótica y la frecuencia de mutaciones es menor.
BIOGÉNESIS:
Es el crecimiento y la división de orgánulos ya existentes, se produce durante la interfase y tiene dos etapas:
- Crecimiento: durante el cual se produce un aumento de la membrana, re sintetizan proteínas propias y se lleva a cabo la importación de proteínas (el 95% del citosol), la importación de lípidos y se lleva a acabo la replicación del DNA mitocondrial.
- División: la mitocondria de divide en dos, suponiendo que es el proceso que se lleva a cabo en la fisión.
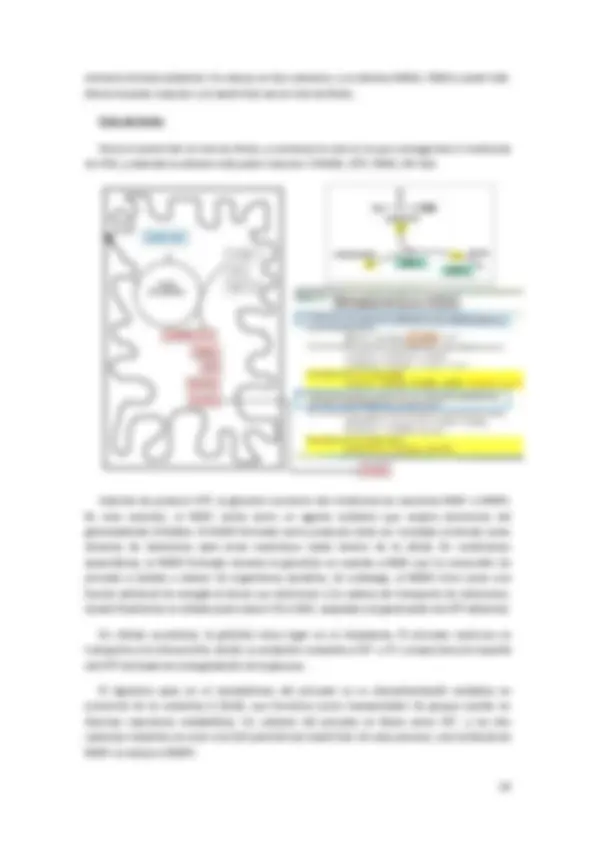
IMPORTACIÓN DE PROTEÍNAS:
A pesar que de las mitocondrias tienen su propio DNA y su maquinaria para la síntesis proteica, la mayoría de sus proteínas están codificadas en el núcleo celular y son importadas desde el citosol. Además, cada proteína importada ha de llegar al subcompartimento determinado en el que es funcional. Cada uno de los compartimentos tiene una colección de proteínas determinadas. Por tanto, el crecimiento de las mitocondrias mediante la importación de proteínas citoplasmáticas, constituye una proeza, que implica la translocación selectiva de estas proteínas a través de
A diferencia del direccionamiento hacia la matriz, el direccionamiento de proteínas al espacio intermembranoso, a la membrana interna y a la externa, suele requerir más de una secuencia señal y tiene lugar a través de una de varias vías posibles (cuadro).
A. Proteínas destinadas a la matriz.
Se lleva a cabo a través de los canales TOM y TIM 23, y se da en los lugares de contacto entre la membrana interna y la externa.
Lo primero es el reconocimiento de la proteína a transportar. Luego se dispone en el primer canal de translocación (TOM). A partir de aquí puede quedar en el espacio intermembranoso o llegar al TIM 23; que la proteína vaya hasta el TIM 23 es gracias al gradiente pH que hay, porque en el espacio intermembranoso hay muchos protones, y van entrando hacia el TIM 23 y de aquí a la matriz; la chaperonas situadas en la matriz son las que facilitan la entrada de la proteína hacia el interior. Una vez ha entrado, s elimina la secuencia señal y se repliega la proteína.
Las chaperonas Hsc70 citosólica se encarga de mantener las proteínas nacientes y recién sintetizadas en estado desplegado, para que puedan ser captadas por la mitocondria. La entra de un precursor mitocondrial desplegado se inicia con la unión de la secuencia específica de destino mitocondrial a un receptor de ingreso presente en la membrana externa de la mitocondria (por ejemplo como receptores TOM 20 Y TOM 22); luego estos receptores transfieren las proteínas a una canal de entra, un canal compuestos fundamentalmente de la proteína TOM 40, y se conoce como poro de entrada general ). TOM 40 forma un canal transmembrana con un poro amplio para que quepa la cadena polipeptídica desplegada. Es un canal fundamentalmente pasivo a través de la membrana externa y la energía motriz para el transporte unidireccional hacia la mitocondria proviene del interior de esta.
Cuando la proteína ha de pasar hasta la matriz, ha de darse el paso anterior en puntos de contacto de la MME y la MMI, para que el paso se continúe rápidamente a través del complejo de TIM 23 y TIM 17.
Después, la secuencia N-terminal es eliminada por la proteasa. La proteína que queda ahora es captada por la Hsc70 de la matriz (una chaperona que se encuentra en los canales de translocación y que interactúa con TIM 44); esta interacción estimula la hidrólisis de ATP por la Hsc70 de la matriz y se cree que estas dos proteínas impulsan en conjunto la translocación hacia la matriz.
Algunas proteína incorporadas pueden plegarse para constituir su estructura final activa sin ayuda adicional; pero el plegado final de muchas proteínas de la matriz requiere la acción de una chaperonina que facilita el plegado mediante un proceso dependiente de ATP. Por eso se necesita un triple aporte de energía para incorporar proteínas a la mitocondria: para el gradiente de protones del espacio intermembranoso, las chaperonas citosólicas y las Chaperoninas mitocondriales.
B. Proteínas destinadas a la membrana mitocondrial interna.
Hay tres tipos de mecanismos de transporte: A, B y C. las proteínas que entran por el mecanismo A y B tienen secuencia señal, pero las de la C no. Además de la secuencia señal encontramos secuencias hidrofóbicas adicionales, que detendrá la translocación de la proteína para que quede a la membrana.
A: la secuencia hidrofóbica actúa de detención de la translocación y la proteína queda insertada en la membrana como una hélice alfa. La proteasa de la matriz elimina la secuencia señal.
C. Proteínas destinadas al espacio intermembranoso.
Existen dos vías que llevan proteínas citosólicas al espacio entre la membranas mitocondriales interna y externa.
Vía A: la vía principal la hacen las proteínas cuyos precursores poseen dos secuencias de direccionamiento diferentes en el extremo N-terminal. La secuencia más cercana al N- terminal es una secuencia de direccionamiento a la matriz que es eliminada por la proteasa de la matriz. La segunda secuencia de direccionamiento es un segmento hidrófobo que bloquea la translocación completa de la proteína a través de la membrana interna. Después que el intermediario resultante incluido en la membrana se difunde lateralmente apartándose del canal de translocación Tim23/17, una proteasa de la membrana corta la proteína cerca del segmento transmembrana hidrófobo y libera la proteína madura en forma soluble en el espacio intermembranoso.
Vía B: en esta vía, la proteína introducida se dirige directamente al espacio intermembranoso a través del poro de entrada general TOM40 pasivamente y queda atrapada en el espacio intermembranoso.
D. Proteína destinadas a la membrana mitocondrial externa: para que una proteína sea dirigida a la membrana externa, debe poseer una corta secuencia de direccionamiento a la matriz en N-terminal, a la que sigue un largo tramo de aa hidrofóbicos. Esta secuencia hidrofóbica larga funciona como una secuencia de detención de la translocación que impide que la proteína entre hacia el espacio intermembranoso y la ancla en la membrana como proteína integral. Normalmente la secuencia señal no es cortada.
FUNCIONES DE LA MITOCONDRIA
En cuanto a las funciones mitocondriales, estas tienen lugar en membranas específicas o dentro de los compartimentos. La mayoría de los intermediarios de la cadena de transporte de electrones son componentes integrales de la membrana interna, donde se encuentran organizados formando grandes complejos. Sobresaliendo desde esa membrana interna hacia la matriz se encuentran unas esferas con forma de puño que se llaman complejos F1; cada complejo consta de seis polipéptido ensamblado; estos complejos tienen un diámetro aproximado de 9nm y son abundantes en las crestas. Cada uno está unido mediante un tallo proteíco a un complejo F0, un conjunto ensamblado de proteínas hidrofóbicas que se encuentra insertado en la membrana interna. La asociación de un complejo F1 y otro F0 se conoce como complejo F1F0 y se considera como una ATPsintetasa, porque esa es su función habitual en el metabolismo energético. El complejo F1F0 es de hecho el responsable de la mayor parte de la producción de ATP que tiene lugar en la mitocondria, así como también en las células procariotas y en los cloroplastos. En todos los casos, la producción de ATO por estos complejos está dirigida por un gradiente electroquímico de protones a ambos lados de la membrana en la que dichos complejos están anclados.
Para que los procariotas puedan realizar la respiración aeróbica, se sabe que el citoplasma y la membrana plasmática de una célula procariota realizan las mismas funciones respectivamente, que la matriz mitocondrial y la membrana interna. En las células procariotas, por tanto, la mayoría de enzimas de la glucolisis, del ciclo de Krebs y del catabolismo de ácidos grasos y aminoácidos se encuentran en el citoplasma, mientras que las proteínas de transporte de electrones está en la membrana plasmática, el igual que el complejo F1F0, sobresaliendo el F1 hasta el citoplasma.
procedente del metabolismo de la glucosa. Esto implica la oxidación inicial del piruvato a acetil CoA, que posteriormente es degradado hasta CO2 a través del ciclo de Krebs. La oxidación de los ácidos grasos también produce acetil CoA, que de forma similar es metabolizado por el ciclo del ácido cítrico (ciclo de Krebs) en las mitocondrias. Por lo tanto, las enzimas del ciclo de Krebs (localizadas en la matriz mitocondrial), tienen un papel central en la degradación oxidativa tanto de los carbohidratos como de los ácidos grasos.
La oxidación del acetil CoA a CO2 está acoplada a la reducción de Nad+ y Fad a Nadh y FADH, respectivamente.
Por lo tanto, la mayor parte de la energía derivada del metabolismo oxidativo es producida por el proceso de la cadena de electrones, que tiene lugar en la membrana mitocondrial interna. Los electrones de alta energía del NADH y FADH2 se transfieren al oxigeno molecular a través de una serie de transportadores de la membrana. La energía derivada de estas reacciones de transferencia de electrones se convierte en energía potencial acumulada en forma de un gradiente de protones a través de la membrana, que es utilizada para dirigir la síntesis de ATP. La membrana interna mitocondrial representa de esta manera el lugar principal de generación de ATP, y este papel fundamental se refleja en su estructura. Primero, el incremento de su superficie mediante su plegamiento en crestas. Además, la membrana interna mitocondrial contiene una proporción inhabitualmente elevada (más del 70%) de proteínas, que intervienen en la cadena de electrones así como en el transporte de metabolitos (p.ej. piruvato y ácidos grasos) entre el citosol y la mitocondria. Por otra parte, la membrana interna es impermeable a la mayoría de los iones y de las moléculas pequeñas- una propiedad crítica para mantener el gradiente de protones que dirige la fosforilación oxidativa.
A diferencia de la membrana interna, la membrana externa mitocondrial es completamente permeable a las moléculas pequeñas. Esto es debido a que contiene numerosas porinas. La membrana mitocondrial interna es la barrera funcional para el paso de moléculas pequeñas entre el citosol y la matriz y mantiene el gradiente de protones que dirige la fosforilación oxidativa.
Patata brava: D’aquest procés generem un gradient de protons del qual obtindrem un infinit de funcions.
- Oxidació d’àcids grassos i del piruvat
- Cadena respiratòria
- Síntesi d’ATP
- Cicle de Krebs
Los carbohidratos se degradan mediante la glicólisis en el citosol de la célula, del cual se obtiene 2 NADH, 2 piruvatos y 2 ATP. Este piruvato entra el interior de la mitocondria. Para entrar necesita transportadores específicos, ya que la membrana de la mitocndria es muy impermeable. La piruvato deshidrogenasa le degrada a dos molédulas de acetil-CoA y obtiene así poder reductor (NADH).
Los ácidos grasos se encuentran en forma de Triacilglicéridos en las células adiposas. El ácido graso se combina con la coenzima CoA y entrará mediante una proteína. Este ácido graso
entrará a la beta oxidación. Se reduce en dos carbonos, y se obtiene NADH, FADH y acetil-CoA. Ahora el poder reductor y el acetil-CoA van al ciclo de Krebs.
Ciclo de Krebs
Entra el acetil-CoA al ciclo de Krebs, y comienza la ruta en la que conseguimos 2 moléculas de CO2, y además se obtiene más poder reductor 3 NADH, GTP, FADH, HS-CoA.
Además de producir ATP, la glicolisis convierte dos moléculas de coenzima NAD+ a NADH. En esta reacción, el NAD+ actúa como un agente oxidante que acepta electrones del gliceraldehido-3-fosfato. El NADH formado como producto debe ser reciclado sirviendo como donante de electrones para otras reacciones redox dentro de la célula. En condiciones anaeróbicas, el NADH formado durante la glucolisis se reoxida a NAD+ por la conversión de piruvato a lactato o etanol. En organismos aerobios, sin embargo, el NADH sirve como una fuente adicional de energía al donar sus electrones a la cadena de transporte de electrones, donde finalmente se utilizan para reducir O2 a H2O, acoplado a la generación de ATP adicional.
En células eucariotas, la glicolisis tiene lugar en el citoplasma. El piruvato entonces se transporta a la mitocondria, donde su oxidación completa a CO” y H”= proporciona la mayoría del ATP derivado de la degradación de la glucosa.
El siguiente paso en el metabolismo del piruvato es su descarboxilación oxidativa en presencia de la coenzima A (CoA), que funciona como transportador de grupos acetilo en diversas reacciones metabólicas. Un carbono del piruvato se libera como CO”, y los dos carbonos restantes se unen a la CoA para formar acetil-CoA. En este proceso, una molécula de NAD+ se reduce a NADH.
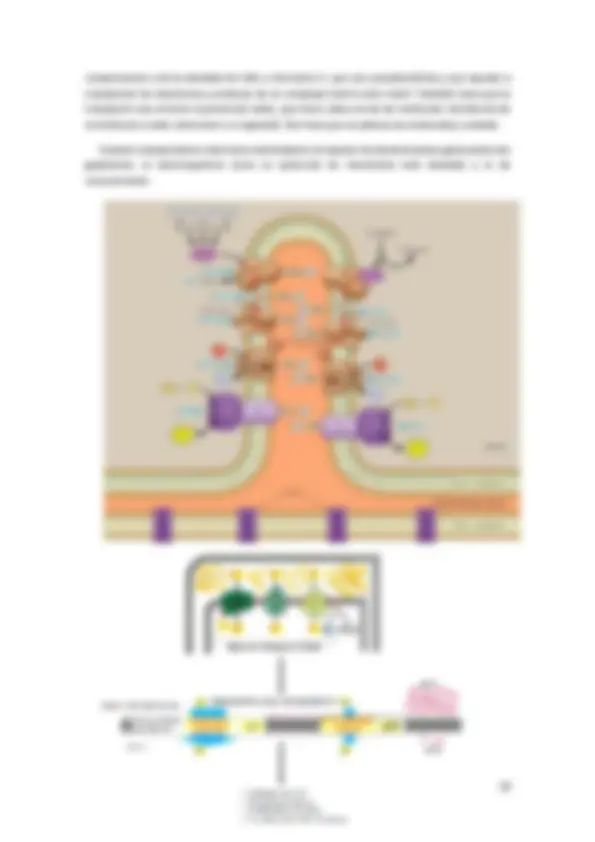
La cadena respiratoria se encuentra en la membrana interna de la mitocondria, formada
por 4 complejos multiproteicas (no móviles) i 2 pequeños (móviles).
- 4 complexos multiproteicos (a la imagen es ven 3). Tienen grupos prostéticos que se
encargarán de transportar los electrones.
I: NADH deshidrogenasa
II: succinat-COQreductasa
III: citocrom BC
IV: Citocrom oxiadasa
- Tenemos 2 componentes móviles:
Ubiquinona: con una cola lipídica que puede estar insertada pero se puede
mover.
Citocrom C: situado a la zona del espacio intramembranoso, hace
interacciones iónicas.
Las diferentes interacciones entre ellos permiten que los electrones circulen per la cadena
y permiten el bombeo de protones desde el espacio intramembranoso.
Grups prostètics: grupos que no son proteína que es por donde se transportan electrones
o protones:
a. Flavoproteïnes: en el transporte de electrones participan varias Flavoproteínas unidas a membrana, que usan como grupo prostético, bien FAD, bien FMN. Un ejemplo de flavoproteína es la NADH deshidrogenasa, que forma parte del complejo proteico que acepta los pares de electrones que provienen del NADH. Otro ejemplo es la enzima succinato deshidrogenasa que tiene al FAD como grupo prostético y forma parte de un complejo respiratorio unido a membrana, que acepta los pares del succinato a través de FAD. Una característica importante, es que transfieren tanto protones como electrones. b. Ferrosulfoproteínas: también llamadas ferredoxinas, forman parte de una familia de proteínas, cada una con un centro ferrosulfuroso (Fe- S), que consiste en átomos de hierro y azufre asociados con los grupos cisteína de la proteína. Los átomos de hierro de estos centros activos son los verdaderos transportadores. Cada átomo de hierro alterna entre la forma oxidada (Fe3+) y la reducida (Fe2+) durante el transporte de electrones, que en este caso afecta a un único electrón.
c. Citocroms: Al igual que las Ferrosulfoproteínas, los citocromos también contienen hierro, pero formando parte del grupo prostético hemo. Existen al menos cinco tipos de citocromos en la cadena de transporte de electrones: b, c, c1, a y a3. El átomo de Fe del grupo prostético, al igual que el del centro en las ferredoxinas, es el transportador de electrones (de uno en uno) y sin transportar H+, mientras que los citocromos c1, b, a y a3 son proteínas integrales de membrana, el citocromo c es una proteína periférica de membrana, asociada a la cara externa de la membrana mitocondrial interna y por tanto puede difundir mucho más rápido, una propiedad clave para desempeñar su papel en la transferencia de electrones entre complejos proteicos. d. Citocroms amb coure: además de sus átomos de hierro, los citocromos a y a también contienen un átomo de cobre unido al grupo hemo del citocromo, donde se encuentra asociado con un átomo de hierro para formar un centro bimetálico hierrocobre (Fe-Cu). De la misma manera que los átomos de hierro, el cobre también puede convertirse reversiblemente de la forma oxidada (Cu2+) a la forma reducida Cu3+. e. Ubiquinona: La coenzima Q (CoQ) el único componente no proteíco de la cadena transportadora de electrones. La ubicuinona tiene la capacidad de reducirse reversiblemente en dos pasos de un electrón, desde la forma quinona (CoQ) a la forma dihidroquinona (CoQH2), pasando antes por la forma semiquinona (CoQH). La ubicuinona, no forma parte de ningún complejo respiratorio, sino que se encuentra en el interior apolar de la membrana mitocondrial interna. Las ubicuinonas, son los transportadores de electrones más abundantes en la membrana, sirviendo como un punto de recolección de electrones desde los grupos prostéticos de las deshidrogenasas unidas al FMN o al FAD en la membrana.
componentes y de la cantidad de CoQ y citocromo C, que son pseudomóviles y que ayudan a transportar los electrones y protones de un complejo móvil a otro móvil. También hace que el transporte sea correcto el potencial redox, que tiene cada una de las moléculas (tendencia de la molécula a ceder electrones o a captarlo). Eso hace que la cadena sea ordenada y cuidada.
Cuando transportamos electrones bombeamos al espacio intramembranoso generando dos gradientes: el electroquímico (crea un potencial de membrana más elevado) y el de concentración.
Resumen del metabolismo energético mitocondrial.
El piruvato y los ácidos grasos que entran en la mitocondria son procesados a acetil CoA y son metabolizados por el ciclo de Krebs, produciéndose NADH y FADH2.
En el proceso de la fosforilación oxidativa, los electrones de alta energía de NADH y FADH se transfieren al oxigeno mediante la cadena respiratoria de la membrana mitocondrial interna, produciéndose ATP mediante un mecanismo quimiosmótico.
El NADH generado en el citosol por la glucólisis también transfiere electrones a la cadena respiratoria. Dado que el NADH no puede pasar a través de la membrana mitocondrial interna, la transferencia electrónica desde el NADH, la transferencia electrónica se ha de efectuar de manera indirecta por medio de un sistema “lanzadera”, que transporta otros compuestos reducidos al interior de la mitocondria; después de ser oxidado, este compuesto vuelve al citosol, donde es reducido de nuevo por el NADH.
Según la hipótesis quimiosmótica, los intermediarios químicos de alta energía son substituidos por una conexión entre procesos químicos (“quimio”) y procesos de transporte (“osmótico”, del griego osmos, empujar) – De ahí el nombre de acoplamiento quimiosmótico. Cuando los electrones de alta energía de los hidrógenos del NADH y del FADH2 son transferidos a lo largo
de la cadena respiratoria de la membrana mitocondrial interna, la energía que se libera cada vez que pasan de una molécula transportadora a la siguiente es utilizada para bombear protones (H+) a través de la membrana interna, desde la matriz mitocondrial hasta el espacio intermembranoso.