¡Descarga embriologia y más Apuntes en PDF de Anatomía solo en Docsity!
Tema 1: De la fecundación a la
implantación
Estudia el desarrollo prenatal (embriones y fetos). Se puede dividir en:
- Fecundación
- Periodo de división celular: 1 y 2 semana, es el periodo de implantación del blastocisto en mucosa uterina
- Periodo embrionario semanas de 2 a 8 es el período de máximo desarrollo orgánico
- Periodo fetal: 9 a 38-40: comprende los 7 últimos meses. Es un periodo de crecimiento y de desarrollo de los órganos.
Gametogénesis:
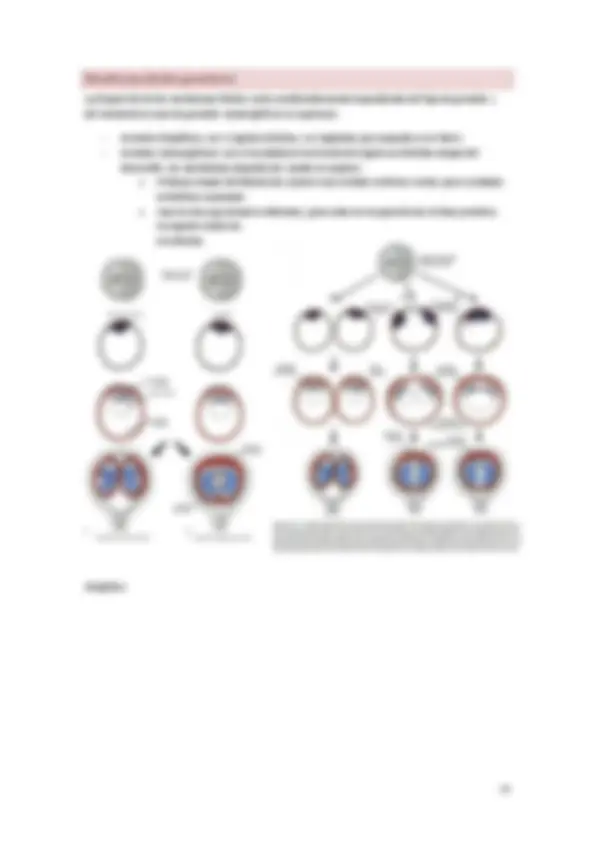
El desarrollo humano comienza con la fecundación, que es la unión de espermatozoide y ovocito para formar el cigoto. Las células germinativas experimentan una serie de cambios (cromosómicos y citoplasmáticos) preparatorios para la fecundación.
La fecundación de la gametogénesis es:
- Reducir cromosomas de número diploide (23 pares) a haploide por meiosis.
- Modificar la forma de las células germinativas para que se conviertan en gametos.
La meiosis transcurre en 2 etapas:
- 1ª: en ella se da la replicación del ADN, el apareamiento de los cromosomas homólogos (sinapsis) y el entrecruzamiento, mediante el cual se intercambia material genético.
- 2ª: en ella obtenemos 4 células haploides de 23 n.
Los gametos derivan de las células germinales primordiales son las que se forman en el epiblasto durante la 2ª semana del desarrollo y se desplazan hacia la pared del saco vitelino. Durante la cuarta semana del desarrollo se desplazan hacia las gónadas, a las que llegan hacia el final de la quinta semana. Hasta aquí no hay proceso de diferenciación.
La gametogénesis puede ser:
- Espermatogénesis: proceso de producción de espermatozoides maduros (gametos masculinos). Van de espermatogonias espermatozoides
- Ovogénesis: proceso de maduración de ovocitos maduros (gametos femeninos). Va de ovogonias a ovocitos maduros.
Espermatogonias y ovogonias proceden de las células primordiales que se diferencian.
Espermatogénesis:
Es el proceso de formación de los espermatozoides maduros a partir de las espermatogonias. Comienza en la pubertad y se produce en los túbulos seminíferos.
Las espermatogonias se diferencian en espermatocitos primarios. Las espermatogonias y los espermatocitos primarios son células diploides.Las células de sostén o de Sertoli se encargan de cuidar de los gametos y derivan del epitelio superficial de la glándula.
Este proceso ocurre en la pubertad que es cuando se transforman en espermatocitos primarios en los túbulos seminíferos. En el que van evolucionando hacia la luz del túbulo.
Los espermatocitos primarios entran en la primera división meiótica y se forman los espermatocitos secundarios, que se convierten en haploides. Los espermatocitos secundarios experimentan uuna 2ª división meiótica para lugar a cuatro espermátidas haploides.
El proceso continúa con la espermiogénesis. Los cambios que se producen son:
- Formación del acrosoma
- Condensación del núcleo
- Formación del cuello, pieza intermedia (mitocondrias) y cola
- Eliminación de la mayor parte del citoplasma por parte de las células de Sertoli.
El proceso dura aproximadamente 74 días y cada día se producen 300 millones de espermatozoides. De una espermátida salen 4 gametos haploides iguales y funcionales.
La espermatogénesis está regulada por la producción de LH de la hipófisis y estimula a las células de Leydig y estimula la producción de testosterona, que se unen a las células de Sertoli y estimula la espermatogénesis.
La FSH se une a las células de Sertoli y estimula la producción de líquido testicular y la síntesis de proteínas receptoras de andrógeno intracelular.
Ovogénesis:
Es el proceso de formación de los ovocitos maduros. Comienza antes del nacimiento.
Una vez que las células germinales primordiales han alcanzado la gónada se diferencia en ovogonias. Las ovogonias proliferan por mitosis.
Alrededor del tercer mes algunas ovogonias dan origen a ovocitos primarios que comienzan la profase de la primera división meiótica. A medida que se forma el ovocito primario se rodea de una sola capa de células epiteliales foliculares. El ovocito rodeado por esta capa celular se denomina folículo primordial.
Hacia el 5º mes de desarrollo intrauterino alcanzan su nº máximos, unos 7 millones. En este momento comienza la muerte celular y muchas ovogonias y ovocitos 1os se vuelven atrésicos.
Alrededor del 7º mes la mayor parte de las ovogonias han degenerados excepto las próximas a la superficie.
Al nacimiento todos los ovocitos primarios ha comenzado la profase de la meiosis I pero no continua con la metafase sin que entran en fase de diploteno de la profase. Esta etapa de reposo dura hasta la pubertad, en la cual solo existen alrededor de 40.000 ovocitos y solo aproximadamente 500 llegarán a ser ovulados.
En la pubertad, los ovocitos 1 terminan de dividirse y forman los ovocitos secundarios (reciben la mayor parte del citoplasma) y un corpúsculo polar (casi no recibe citoplasma).
A continuación el ovocito secundario entre la meiosis 2 pero se detiene en metafase unas 3 horas antes de la ovulación. Si no hay fecundación el ovocito no madura más, ya que necesita la fecundación para convertirse en óvulo y un 2º corpúsculo polar.
Al final del proceso obtenemos un óvulo funcional y 3 corpúsculos polares.
Las diferencias entre un proceso y otro:
- Ovogénesis es un proceso parcial (por etapas) mientras que en el hombre es continúa
- La ovogénesis es incompleta mientras que el masculino es completa
- La ovogénesis es asimétrica ( un óvulo funcional) y la espermatogénesis (4 células iguales)
- Se inicia antes del nacimiento mientras que la masculina se inicia en la pubertad.
Ciclo ovárico y uterino:
Ciclo ovárico:
Cuando llegamos a la pubertad y durante todos los ciclos maduran los folículos primordiales. A esto se le conoce como ciclo ovárico y en el distinguimos:
- Fase folicular
- Ovulación
- Fase lútea
Fase folicular:
En la pubertad comienzan los ciclos menstruales regulares en la mujer que preparan al aparato reproductor para el embarazo. El hipotálamo (células neurosecretoras) sintetiza la hormona liberadora de gonadotropinas (GnRH) que se transporta hasta la hipófisis (lóbulo anterior o adenohipófisis). La GnRH estimula la liberación de FSH /hormona folículo estimulante) y LH (hormona luteinizante). El órgano diana es el ovario.
- FSH: estimula el desarrollo de los folículos ováricos y la producción de estrógenos por las células foliculares, que producen estrógenos.
- LH: estimula la ovulación y las células foliculares y el cuerpo lúteo para que produzca progesterona.
Ambas inducen el crecimiento del endometrio.
Al comienzo del ciclo la FSH estimula el crecimiento de 5 a 10 folículos primordiales. Sólo uno de ellos (en condiciones normales) evoluciona a un folículo maduro y es expulsado únicamente un ovocito; los demás degeneran y se convierten en folículos atrésicos.
El desarrollo de un folículo ovárico se caracteriza por:
El ovocito primario comienza a crecer y las células foliculares que le rodean cambian de forma plana a cúbica y luego proliferan formando un epitelio estratificado de células de la granulosa. Este conjunto se denomina folículo primario.
Las células de la granulosa asientan sobre una membrana basal que las separa de las células de estroma circundante que constituyen la teca folicular.
Fase lútea:
Después de la ovulación las células de la granulosa que quedan en la pared del folículo roto, junto con las células de la teca interna, se vascularizan por los vasos sanguíneos que la rodean. Por efecto de la LH se convierten en células luteínicas que forman el cuerpo lúteo o cuerpo amarillo y secreta progesterona. La progesterona y los estrógenos determinan que la mucosa uterina alcance la fase progestacional, luteínica o estado secretor como preparación para la implantación del embrión.
Si el ovocito no es fecundado, el cuerpo lúteo involuciona y se degenera en los próximos días y transforma en un cuerpo cicatricial blanco o cuerpo albicans. Al mismo tiempo disminuye la producción de progesterona, lo que desencadena la hemorragia menstrual.
Si el ovocito es fecundado, la gonadotropina coriónica humana (hCG), hormona se secreta en el sincitiotrofoblasto del embrión en desarrollo, impide la degeneración del cuerpo lúteo, que sigue creciendo y forma el cuerpo lúteo del embarazo (cuerpo lúteo gravídico). Este cuerpo lúteo gravídico conserva su función durante las primeras 20 semanas y después experimenta una regresión lenta a medida que la secreción de progesterona es asumida por la placenta.
Ciclo uterino:
La pared del útero esta formada por 3 capas, que son:
- Endometrio: mucosa que reviste la pared interna. Experimenta cambios ciclícos cada 28 días, en los que pasa por 3 fases: o Fase folicular o proliferativa: está regulada por los estrógenos y se corresponde con el crecimiento de los folículos ováricos. Esta irrigado por sus propias arterias (arterias basales), funciona como capa regeneradora y reconstruye la glándulas y las arterias. o Secretora o progestacional: en respuesta a la progesterona producida por el cuerpo lúteo, la fase secretora se inicia 2 o 3 días después de la ovulación. o Menstrual: comienza si no tiene lugar la fecundación, el endometrio se desprende. Se van compactando con las células sanguíneas y la sangre escapa de las arterias superficiales y se desprenden pequeños fragmentos de estroma y glándulas. Los estratos esponjoso y compacto son expulsados del útero, siendo el estrato basal la única parte del endometrio.
- Miometrio: capa gruesa de músculo liso
- Perimetrio o revestimiento peritoneal de la pared externa.
Si hay fecundación el endometrio ayuda a la implantación y contribuye a la formación de la placenta, que asume el papel productor de hormonas y el cuerpo lúteo degenera.
En el momento de la implantación, la mucosa del útero se encuentra en la fase secretora y durante ese tiempo las arterias y las glándulas uterinas se desarrollan. En el tejido se reconocen 3 capas en el endometrio:
- Estrato compacto superficial
- Estrato esponjoso intermedio
- Estrato basal delgado
Fecundación:
Tiene lugar en la ampolla de la trompa de Falopio. Solo el 1% de los espermatozoides depositados en la vagina entran en el cuello del útero, de ahí se desplazan hasta la trompa de Falopio gracias a su propia propulsión y a las contracciones musculares del útero y una vez en el istmo pierden movilidad. El viaje dura entre 2 y 7horas.
En el momento de la ovulación, los espermatozoides recuperan la movilidad, en parte por los quimioatrayentes del óvulo. Antes de fecundar al ovocito tiene que pasar por la captación y la reacción acrosómica:
- Captación: es un periodo de acondicionamiento en la trompa de Falopio que consiste en interacciones epiteliales entre los espermatozoides y la mucosa de la trompa. Durante este proceso se eliminan la capa de glucoproteínas y las porteínas seminales de la membrana plasmática que recubre la acrosómica de los espermatozoides. Solo los capacitados pueden atravesar la corona y experimentar la reacción acrosómica.
Segmentación:
El cigoto ha llegado a la fase de dos células, experimenta una serie de divisiones mitóticas, cuyo tamaño se reduce con cada división y recibe el nombre de blastómeros.
Hasta la fase de 8 células forman un grupo laxo y después de la tercera división los blastómeros maximizan el contacto entre ellos y forman una pelota compacta de células que se mantienen juntas con uniones herméticas. El proceso de compactación separa las células internas.
3 días después de la fecundación, las células del embrión compactado se dividen de nuevo y forman una mórula de 16 células (mora).
Las células internas de la mórula constituyen la masa celular interna y las células que las rodean componen la masa celular externa. La masa celular interna origina los tejidos propios del embrión, y la masa celular externa forma el trofoblasto, que más adelante formará la placenta.
Formación del blastocisto:
4 días después de la fecundación comienza a penetrar líquido entre las células, mórula a través de la membrana pelúcida. Este líquido forma el blastocele o cavidad y a partir de aquí el embrión se conoce como embrioblasto.
Conforme se llena la cavidad de líquido, los blastómeros se separan en dos grupos:
- Una capa celular externa delgada: trofoblasto
- Un grupo central: embrioblasto
El trofoblasto dará lugar a la placenta mientras que el embrioblasto dará lugar al embrión. Durante dos días el blastocisto permanece en la superficie uterina y la membrana pelúcida desaparece. El embrioblasto se sitúa en un polo del blastocisto. El polo embrionario es el polo animal.
Implantación:
6 días después de la fecundación el embrión comienza a implantarse en el endometrio por el polo animal o embrionario. El trofoblasto comienza a proliferar rápidamente en cuanto se adhiere al endometrio diferenciándose en dos capas el citotrofoblasto (con membrana) y el sincitiotrofoblasto (capa más cercana al útero). Aprovechan las cavidades para implantarse en la cara anterior o posterior, en la mayoría de los casos.
A los 6 días el sincitiotrofoblasto invade el tejido conjuntivo de la lámina propia del endometrio y el embrión se nutre de los tejidos maternos invadidos.
A los 7 días con el embrión ya implantado se diferencian un grupo de células aplanadas en el embrioblasto en la superficie en contacto con el blastocele: el hipoblasto o endodero primitivo. El epiblasto la que queda por encima. Se cierra con un coágulo de fibrina.
La gonadotropina coriónica es la que se detecta en las tiras de orina y se segrega en el sincitiotrofoblasto.
Alteraciones:
- Abortos espontáneos: Se dan hasta el 45% de los abortos espontáneos. Esta tasa es el resultado de: o Alteraciones en la concentración de las hormonas del cuerpo lúteo o Anomalías cromosómicas fetales (>50%) o Otras causas
El diagnóstico in vitro de anomalías puede hacerse a los 3 o 5 días de la fecundación.
- La fecundación ectópica es una implantación fuera del útero, que es una urgencia quirúrgica.
Tema 3: Formación del disco bilaminar
Es la segunda semana.
La implantación se completa en la segunda semana. A medida que esto ocurre, el embrioblasto se transforma en un disco con dos capas celulares: epiblasto e hipoblasto. También forman la cavidad amniótica y el amnios, el saco vitelino (vesícula umbilical), el tallo de conexión y el saco coriónico.
Finalización de la implantación:
Las células conjuntivas del lugar de la implantación experimentan una reacción decidual: se hacen poliédricas y acumulan glucógeno y lípidos (células deciduales). Algunas células de este tejido degeneran y son engullidas por el sincitiotrofoblasto constituyendo una fuente de nutrición embrionaria. El sincitiotrofoblasto produce beta- HCG que es la base de las pruebas de embarazo. Hacia la segunda semana ya hay cantidad suficiente para positivar la prueba.
Entre los días 8 y 9 el sincitiotrofoblasto erosiona el tejido conjuntivo endometrial formando lagunas que se llenan de una mezcla de: sangre materna, procedente de los capilares rotos y restos celulares del endometrio lesionado. Lo que esta relacionado con el crecimiento del embrión es el embriotrofo. Hasta el día 12 esta el coagulo de fibrina y después se reepitelizará.
Las células que cubren la cavidad es el amnioblasto y la cavidad es el amnios. Que se comunica con el saco vitelino primitivo mediante el pedículo de fijación.
El líquido de las lagunas (embriotrofo) nutre al embrión por difusión. La comunicación entre los espacio lagunares y los capilares erosionados establece la circulación útero-placentaria primitiva.
Las sangre oxigenada es aportada por las arterias espiraladas del endometrio y la desoxigenada es retirada por las venas endometriales. La implantación y la anidación se completa totalmente en el día
El lugar de entrada del embrión al endometrio es taponado en un principio por un tapón de cierre formado por un coágulo fibroso que es sustituido hacia el día 12 por epitelio uterino regenerado. Hacia el día 12 las lagunas se fusionan formando redes lagunares que serán los precursores de los espacios intervellosos placentarios.
Desarrollo del disco embrionario:
Entre los 11 y 12 días algunas células que rodean el saco vitelino se diferencian y forman una capa entre el citotrofoblasto y las células de revestimiento de la cavidad exocelómica que constituyen el mesodermo embrionario que rodea a todo el embrión.
Poco después se forman cavidades en el mesodermo extraembrionario que van extrusionando otras zonas y formándose la cavidad amniótica definitiva. Si al extrusionar ciertas zonas dejan un quiste, hablaremos de quiste exocelómico.
Distinguimos ahora un pedículo de fijación (que dará el cordón umbilical) o lámina coriónica, cavidad amniótica, saco vitelino y esplacno o somatopleura. Queda todo rodeado de cavidad coriónica y sincitiotrofoblasto.
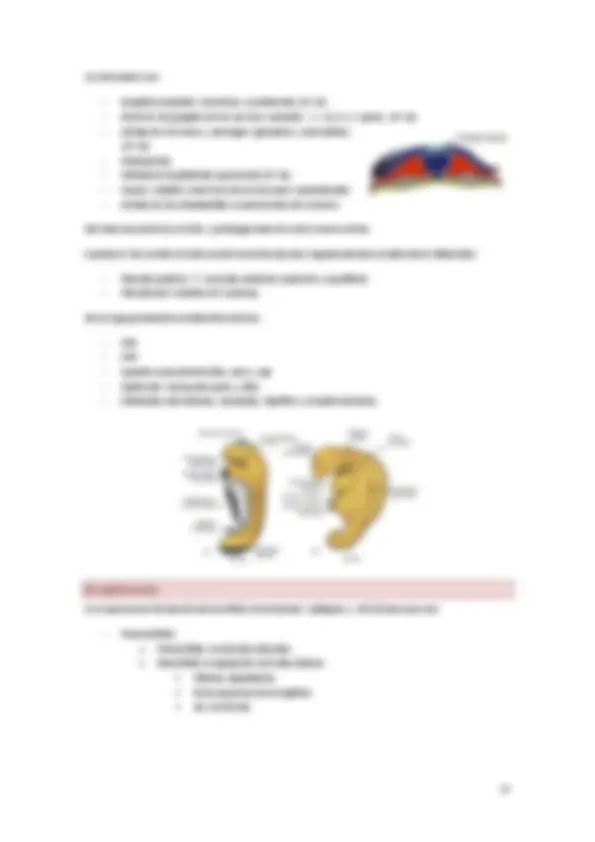
Hacia el día 14 el embrión tiene forma de disco bilaminar plano, Algunas células del hipoblasto de una zona concreta se hacen cilíndricas y forman un área circular engrosada la placa precordal que es el lugar donde se localizara la boca y representa una guía organizativa para el embrión.
Diferencia de la capas:
Prolongación cefálica y notocorda:
La notocorda es un eje real, rígido que define la simetría del individuo y que induce la diferenciación, primero la del ectodermo y también la del mesodermo.
La prolongación cefálica o notocordal es parte del mesodermo dorsal y se forma por las células que emigran desde la fosa primitiva hacia la porción cefálica del embrión hasta la placa precordal o membrana bucofaríngea.
La prolongación cefálica adquiere luz (conducto notocordal. El suelo de la prolongación notocordal se fusiona con el endodermo. Esta fusión provoca aberturas en el suelo de la prolongación por lo que el conducto notocordal se pone en contacto con el epiblasto.
Las aberturas confluyen y desaparece el suelo del conducto notocordal, quedando unidas las cavidades amniótica y vitelina a través del conducto neurentérico. Los resto de la prolongación notocordal forma la placa notocordal.
En el extremo craneal la placa comienza a plegarse para forma la notocorda. Cuando se forma la notocorda se cierra el conducto neurentérico. La notocorda se separa del endodermo, quedando de nuevo como una capa continúa. La notocorda se considera un inductor del desarrollo del sistema nervioso. No se habla de notocorda hasta que el tubo comienza a cerrarse.
La membrana cloacal se forma en el extremo caudal del disco. Es similar a la bucofaríngea y está formada por células ecto y endodérmicas. En la pared posterior se forma un divertículo que es el alantoides y aparece en el día 16.
La notocorda es un inductor primario y define el eje longitudinal del embrión, le da rigidez, es inductor para el desarrollo del SNC y las estructuras paraaxiales musculoesqueléticas y participa en la formación de los discos intervertebrales.
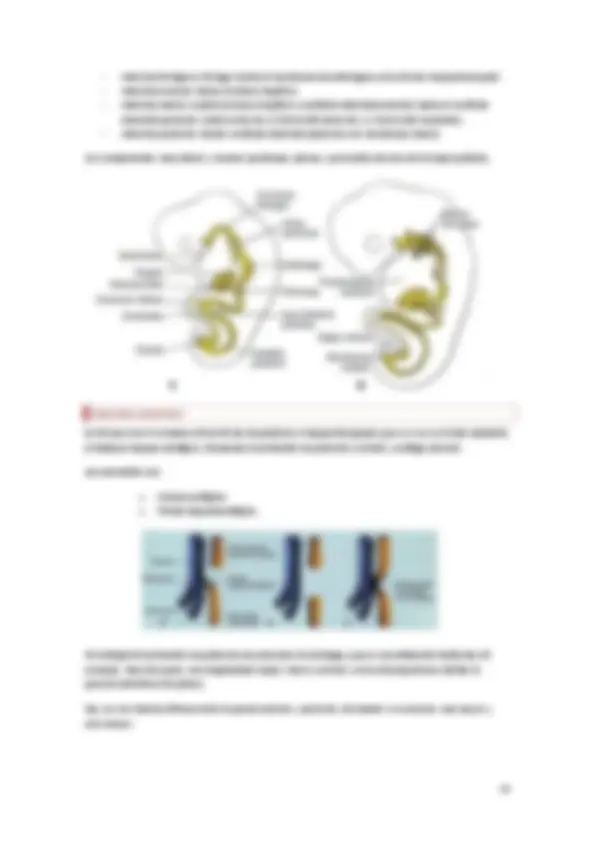
Desarrollo del mesodermo :
El mesodermo a lo largo del eje se divide en 3 partes. Uno que queda junto a la notocorda y que se conoce como paraaxial. Otro lateral y en contacto con el ectodermo que se conoce como somático. El intermedio da lugar a las vísceras.
El disco se empieza a plegar y las localizaciones cambian, por lo que el mesodermo paraaxial pasa a posición lateral y parece que sube, pero no migran las células.
Entre la somática y la esplánica se forma una cavidad que es la celómica, que da lugar a las cavidades en el adulto.
Mesodermo paraaxial:
De la 3 a la 8ª semana es la organogénesis.
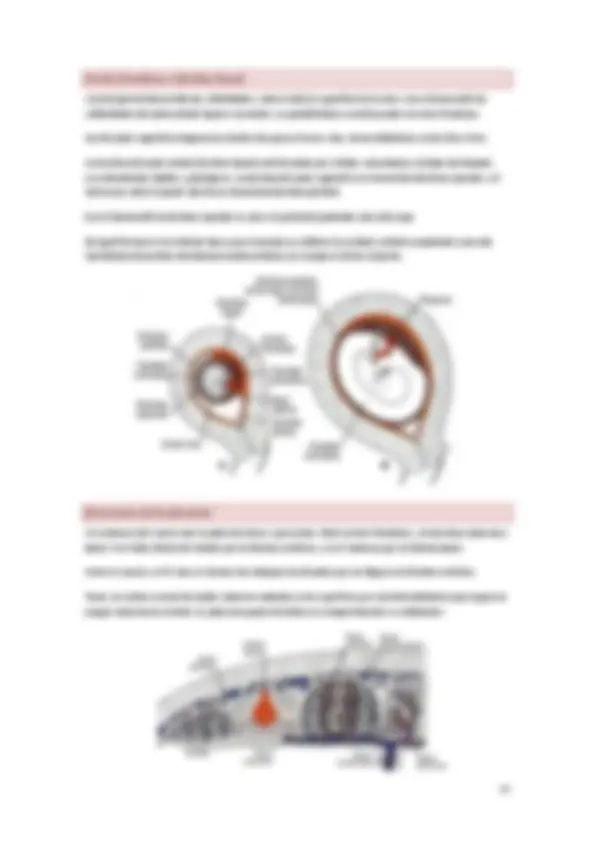
Al final de la tercera semana el mesodermo paraaxial se condensa y fusiona en elementos cuboides denominados somitómeras. Las somitómeras desde la zona occipital continúan condensándose formando las somitas, que se encuentran emparejados a cada lado del tubo neural en el desarrollo.
El primer par de somitas se desarrolla en la región occipital del embrión al final de la tercera semana. El ritmo de aparición es 2 a 3 pares por día. Del día al 20 al 30 aparecen 38 pares de somitas y a finales de la 5ª semana hay entre 42 y 44 pares originándose en un sentido craneocaudal.
Las somitas crean elevaciones en el embrión visto desde su porción dorsal: nº de somitas se toma como criterio para determinar la edad del embrión.
Las somitómeras van de la zona cervical a caudal y formaran la musculatura cráneo y cara. Las somitas evolucionan desde somitómeras occipitales. No hay somitas cefálicos. Los somitas se corresponden casi totalmente con las vértebras.
La somita se va diferenciando en dos partes:
- Esclerotomo: de aquí derivan las vértebras y los discos intervertebrales.
- Dermomiotomo: se subdivide en o Dermatoma: se formará la dermis de la correspondiente metámera. o Miotomo Epimero: músculos del dorso de los canales vertebrales de la correspondiente metámera. Musculatura profunda. Hipomero: tres capas para dar los músculos ventrales del dorsos y la de los miembros correspondientes de la metámera. En la parte ventral hay otro tabique a partir del cual se forma la columna del músculo recto. En los niveles cervicales, torácicos y superiores lumbares vemos los esbozos de los músculos de los miembros.
Como el embrión continúa desarrollándose la somita se extiende y rodea al embrión. Ya tenemos genes de activación que condicionan el tipo celular en el que evolucionaran las células.
Hay un tabique intermuscular entre epímero e hipómero. Lo que queda más lejos del estímulo de no madurar, son los primero en madurar.
Extremidades:
En ciertos niveles a partir del hipómero de la zona, y gracias al mesodermo mesenquimal, se formarán esbozos de los miembros. Se formará de distal a proximal (5ª semana) viéndose la primera formación de la musculatura distal, pues se diferencian al perder el estímulo de no maduración predominante en estas zonas (núcleo mesenquimal).
Los miembros superiores salen de C3 a C8 y T1 y T2, hay un giro de 90º en dirección dorsal. Los miembros inferiores van de L2 a S2, sufren un giro 90º lateral, mientras que los inferiores giran 90º en dirección medial. La diferenciación va con 1 o 2 días de retraso respecto a los miembros superiores.
El repliegue ectodérmico apical viene del mesénquima de la hoja somática del mesodermo lateral.
A partir de la 5ª semana hay diferenciación de distal a proximal. Se dan unas constricciones circulares en el miembro (2, codo y muñeca) que forman 3 segmentos.
Las radiaciones digitales son un fenómeno de muerte celular de la manopla. Es inducido por el esqueleto de los huesos de distal a proximal.