¡Descarga embriologia y más Apuntes en PDF de Biología Celular solo en Docsity!
CAPÍTULO 13: GAMETOGÉNESIS, FECUNDACIÓN Y EMBRIOLOGÍA
I. FORMACIÓN DE LOS GAMETOS Y FECUNDACIÓN
La fecundación es el proceso por el cual los gametos (óvulo y espermatozoide) se fusionan para la formación de un nuevo individuo, cuyo genoma deriva del de los dos progenitores. Aunque con diferencias entre especies, se reconocen cuatro even- tos comunes:
- Contacto y reconocimiento. Asegura que la fecundación sólo tenga lugar entre individuos de la misma especie (o muy próximas).
- Regulación de la entrada del núcleo masculino en el óvulo: impide la polispermia.
- Fusión del material genético del óvulo y el espermatozoide.
- Activación del óvulo para iniciar el desarrollo.
I.1 Gametogénesis (fig. 1)
Los gametos se forman a partir de las células madre germinales, que sufren un proceso de reducción cromosómica ( meio- sis ), acompañado de una maduración celular, que en el caso de los espermatozoides es muy compleja y que tiene lugar después de la meiosis. En los óvulos, la maduración es temprana y consiste, principalmente en la síntesis de grandes cantidades de pro- teínas, tanto de “reserva” (vitelo), como las que permiten la fecundación (ejemplo receptores de membrana) y el posterior desa- rrollo (ejemplo, factores de transcripción característicos, que en muchos caso se distribuyen de forma asimétrica en el citosol). En la meiosis hay dos ciclos celulares consecutivos, sin una fase S intermedia, por lo que, partiendo de una célula con 2n cromosomas , se consiguen cuatro gametos con n cromátidas. Durante la profase de la primera de las divisiones se produce el apareamiento de los cromosomas, del que resultan los fenómenos de recombinación génica. Esta etapa es la más larga y en muchos casos se detiene durante períodos prolongados de tiempo: en el caso de las hembras de mamífero, se inicia antes del nacimiento y se prolonga hasta después de la pubertad, justo antes de cada ovulación. Por otra parte, la fecundación de los gametos femeninos puede producirse antes de que se complete la meiosis. En el caso de los mamíferos, la fecundación no es del óvulo, sino del ovocito secundario. Figura 1. Esquemas de la espermatogénesis (izda) y la ovogénesis (dcha) en mamíferos.
I.2 Espermatozoide
El producto final de la meiosis son las espermátidas, que sufren un proceso de maduración hacia los espermatozoides, con- sistente en la pérdida de casi todo el citoplasma, condensación del núcleo, desarrollo de un largo flagelo rodeado en la porción proximal por mitocondrias y la aparición de una gran vesícula de secreción y, por tanto, derivada del aparato de Golgi, el acro- soma , que contiene enzimas que degradan proteínas y azúcares complejos.
I.3 Óvulo
A diferencia del espermatozoide, que se libera de la mayoría del citoplasma, el óvulo tiene que mantenerlo y hacerlo crecer. El citoplasma del óvulo contiene, principalmente:
- Proteínas nutritivas (vitelo). La mayoría no las sintetiza el óvulo, sino el hígado, desde donde se distribuyen por vía san- guínea.
- Ribosomas, tRNAs y mRNAs.
- Factores morfogenéticos: factores de transcripción y moléculas que se segregarán, para ejercer efectos paracrinos en células próximas del futuro embrión.
- Cubiertas extracelulares. Por fuera de la membrana plasmática hay una cubierta formada por una matriz extracelular compleja, la envuelta vitelina (en mamíferos se denomina zona pelúcida ), que entre otras funciones tiene las de atraer y activar a los espermatozoides. Por fuera de la envuelta vitelina suele haber una cubierta gelatinosa.
- Córtex celular complejo, rico en actina y que contiene los gránulos corticales. Dichos gránulos son vesículas con enzi- mas proteolíticas, cuya exocitosis tiene lugar tras la penetración del espermatozoide.
I.4 Fecundación
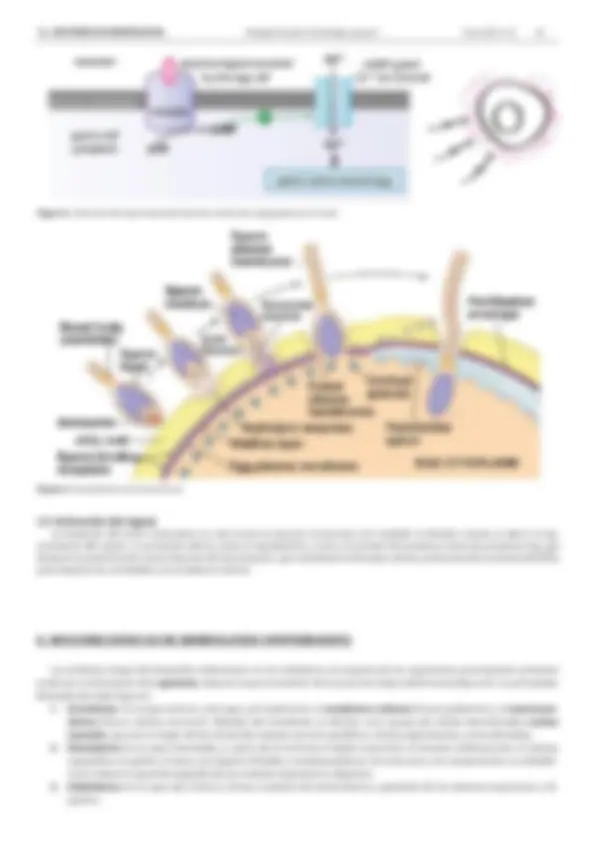
La interacción entre el espermatozoide y el óvulo suele incluir 5 etapas, que se resumen a continuación, tomando como referencia el modelo bien conocido de la fecundación en el erizo de mar y que, en muchos aspectos es equiparable a lo que ocurre en otros invertebrados y en cordados. La figura 2 esquematiza la primera etapa y la figura 3, las etapas segunda a quinta.
1. Atracción del espermatozoide por moléculas solubles que libera el óvulo. Dichas moléculas se unen a un receptor con actividad guanilil-ciclasa. El cGMP activa un canal de calcio, que entra a favor de gradiente en el espermatozoide, acti- vando su motilidad (fig. 2). El espermatozoide va a ir siguiendo el gradiente de las moléculas atractivas. 2. Reacción acrosomal. La membrana del espermatozoide reconoce ciertos azúcares de la cubierta del óvulo. Este es un reconocimiento especie-específico, es decir, característico de cada especie. Durante este reconocimiento, mediado por receptores (en el espermatozoide) y ligandos (en la cubierta del óvulo), en el espermatozoide se activan nuevos canales de Ca2+^ de la membrana y una fosfolipasa que libera IP 3 , por lo que sale también Ca2+^ del retículo endoplásmico. Esto ac- tiva la fusión de las membranas plasmática y del acrosoma, con la correspondiente exocitosis de las enzimas hidrolíticas, que degradan, primero la cubierta gelatinosa y más tarde la envuelta vitelina. Paralelamente, el Ca2+^ activa también la polimerización de la G-actina, que aparece en concentraciones muy altas por detrás del acrosoma. La F-actina forma así la prolongación acrosomal , que permite alcanzar a la membrana vitelina. 3. Unión del espermatozoide a la envuelta vitelina (zona pelúcida de mamíferos). Es también una unión especie-es- pecífica, mediada por pares moleculares: ligandos, de la membrana plasmática del espermatozoide y receptores, de la envuelta vitelina. De esta manera se produce el anclaje necesario del espermatozoide en la superficie del óvulo. 4. Fusión de las membranas de los gametos. Las hidrolasas del acrosoma terminan por romper también la envuelta vite- lina, facilitando el contacto entre las membranas plasmáticas de ambos gametos. En dichas membranas existe una pro- porción muy alta de las denominadas proteínas ”fusogénicas”, altamente hidrofóbicas, por lo que ayudan en la fusión. Completada la fusión, el núcleo y el centriolo del espermatozoide entran el citoplasma del óvulo. El huso acromático necesario para la primera división se formará a partir de centriolo masculino (degenera el del óvulo). La entrada de un segundo núcleo sería fatal, pues se obtendría un zigoto triploide, por lo que nada más entrar el núcleo masculino se inicia el “bloqueo de la polispermia”, con dos fases, una rápida y otra lenta. La rápida depende de la despolarización del óvulo, gracias a la entrada de Na+^ por canales que se abren tras la fusión de las membranas: el potencial pasa de -70 mV a +20 mV. La fusión solo se puede producir si el óvulo se encuentra en su potencial de reposo (-70 mV); una vez despolarizada (+20 mV), la fusión es imposible. 5. Reacción cortical. La despolarización aludida en el epígrafe 4, es extraordinariamente rápida, pero limitada en el tiem- po (dura más o menos un minuto). Para evitar la polispermia pasado ese plazo, se produce la reacción cortical, que asegura un bloqueo lento, pero prolongado. La fusión de membranas no solo activa canales de sodio del óvulo, sino también canales de calcio. La consecuencia es la exocitosis de los gránulos corticales. Estos contienen: - Proteasas, que hidrolizan los anclajes entre la envuelta vitelina y la membrana plasmática. - Glicosaminoglicanos, que con su alta significación osmótica facilitan la entrada de agua, separando las membranas vitelina y plasmática. - Sustancias que modifican a la envuelta vitelina endureciéndola. Como consecuencia se forma la denominada en- vuelta de fecundación, que impide que entre un segundo espermatozoide.
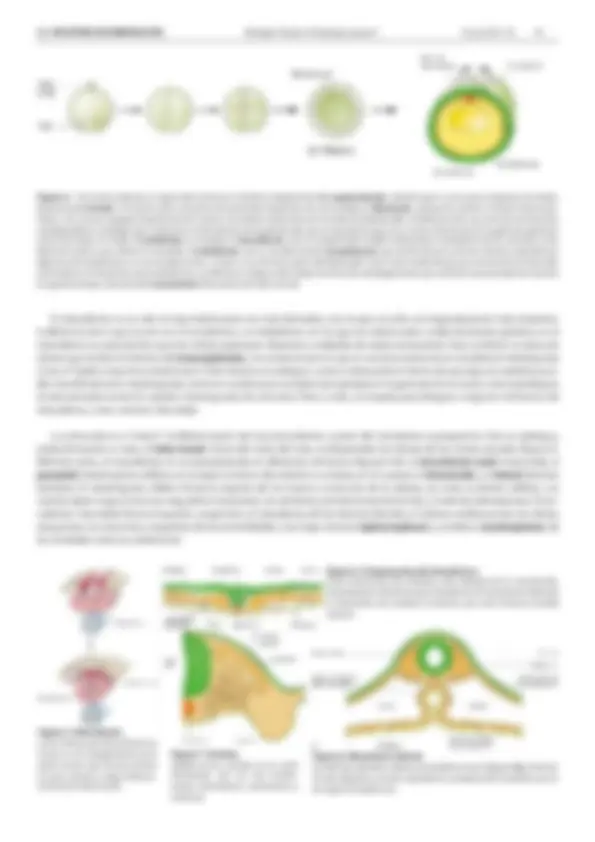
(e) Gastrula Figura 4. Tras la fecundación, el zigoto ( a ) comienza a dividirse rápidamente ( b , segmentación ), dando lugar a una masa compacta de células, denominada mórula. La mórula sufre un proceso de cavitación (aparición de una cavidad, el blastocele , porque las células re retiran hacia la pe- riferia, a la vez que segregan líquido hacia el centro). El embrión recibe ahora el nombre de blástula ( d ). La blástula sufre una serie de movimientos morfogenéticos complejos que conducen a la formación de la gástrula ( e ), que se representa aquí en un corte transversal. En la gástrula aparecen ya las tres hojas: en verde, el ectodermo , en naranja, el mesodermo , con un componente medial (notocorda y mesodermo de los somitas) y otro lateral (el resto) y, por último, en amarillo, el endodermo , con su cavidad central ( arquénteron ) que será la futura luz de los sistemas respiratorio y digestivo (el arquénteron es una cavidad nueva, es decir, no se forma a partir del blastocele). En la zona media dorsal, por encima de la notocorda, el ectodermo se denomina neuroectodermo. Las flechas en la figura ( e ) señalan la dirección del plegamiento que sufrirá el neuroectodermo durante la siguiente etapa, denominada neurulación (formación del tubo neural). El mesodermo no es sólo la hoja embrionaria con más derivados, sino la que se sufre una regionalización más temprana. A diferencia de lo que ocurre con el ectodermo y el endodermo, en los que las células están unidas formando epitelios, en el mesodermo es característico que las células aparezcan dispersas y rodeadas de matriz extracelular. Esta condición es típica de células que reciben el término de mesenquimales , circunstancia por la cual, en muchas ocasiones se considera al mesénquima como el “tejido conjuntivo embrionario”. Este término es ambiguo, como lo demuestra el hecho de que algunos epitelios pue- den transformarse en mesénquimas, tanto en condiciones normales (por ejemplo en la gastrulación en aves), como patológicas (la denominada transición epitelio-mesenquimal de cánceres). Pese a todo, se emplea para designar a algunos territorios del mesodermo, como veremos más abajo. La notocorda va a “inducir” la diferenciación del neuroectodermo a partir del ectodermo suprayacente. Este se repliega y acaba formando un tubo, el tubo neural. Antes del cierre del tubo, se desprenden las células de las crestas neurales (figura 5). Mientras tanto, el mesodermo se va especializando en diferentes territorios (figuras 6-8): el mesodermo axial (notocorda), el paraaxial (mesénquima cefálico, en la región anterior del embrión y somitas, en el cuerpo), el intermedio y el lateral (láminas laterales). El mesénquima cefálico forma la mayoría de los huesos y músculos de la cabeza, así como la dermis cefálica. Los somitas darán lugar al músculo esquelético (miotomo), las vértebras y tendones (esclerotomo), y la dermis (dermatomo). El me- sodermo intermedio forma el aparato urogenital y el mesodermo de las láminas laterales, el sistema cardiovascular, las células sanguíneas, los músculos y esqueleto de las extremidades y las hojas visceral ( esplacnopleura ) y somática ( somatopleura ) de las cavidades torácica y abdominal. Figura 5. Neurulación. Corte transversal del embrión en el que se ve el plegamiento de la placa neural, que forma primero el surco neural y luego evolucio- na hacia el tubo neural. Figura 6. Componentes del mesodermo. Corte transversal del embrión, justo después de la neurulación, mostrando los elementos del mesodermo. En las láminas laterales se desarrolla una cavidad, el celoma, que será la futura cavidad visceral. Figura 7. Somitas. Detalle de los somitas en un corte transversal, con sus tres compo- nentes: dermatomo, esclerotomo y miotomo. Figura 8. Mesodermo lateral. Las láminas laterales rodean al endodermo (ver la figura 4e ) y forman el tubo digestivo y tracto respiratorio, exceptuando el epitelio, que es de origen endodérmico.