









Prepara tus exámenes y mejora tus resultados gracias a la gran cantidad de recursos disponibles en Docsity

Gana puntos ayudando a otros estudiantes o consíguelos activando un Plan Premium

Prepara tus exámenes
Prepara tus exámenes y mejora tus resultados gracias a la gran cantidad de recursos disponibles en Docsity
Prepara tus exámenes con los documentos que comparten otros estudiantes como tú en Docsity
Encuentra los documentos específicos para los exámenes de tu universidad
Estudia con lecciones y exámenes resueltos basados en los programas académicos de las mejores universidades
Responde a preguntas de exámenes reales y pon a prueba tu preparación
Consigue puntos base para descargar
Gana puntos ayudando a otros estudiantes o consíguelos activando un Plan Premium
Comunidad
Pide ayuda a la comunidad y resuelve tus dudas de estudio
Ebooks gratuitos
Descarga nuestras guías gratuitas sobre técnicas de estudio, métodos para controlar la ansiedad y consejos para la tesis preparadas por los tutores de Docsity
Asignatura: bioq, Profesor: Felix Busto, Bioquimica, Carrera: Veterinaria, Universidad: UNILEON
Tipo: Apuntes
1 / 12

Esta página no es visible en la vista previa
¡No te pierdas las partes importantes!
Las enzimas son proteínas específicas con propiedades catalíticas que aceleran la velocidad de una reacción química, no sufriendo transformaciones durante el proceso, es decir, que permanecen químicamente invariables al final de la reacción.
En algunos casos, las enzimas precisan de componentes no proteicos para llevar a cabo su actividad, denominados cofactores, que pueden ser iones metálicos (Mg, Mn, Zn...) o moléculas orgánicas, llamadas coenzimas, las cuales pueden estar unidas a la enzima a través de interacciones débiles, como pueden ser puentes de hidrógeno, fuerzas de Van der Waals (como es el caso del NAD, coenzima A) o por enlaces covalentes, denominándose grupos prostéticos (caso de FAD, biotina)
S
P
(S … P) (Estado de transición)
Estado inicial Energía libre Estado final Progreso de la reacción
DG^ (no catalizada)
DG^ (catalizada)
Las enzimas aceleran las reacciones porque hacen que disminuya la energía de activación (DG) del proceso, favoreciendo la formación del estado de transición
DG (de la reacción)
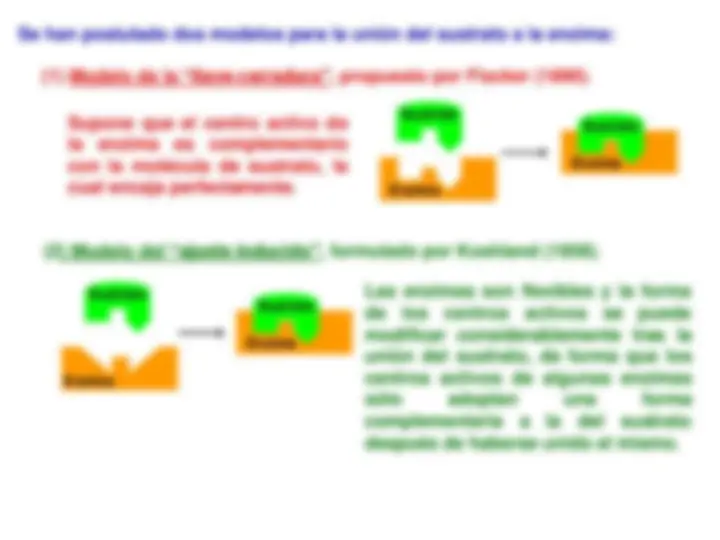
Se han postulado dos modelos para la unión del sustrato a la enzima:
(1) Modelo de la “llave - cerradura” , propuesto por Fiscker (1890).
Enzima
Sustrato
Enzima
Supone que el centro activo de Sustrato la enzima es complementario con la molécula de sustrato, la cual encaja perfectamente.
(2) Modelo del “ajuste inducido” , formulado por Koshland (1958).
Sustrato
Enzima
Enzima
Sustrato
Las enzimas son flexibles y la forma de los centros activos se puede modificar considerablemente tras la unión del sustrato, de forma que los centros activos de algunas enzimas sólo adoptan una forma complementaria a la del sustrato después de haberse unido el mismo.
Clasificación de las enzimas
La velocidad de la reacción (v) es: v =^ k 2 [ES]^ (1)
Velocidad de formación de ES (v 1 ): v^1 =^ k^1 [E][S] Velocidad de descomposición de ES (v-1): v-1 = k-1 [ES] + k 2 [ES] = ( k-1 + k 2 )[ES] Condición de estado estacionario:
k 1 [E][S] = ( k-1 + k 2 )[ES]
k-1 + k 2 k 1
Condición de conservación de la materia:
k-1 + k 2
k 1 [E][S] (2)
Si dividimos la expresión (1) por [E]T, tenemos: v [E]T
k 2 [ES] [E]T
v [E]T
k 2 [ES] [E]+[ES]
Sustituyendo [ES] por el valor de la expresión (2), tenemos:
v [E]T^ =
k 2 [E]+[E][S]
k-1 + k 2
k 1
k-1 + k 2
k 1
v [E]T^ =
k 2 1+[S]
k-1 + k 2
k 1
k-1 + k 2
k 1
Multiplicando numerador y denominador por^ k-1^ +^ k^2 , tenemos: k 1
v 1 = v-
v [E]T
k 2
k-1 + k 2 k 1
Si definimos: k-1 + k 2 k 1^ =^
Km =
Km = Constante de Michaelis
tenemos: v [E]T^ =^
k 2 Km + [S]
v (^) = k 2 [E]T
Km + [S]
k 2 [E]T = Vmax Vmax es la máxima velocidad que se puede obtener cuando toda la enzima está presente en forma del complejo ES.
v (^) = Vmax
Km + [S]
Ecuación de Michaelis-Menten
Si k 2 <<<k-1 (condición de equilibrio rápido): Km =
k- k 1
Representación directa (v vs [S]) de la ecuación de Michaelis-Menten
v (^) = Vmax
Km + [S] Proporciona una curva hiperbólica
Km
lim v = 0 Vmax [S] 0
lim v = Vmax
Vmax 2
Velocidad de reacción [Sustrato]
Representación doble inversa (1/v vs 1/[S]) (Lineweaver-Burk)
v (^) = Vmax
Km + [S] v^
Vmax [S]
1 Km + [S] ;
v =
Vmax
Km
[S]
Vmax
Al representar 1/v vs 1/[S], se obtiene una línea recta:
-1/Km 1/Vmax
Ordenada (b) = 1/Vmax
Pendiente (m) = Km/Vmax
Abscisa (x) = -1/Km
Ecuación de Lineweaver-Burk
1/v
1/[S]