






Prepara tus exámenes y mejora tus resultados gracias a la gran cantidad de recursos disponibles en Docsity

Gana puntos ayudando a otros estudiantes o consíguelos activando un Plan Premium

Prepara tus exámenes
Prepara tus exámenes y mejora tus resultados gracias a la gran cantidad de recursos disponibles en Docsity
Prepara tus exámenes con los documentos que comparten otros estudiantes como tú en Docsity
Encuentra los documentos específicos para los exámenes de tu universidad
Estudia con lecciones y exámenes resueltos basados en los programas académicos de las mejores universidades
Responde a preguntas de exámenes reales y pon a prueba tu preparación
Consigue puntos base para descargar
Gana puntos ayudando a otros estudiantes o consíguelos activando un Plan Premium
Comunidad
Pide ayuda a la comunidad y resuelve tus dudas de estudio
Ebooks gratuitos
Descarga nuestras guías gratuitas sobre técnicas de estudio, métodos para controlar la ansiedad y consejos para la tesis preparadas por los tutores de Docsity
Extincion sobre la reduccion del genoma
Tipo: Traducciones
1 / 7

Esta página no es visible en la vista previa
¡No te pierdas las partes importantes!
¿Con cuántos genes puede sobrevivir
una célula? El estudio de ciertas bacterias
simbiontes, que se hallan al borde
de la extinción por pérdida de genes,
arroja luz sobre esta cuestión
VICENTE PÉREZ BROCAL, AMPARO LATORRE Y ANDRÉS MOYA
■ Las bacterias que viven en simbiosis con insectos son un modelo idóneo para estudiar la reducción de los genomas en la naturaleza.
■ La reducción del genoma lleva a la conversión de las bacterias en orgánulos, a su extinción y sustitución por otra bacteria o a su perpetua ción a lado de otras bacterias. ■ Al concepto de genoma mínimo se ha llegado por dos grandes vías, la vía computa- cional y la vía experimental, que tienden a integrarse.
£!!•
'* _*_ *
V f
A
1
iMlfi iV
SfiJ
' * • ■ •
?
1. UN GENOMA PUEDE SOPORTAR la pérdida de ciertos genes. siempre que no sean esenciales. Cuando la reducción genónuca altera las funciones celulares vitales, la especie puede llegar a extinguirse.
■ i
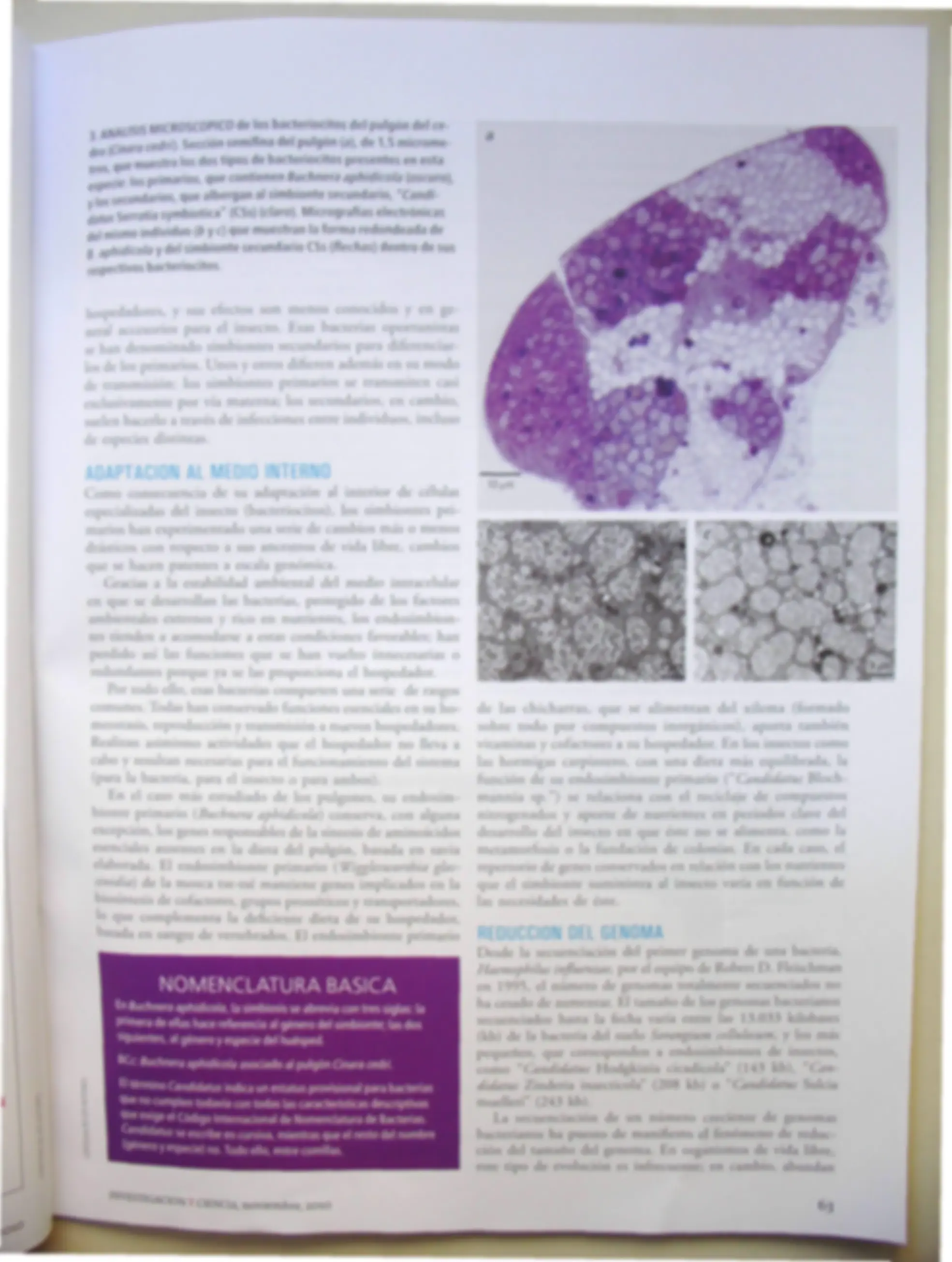
ANÁLISIS MICROSCÓPICO de los bacteriocitos del pulgón del ce- íCinara cedri). Sección semifina del pulgón {a), de 1,5 microme- que muestra los dos tipos de bacteriocitos presentes en esta oécie: los primarios, que contienen Budinera aphidicola {oscuro), los secundarios, que albergan al simbionte secundario, "Candi datos Serratia symbiotica" (CSs) {claro). Micrograflas electrónicas del mismo individuo {b y c) que muestran la forma redondeada de 0 aphidicola y del simbionte secundario CSs {flechas) dentro de sus respectivos bacteriocitos.
hospedadores, y MIS efectos son menos conocidos y en ge- IUMi,|^ ,,,,,.MIH.S^ para el insecto. Esas bacterias oportunistas se han denominado simbiontes secundario para dif¡ n m ¡ar los ,|, [os primarios. I fnos y otros difieren además en su modo de transmisión: los simbiontes primarios se transmiten casi oclusivamente por vía materna; los secundarios, en cambio, Mirlen hacerlo .1 través de inn 11 iont 1 ntre individuos, incluso i!, espet ies distintas.
ADAPTACIÓN AL MEDIO INTERNO Como consecuencia de su adaptación al interior de células specializadas del insecto (bacteriocitos), los simbiontes pri marios han experimentado una serie de cambios más o menos drásticos con respecto a sus ancestros de vida libre, cambios que se hacen patentes a escala genómica. Gradas a la estabilidad ambiental del medio ¡niracelular en que se desmolían las bacterias, protegido de los factores ambientales externos y rico en nutrientes, los endosimhion- tes tienden a acomodarse a esias condiciones favorables; han perdido así las (unciones que se han vuelto innecesarias o redundantes porque va se las proporciona el hospedador. Por todo ello, esas bacterias comparten una serie de rasgos comunes. Todas han conservado funciones esenciales en su ho- meostasis, reproducción y transmisión a nuevos hospedadores. Realizan asimismo actividades que el hospedador no lleva a cabo y resultan necesarias para el funcionamiento del sistema (para la bacteria, para el insecto o para ambos). En el caso más estudiado de los pulgones, su endosim- bionte primario {Budinera aphidicola) conserva, con alguna p< ion, los genes responsables de la síntesis de aminoácidos esenciales ausentes en la dieta del pulgón, basada en savia elaborada. El endosimbionte primario (Wigglcsworíhia gfos~ sinidia) de la mosca tse-tsé mantiene genes implicados en la biosíntcsis de cofactores, grupos prostéticos y transportadores, lo que complementa la deficiente dieta ele su hospedador, basada en sangre de vertebrados. El endosimbionte primario
t*
NOMENCLATURA BÁSICA En Buchnera aphidicola, la simbiosis se abrevia con tres siglas: la Primera de ellas hace referencia al género del simbionte; las dos siguientes, al género y especie del huésped. *
BCc: Buchnera aphidicola asociado al pulgón Cinara cedri. El término Candidatus indica un estatus provisional para bacterias que no cumplen todavía con todas las características descriptivas que exige el Código Internacional de Nomenclatura de Bacterias. andidatus se escribe en cursiva, mientras que el resto del nombre (género y especie) no. Todo ello, entre comillas.
10 pm *.
'NVES11GACION Y CIENCIA, noviembre. 2010
de las chicharras, que se alimentan del xilerna (formado sobre todo por c o m p u e s t o s inorgánicos)» aporra también vitaminas y cofactores a su hospedador. En los insectos como las hormigas carpintero, con una dieta más equilibrada, la función de su endosimbionte primario (uCandidatttí Bloch- mannia sp.") se relaciona con el reciclaje de compuestos nitrogenados y aporte de nutrientes en períodos clave del desarrollo del insecto en que éste no se alimenta, como la metamorfosis o la fundación de colonias. En cada caso, el repertorio de genes conservados en relación con los nutriente que el simbionte suministra al insecto varía en función de las necesidades de éste.
REDUCCIÓN DEL GENOMA Desde la secuetu iación del primer genoma de una bacteria, liaemophilus influenzae, por el equipo de Kobert D. FIcischman en 19°0, el número de genomas totalmente sccucnciados no ha cesado de aumentar. El tamaño de los genomas bacterianos secuenciados hasta la fecha varía entre las 13.033 kílobases (kb) de la bacteria del suelo Sorangium cellulosum* y los más I» qucfioSi que corresponden a endosimhiontes de insectos, 1 omo "Candidatus Hodgkinia cicadícola" (143 kb), "G/w- didatus Zinderia insecticola" (208 kb) o "Candidatus Sulcia muelleri" (243 kb). I a secuenciación de un número creciente de genomas bacteriano, ha puesto de manifiesto el fenómeno de reduc ción del tamaño del genoma. En organismo, de vida libre, le tipo de evolución es infrecuente; en cambio, abundan
63
los casos referidos ^ parásitos y mutualistas intracelularcs obligados, Entre estos últimos, cabe mencionar la secuenciación realizada por nuestro equipo dd genoma de Buchnem aphidicola aso< iado al pulgón Gnant cedrí (Bcc). Se trata del octavo endosimbionte primario de insectos secuen- ciado por completo. I os anteriores correspon den n & aphidicola^ endosimbionte de otras tres especies de pulgones {Acyrtosiphon puum> Schizaphh graminum y B<iizongi<i pistaciae)> ¿os especies del endosimbionte de las hor migas carpintero {^Candidatos Blochmannia floridanus" y %%Candidatus B, pennsylvani- cus"), el de la mosca tse-tsé {Wigglesworthia ^lossivirlid) y el de la chicharra ^Candidatos Baumannia cicadellicola"). Otros proyectos de secuenciación están en camino. ¿Cómo se ve afectado el genoma de esas bacterias por la pérdida selectiva de genes? ¿Por que unos genes se han retenido en mayor proporción que oíros? En primer lugar, el habitat de esas bacterias y su modo de transmisión han resultado determinantes en su evolu ción. Por tratarse de bacterias confinadas en un habitat muy restringido y aislado del exterior, presentan poblaciones de tamaño reducido y con escasas posibilidades de intercam bio de material genético con otras poblaciones o especies. Asimismo, medran en un ambiente rico en nutrientes y protegido. Todo ello ha provocado un debilitamiento de la selección natural en esas poblaciones; esto es, las mutacio nes perjudiciales que en otras circunstancias provocarían la eliminación de los individuos portadores de las mismas no son retiradas de la circulación. El genoma de las bacterias se va degradando de forma progresiva, con la consiguiente pérdida de función de genes.
Sorangium ceilulosum Bradyrtiizobium japonicum Vibrio parahaemoÍ)l¡cus EscbeñchiacoliMGieSS Yersinia pestis Bacillus subtilis Pnxhlonxoccus marinus CCMP
Saimonel ¡a entérica M^obacterium leprae Haemophilus influenzae Wolbachia Dm RtckettsJa prowazekii Chtamydia trachomatis Mycoplasma pneumonía? M. genitalium Blochmannia pennsyivankus B floridanus Wigglesworthia glossinidia Baumannia cicadeilinicoia Budinera aphidicola BSg & aphidicola BAp Blartabacterium sp. a aphidicola BBp & aph;d¡cola BCc 'Candidatos Carsoneüa ruddii" 'Canádatus Hoógkinin cícadicola" Nanoarthaeum equitans
Al hallarse
confinadas
en hábitats aislados
y ricos en nutrientes,
las bacterias
endosimbiontes
apenas sufren
la presión de la
selección natural
Si [a ausencia de esos genes no entraña grav consecuencias para la supervivencia del sistCnia éste se desprenderá de ellos en breve tiCm ' El resultado es la pérdida paulatina de (^) genes Puesto que los genomas bacterianos se carácter- zan por su elevado grado de comparación (Cas¡ codo el genoma lo constituyen secuencias codi. ficantes, con una escasa proporción de regio^ intergénicas), la pérdida de genes se traduce de inmediato en una reducción proporcional (^) eil el tamaño del genoma. Por tanto, los genon^ de menor longitud reflejan una mayor pérdida de genes a lo largo de la evolución.
EL FIN DE LA SIMBIOSIS B. aphidicola BCc representa un claro ejemplo de inmunización genómica en bacterias, con un genoma de 422 kb y tan sólo 362 genes codificantes de proteínas. Presenta, además, otra característica de sumo interés: todas las poblaciones de pulgones del cedro que liemos analizado contienen en abundancia una segunda bacteria endosimbionte, "Candidatus Serratia symbiotica", con siderada simbionte facultativo en otras especies de pulgones. Su morfología no guarda semejanza con la de otras cepas de su especie, sino más bien con la de B. aphidicola. Estudios preliminares sobre el genoma de S. symbiotica sugieren que éste sintetiza dos nutrientes esenciales que B. aphidicola BCc no aporta a su hospedador: triptófano y riboflavina. Según los datos microscópicos, funcionales y evolutivos, B. aphidicola BCc habría perdido parte de sus funciones mutua listas como consecuencia de su extrema degradación eenómica. La simbiosis observada implicaría, pues, a tres actores: el pulgón. B. aphidicola y S. symbiotica. Si este proceso de degradación genómica continúa —algo que nuestros datos apuntan—, d destino de esta cepa de B. aphidicola terminaría en la pérdida
Longuuddelcjenon,, (^) ( U | o b a (^) '2. I -1.
GENOMICO de diferentes bact rias.de acuerdo con su mod° de vida: bacterias de vida W* o patógenos no obligados^ * patógenos obligados ( m a ^ endomutualistas obligados (violeta), asi como los casos C. ruddii y H. cicadicob ( ^ Se muestra además el tania"^ del genoma de la arquea A/ * r tens [azul), que no es una W ria propiamente dicha.
m 1
es O ; s
^ ^ T l G A a o N Y CIENCIA, noviembre-^301
I
¿CUANTOS GENES NECESITA
UNA CÉLULA PARA VIVIR?
experimentales (rojo) o combinadas (verde).
1996 XI 2003
2006
1995 1999 2001 2002 2002 2003 2003 2004
2C
2006
QenitalnimyH.inlh « genom. 3 cepas de fl k W.glon .■* ñotidam
. as de B. aphidicola y M. gemtalium 3 cepas de fl. aphidicola, R. piov. trachomatís E. coli, 3 c • de fl. aphidicola y W. glossinidia
Bacillus subtilis M. genitalium y M. pneumomae
fl. subtilis, M. genitalium, E. coli y W. glossinidia, fl. aphidicola BAp, BSg, BBp, 8. Iloridanus fl. subtihs. M. genitalium, E. coliv W. glossinidia, fl. aphidicola BAp, BSg, BBp, fl. iloridanus
256 150 277 180 156 245 ± 4 8
300 265-
620 271 433-
206
208
genes tic una célula viva, debería identificarse el menor grupo posible de genes necesarios para mantener una forma de vida celular en funcionamiento bajo las condiciones más favorables imaginables, esto es, en presencia de un complemento de nu trientes esenciales y la ausencia de estrés ambiental. Los intentos recientes por perfilar una definición y recons trucción del genoma bacteriano mínimo lian seguido dos estrategias complementarias, los métodos computacionales y las aproximaciones experimentales, así como una integración de ambas. Los análisis mediante genómica comparada computacional se fundan en el alineamiento de secuencias de A D N para la identificación de genes ortólogos. A pesar de que el «enoma mínimo obtenido dependerá de cuántos y cuáles sean los taxones comparados, estos estudios son de gran utilidad para entender que funciones resultan esenciales para definir una célula viva. Su principal limitación estriba en que pueden subestimar el número de genes candidatos a ser incluidos en el genoma mínimo. Por una parte, la identificación de genes ortóloo 0 's entre genomas lejanamente emparentados no siempre residía sencilla. Por otra, genes no ortólogos pueden desempeñar funciones similares en diferentes organismos. Ello implica que incluso bajo las mismas condiciones ambientales y de vida pueden prever numerosas versiones de genomas mínimos " Las estrategias experimentales se basan en datos indirectos obtenidos a partir de mutagénesis al azar o disrupción siste mática de genes. Se lian seguido tres grandes tipos de análisis- mutagénesis masiva mediante transposones (elementos genét" eos móviles que pueden moverse de una localización gen (Íntica a otra), uso de ARN antisentido para inhibir la expresión ir¿t ' *
¡ m jj( ;K|os en pro. eSOS mc.abólicos.^ Ello se debe a la imposibj.
!
PARA SABER MAS ^ ^ ^ ^ ^ ^ ^ ^
M (^) ° v a y A. Latorre en Science, vol. 314, págs. 312-313; 2006-
^. M o ^ FOR L.FE. R. Gil, V. Pérez Brocal. A. Latorre N- A- Logan H M ^ e (^) " ^ M^anisms andSignificares, * £ * Cambridge 2006 ' L a p p m^ - S c o t l^ y P- C F. Oyston. Cambridge Un.vers.ty
I I
!^° n^ ° 5: P é g s ^ T - Í ^ I ' r ' ^ - ' of Medical Micros _ (^) E
, 2^