¡Descarga Fotofosforilación y más Apuntes en PDF de Fisiología de las Plantas solo en Docsity!
Fotofosforilación
La fotofosforilación fue descrita por el grupo de ARNON en 1954, en esa década también se describe la asimilación de carbono por Calvin.
Se observa que en cloroplastos iluminados en presencia de NADP, ADP y Pi(fosfato) se produce la reducción de NADP+ y la síntesis de ATP. Se establece la relación entre magnitudes del transporte de electrones y la fotofosforilación. Además, descubre que ambos procesos están acoplados, es decir, si no hay tpte de electrones no hay fotofosforilación y viceversa.
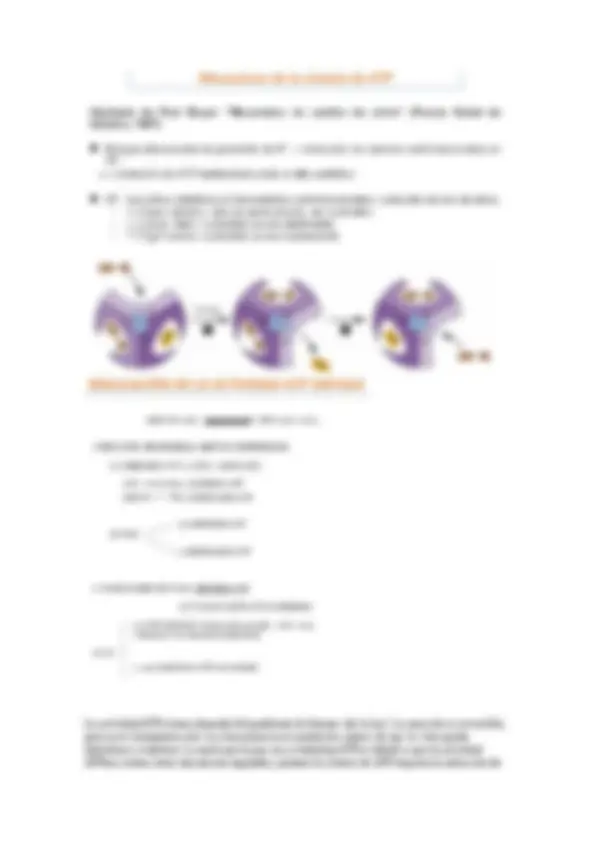
Todo transporte de electrones ya sea cíclico o no cíclico o pseudociclico genera un gradiente de energía que va a ser utilizado para sintetizar ATP.
Existe un proceso denominado control fotosintético hace referencia al gráfico. Cuando se aísla el cloroplasto, in vitro, se observa que en ausencia de adp y fosfato poco trasporte de electrones cuando se añade esto incrementa, esta diferencia de transporte de electrones se denomina control fotosintético.
Esto se debe a que si hay un pequeño transporte de electrones y no hay fotofosforilación se está disipando el gradiente de protones (hablaremos de esto más adelante)
En el tilacoide la energía se acumula en forma de gradiente de protones entre el estroma y el lumen tilacoidal. El aumento de protones en el lumen tilacoidal se produce con la reducción de plastoquinona a plastoquinol, la oxidación de plastoquinol y de agua, luego tenemos dos puntos de conservación de energia.
Se comprueba porque si se inhibe la reducción de plastoquinona disminuye la síntesis de atp, solo hay un punto de conservación de energía.
La relación ATP por cada 2 electrones o en vez de dos electrones oxigeno es la forma de valorar la relación entre ATP sintetizado y electrones transportados.
Si medimos reducción de atp esta razón de síntesis de atp cae aproximadamente a la mitad.
Fosfato asimilado por cada 2 electrones que se mueven en el sistema
La síntesis de ATP depende del flujo de electrones. La relación de la síntesis de ATP y el transporte de electrones es lo que describió ARNON.
En este esquema se encuentra todos los posibles: Tpte cíclico, no cíclico y pseudociclico.
En todos los casos se esta produciendo la síntesis de ATP porque se están transportandos electrones.
El sistema de reducción de plastoquinol, oxidacion de plastoquinol se encuentran los puntos. Y además en el tpte no cíclico y en el pseudocilico, es decir, cuando funciona el PSII se está oxidando agua en el lumen tilacoidal.
Cuando se mueve un proton, ion a traves de la mb se mueve una carga. Una carga que se desplaza a través de una mb genera una diferencia de potencial entre los dos puntos entre los que se está desplazando, es decir, modifica el potencial de la membrana. (NINGUN POTENCIAL SE PUEDE CALCULAR O DETERMINAR, SE MIDE LA DIFERENCIA DE POTENCIAL).
Ningun potencial se puede medir ya que para medirlo se necesita el potencial de membrana estándar de cada especie.
El potencial quimico de una subespecie J depende del potencial de membrana estándar de su concentración, de la preison ejercida sobre el sistema, de la carga y de la fuerza ejercida por la gravedad sobre esa especie J. Cuando esa especie J es una especie cargada, es un ion, el termino de presión es despreciable frente al termino eléctrico y si estamos viendo movs a nivel celular, es despreciable así que el termino de potencial químico para iones se queda convertido en potencial electroquímico que tiene un componente de carga y un componente de concentración.
Cuando se altera el potencial eléctrico, cuando se altera el potencial de la mb se activan mecanismos de rectificación. Se altera la actividad de proteínas de la membrana entre ellas las de tpte ya sean canales o incluso bombas, se van a activar canales que vuelvan al potencial de mb estándar. Se rectifica mediante el mov de iones en sentido contrario. En el caso del cloroplasto se rectifica por la salida de magnesio, sobre todo, y de potasio y la entrada de aniones iones cloruro, esto es rápido. De forma que la variación del potencial es rápido. Posteriormente se valora la concentración de protones que se calcula mediante el pH.
Cuando se da la luz el potencial eléctrico, la variación es instantánea, en el momento en el que se produce transporte de electrones se está alterando la concentración de cargas a ambos lados de la mb como consecuencia de la reducción de plastoquinona a plastoquinol, y de la oxidación de plastoquinol y de agua.
La variación de potencial es rápidamente amortiguada por el movimiento y la concentración de protones salen a través de la ATP sintasa. (La concentración incrementa a medida que pasa el
conocemos, el mecanismo de funcionamiento de Paul Boyer que ya está demostrada así que las siguientes diapos no están explicadas.
Lo importante es la regulación de la actividad
La actividad ATP sintasa depende del gradiente de fotones (de la luz). La reacción es reversible, pero en el cloroplasto solo va a funcionar en el sentido de sintesis de atp. In vitro puede hidrolizar y sintetizar. La razón por la que no se hidroliza ATP es debido a que la actividad ATPasa sintasa tiene mecanismo regulador, primero la síntesis de ATP requiere la reducción de
producción de ATP ya veremos con se hace (vamos que lo que pasa es que en general estos son los valores pero cuando veamos lo que hacen las superplantas los valores cambiaran).
Habiamos visto que el tpte de e va a generar un gradiente de energía debido a la generación de tpe de protones y a la oxidación de agua entre el lumen y el estroma tilacoidal,
Si anulamos el gradiente de protones que se origina consumiendolos protones o mendiante algun meanismo que permita salir a los protones del lumen tilacoidal se esta afectando al proceso de fotofosforilacion. Estos metabolitos que afectan al gradiente se denominan desacoplaodres porqueno afectan al transporte de electrones (ni la atpsintasa.)
Los protones van a acceder por la via que tenga mayor facilidad,por canales no por la atsintasa en la uqe los protones tienen que ir ordenados para salir. Ej: un transportador de potasio, otra posisbilidad es consumir protones mediante una sal amonica CLnH3 se va as va a disociar, lo que tenemos es un equilibrio amonio,amoniaco. SI tengo un ph elevado se va a desplzar hacia amoniaco, el amonio fluye porque es muy toxico ya que atraviesa libremente membrana.
Dentro de la mebrana hay un ph bajo por lo que se usan protones y el equilibrio se desplaza hacia el amonio.
Hay otros ekenentos que nno sosn desacopladoresss
Si se desacoplan F0 y F1 mediante la extraccion de magneiso con un quelantede cationes se sidoscian
Si inactivamos la enzima no afectamos al greadiente protonico, simplimente la enzima esta ing ¡hibidia se denomminan inhibidores de la energia de la enzima, se puede hacer bloqueando el sitiio activo de la enzima algunos de ellos estan en la diapo.
Siinhibiosla enzima ya seapor bloqueo del subcomplejo F0, el gradiente prtonico no se disipa se mantien o bien bloqueando, el transporte de electrones la por completo. Porque la fuerxa ionica