



!
!
GENÉTICA!
HUMANA!
Grado&en&Medicina&,&UJI&
&
&
&
&
&
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
























































































Prepara tus exámenes y mejora tus resultados gracias a la gran cantidad de recursos disponibles en Docsity

Gana puntos ayudando a otros estudiantes o consíguelos activando un Plan Premium

Prepara tus exámenes
Prepara tus exámenes y mejora tus resultados gracias a la gran cantidad de recursos disponibles en Docsity
Prepara tus exámenes con los documentos que comparten otros estudiantes como tú en Docsity
Encuentra los documentos específicos para los exámenes de tu universidad
Estudia con lecciones y exámenes resueltos basados en los programas académicos de las mejores universidades
Responde a preguntas de exámenes reales y pon a prueba tu preparación
Consigue puntos base para descargar
Gana puntos ayudando a otros estudiantes o consíguelos activando un Plan Premium
Comunidad
Pide ayuda a la comunidad y resuelve tus dudas de estudio
Ebooks gratuitos
Descarga nuestras guías gratuitas sobre técnicas de estudio, métodos para controlar la ansiedad y consejos para la tesis preparadas por los tutores de Docsity
Asignatura: Genètica Humana, Profesor: , Carrera: Bioquímica i Ciències Biomèdiques, Universidad: UV
Tipo: Apuntes
1 / 128

Esta página no es visible en la vista previa
¡No te pierdas las partes importantes!
-‐ El ordenamiento lineal de los genes: El biólogo Thomas H. Morgan (1866-‐1945) estudió el ciclo vital y la herencia en la mosca de la fruta Drosophila melanogaster, descubrió que: -‐ La recombinación génica ocurre durante la profase de la meiosis -‐ Los genes están localizados en los cromosomas (confirmando la teoría cromosómica de la herencia e iniciando la Citogenética ) -‐ Los genes están colocados de forma lineal en cada cromosoma (hoy sabemos: esa línea es la cadena de ADN) -‐ Realizó también los primeros mapas de ligamiento : es la asociación física entre 2 loci, puede definirse como la tendencia de los loci cercanos a heredarse conjuntos, debido a que las quiasmas se producen al azar. A mayor cercanía entre dos loci , menor probabilidad de quiasma y de generar variantes recombinantes y, por tanto, una mayor probabilidad de herencia conjunta. -‐ Existen tres tipos de mapas del genoma: -‐ Mapa genético: también llamado de ligamiento, se definen mediante unidades de recombinación genética llamadas Morgan (1 cM = 1% de probabilidad de recombinación entre dos genes). -‐ Mapa físico: definen la separación real de los loci mediante unidades de distancia: pares de bases (bp) -‐ Mapa cromosómico: también llamado cariotipo o ideograma de cromosomas, son la representación gráfica del patrón de bandas de los diferentes cromosomas cuando son teñidos (según compactación)
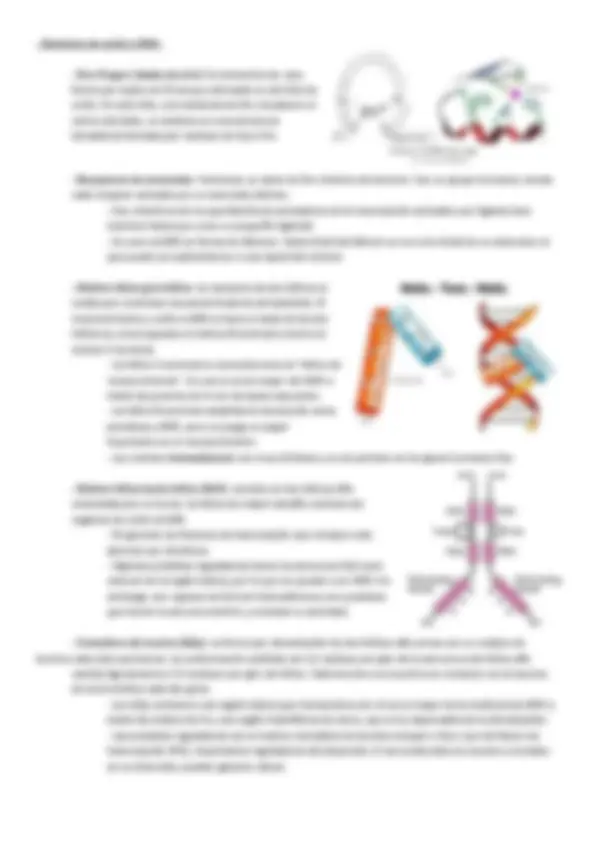
-‐ Leyes de Chargaff: -‐ A mitad de siglo prevalecía la idea de que el ADN era una molécula demasiado simple como para ser portadora de la información genética. -‐ En 1950, el científico Erwin Chargaff, llegó a la conclusión de que las 4 bases nitrogenadas no se encontraban en proporciones iguales en distintas especies , que sugería que el ADN no era tan monótono como se pensaba. -‐ Chargaff demostró además que, independientemente del origen del ADN, la proporción de purinas (A, G) era igual a la de pirimidinas (T, C). Es decir, había dos juegos de equivalencias, A y T por un lado y G y C por otro. Esto es lo que actualmente se llama complementariedad de bases. -‐ Ley de Chargaff: relación cuantitativa de los nucleótidos del ADN. Establece que la cantidad de Adenina es igual a la de Timina, y la cantidad de Citosina es igual a la de Guanina (n° purinas = n° pirimidinas) (A+G = C+T). Sin embargo existen diferencias en relación AT/CG, según la especie. -‐ Estudios de difracción de rayos X de Rosalind Franklin: En 1952-‐53, la biofísica Rosalind Franklin (1920-‐1958), realizó estudios cruciales de la estructura molecular del ADN mediante difracción de Rayos X de cristales puros de esta sustancia en estado nativo: -‐ Descubrió las formas A y B del ADN, demostró que las bases nitrogenadas estaban dirigidas hacia el interior de la molécula y dio detalles de las distancias moleculares entre repeticiones estructurales. -‐ Todos estos datos se infirieron de la famosa foto 51 , donde se observa una estructura de X (doble hélice). -‐ Sin embargo, no elaboró un esquema funcional de la molécula. -‐ Sus datos e informes confidenciales fueron usados por Watson y Crick -‐ Murió a los 37 años de carcinomatosis y cáncer de ovario, debido a las repetidas exposiciones a la radiación. -‐ La doble hélice de Watson y Crick: Basándose en los datos de Franklin y las proporciones de Chargaff, James Watson (1928) y Francis Crick (1916-‐2004) elaboraron un modelo molecular del ADN: -‐ El modelo consiste en dos hélices que corren en sentidos opuestos, con las bases de cada hélice dirigidas hacia el centro y guardando una relación espacial de estricta complementariedad. -‐ Esta estructura sugiere de inmediato una forma simple de autorreplicación de la molécula, por separación de las hélices y síntesis de las hélices complementarias. -‐ Consecuencias del modelo de la doble hélice: El modelo explicaba todas las propiedades esperables de una molécula que contuviese la información genética, en este caso el ADN: -‐ Autorreplicable por separación de las dos hélices -‐ Lineal a lo largo de su eje -‐ Localizada en los cromosomas como ya había sido demostrado por citólogos y bioquímicos. Los cromosomas están formados por proteínas y ADN, de ahí la duda entre las dos biomoléculas. -‐ Capaz de contener la información hereditaria , sugerido por la secuencia de bases con enorme cantidad de variaciones a lo largo de cada una de las hélices, acabando con la idea de que era monótono. -‐ Detalles estructurales: -‐ Total mente desenrollada, la molécula de ADN única de cada cromosoma humano mediría 7 cm -‐ Se trata de moléculas de longitud gigantesca pese a su anchura sub-‐microscópica (23nm) -‐ La molécula de ADN es flexible y puede curvarse, lo cual le permite adoptar sucesivos órdenes de curvatura, que a su vez permiten empaquetar estas moléculas larguísimas en los pequeños cromosomas
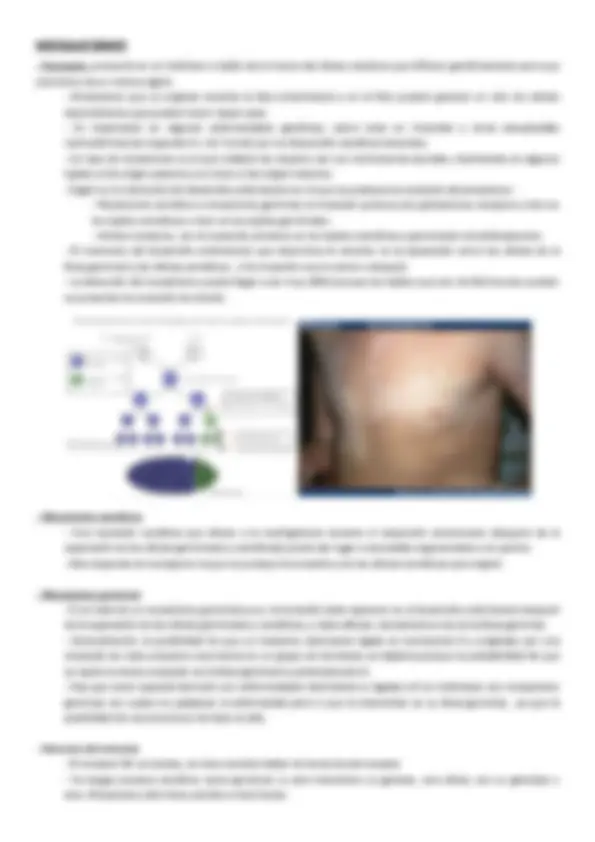
GENERALIDADES DEL GENOMA HUMANO -‐ El proyecto Genoma Humano: -‐ A partir del desciframiento del código genético, la genética sufrió un viraje, des de los estudios estructurales hacia los de descodificación. -‐ El clonaje e identificación de un único gen costaba unos 5 años de media en los años 80. En 2003 se publicó la secuencia (casi) completa de todo el genoma humano. Los saltos en genética continúan siendo vertiginosos. -‐ En presencia del presidente Clinton, los resultados de la secuenciación se hicieron públicos de forma conjunta por Francis Collins (director del programa gubernamental) y Craig Venter (de la empresa Celera) que había competido con la iniciativa pública. -‐ Consecuencias de la secuenciación del genoma humano: -‐ Se dispone de la secuencia de los 3.000 millones de bases que constituyen el ADN humano -‐ Estas 3.000 Mb están distribuidas en los 23 pares de cromosomas humanos -‐ La gran cantidad de información acumulada ha impulsado una nueva disciplina, la bioinformática , clave hoy en día en los estudios genéticos -‐ Se han detectado alrededor de 25.000 genes (mucho menor de lo esperado, aunque la complejidad del ADN no reside en el número de genes), todos los conocidos hasta el momento han sido secuenciados pero hay muchos cuya función es todavía desconocida. -‐ Se ha estimado que hay sobre un millón de proteínas ya que cada gen codifica para varias proteínas. Esto se consigue mediante: -‐ Splicing alternativo: de exones e intrones, lo que permite obtener distintas proteínas a partir del mismo transcrito primario y por tanto a partir del mismo gen. -‐ Diferentes promotores para un gen que lo activan en función del tejido y del momento del ciclo celular. Están regulados por factores de transcripción. -‐ Modificaciones postranscripcionales -‐ Modificaciones postraduccionales -‐ El genoma humano y sus características: -‐ 23 pares de cromosomas en el núcleo. 22 autosomas y uno sexual -‐ Una parte pequeña pero importante del genoma humano se localiza en las mitocondrias -‐ Características del ADN mitocondrial: -‐ Herencia materna exclusivamente -‐ Molécula circular de 16 kb (un 0,03% del tamaño del cromosoma más pequeño) -‐ Codifica sólo 37 genes que realizan su función en la mitocondria -‐ La gran mayoría de las proteínas mitocondriales son productos de genes nucleares -‐ Estructura del ADN: -‐ Estructura de soporte : Formada por unidades alternas de grupos fosfato y un azúcar, la desoxirribosa (pentosa). Las desoxirribosas se unen entre sí a través de enlaces fosfodiéster entre los átomos 3ʹ′ y 5ʹ′. La formación de enlaces asimétricos implica que cada hebra de ADN de una doble hélice tiene una dirección (se llaman antiparalelas) -‐ Bases nitrogenadas: Son adenina (A), citosina (C), guanina (G) y timina (T). Cada una de ellas unida al armazón de azúcar-‐fosfato a través del azúcar para formar el nucleótido completo (base-‐azúcar-‐fosfato). Las bases son compuestos heterocíclicos y aromáticos con dos o más átomos de nitrógeno y se clasifican en: -‐ bases púricas o purinas (A y G), derivadas de la purina y formadas por dos anillos unidos entre sí -‐ bases pirimidínicas o pirimidinas (C y T), derivadas de la pirimidina y con un solo anillo
ESTRUCTURA DE LOS CROMOSOMAS HUMANOS -‐ Doble hélice de ADN -‐ Nucleosomas: -‐ En los seres humanos hay cinco tipos principales: la histona H1 y las histonas H2A, H2B, H3 yH4. Estas últimas, llamadas histonas nucleosomales, forman un octámero con dos histonas de cada; alrededor de este núcleo se enrolla dos veces un hilo de ADN (147 bp aprox.) -‐ Este complejo ADN-‐histona recibe el nombre de nucleosoma y constituye el componente primario del cromosoma -‐ Tras el nucleosoma, el DNA se desplaza unos 20-‐70 bp en un giro hacia la izquierda hasta alcanzar el siguiente nucleosoma -‐ La pieza intermedia, también denominada DNA de conexión está “desnuda”. Las histonas H1 se colocan como pie a de cierre en cada nucleosoma y al mismo tiempo toma contacto con las agrupaciones vecinas -‐ De este modo, las proteínas H1 van “grapando” los nucleosomas para formar un hilo denso -‐ Cromatina. Fibras de 30 nm: -‐ La doble hélice de ADN de cada cromosoma se dispone formando la cromatina, maraña de fibras de 30nm -‐ En la cromatina las moléculas de ADN se acomplejan con las histonas y otras proteínas no tan bien conocidas -‐ Solenoides: -‐ Incluso en la interfase, el ADN se encuentra empaquetado en una estructura llamada solenoide, formados tras el arrollamiento de las fibras de 30nm -‐ Los solenoides se agrupan en bucles o dominios (de unos 100 kb) agregados por proteínas no histonas que forman la matriz nuclear -‐ Se ha propuesto que los bucles pueden ser la unidad funcional de replicación y/o de transcripción -‐ Así un punto de control de la expresión génica podría depender de la forma en que estén empaquetados el DNA y los genes en los cromosomas -‐ Cromosomas
-‐ ADN repetido disperso: consiste en secuencias de ADN que se repiten de modo disperso por todo el genoma, constituyendo el 35-‐ 40% del genoma humano. -‐ LINEs y SINEs : elementos más importantes que se distinguen por el tamaño de la unidad repetida -‐ Estas secuencias pueden autopropagarse al transcribirse a un ARNm intermediario, retrotranscribirse e insertarse en otro punto del genoma. Este fenómeno se produce con una baja frecuencia, pero podría ser el responsable de hasta 1 de cada 500 nuevas mutaciones. -‐ La inserción puede resultar lesiva por mutagénesis insercional o por desregulación de la expresión de genes próximos (estos elementos tiene su propio promotor) -‐ Esquema simplificado del mecanismo de retrotransposición de un elemento LINE y un SINE. Un elemento LINE es transcrito que sale del núcleo celular. En el citoplasma se traduce en sus dos marcos de lectura abiertos generando dos proteínas (se han representado como ORF1p y ORF2p). Ambas permiten retrotranscribir el ARNm del LINE y de otros retrotransposones no autónomos, como SINEs. Durante la retrotranscripción la nueva secuencia de ADN se integra en otro punto del genoma, pudiendo causar mutaciones. La retrotransposasa (transposasa) reconoce sitios diana en el ADN, inserta el retrotransposón (transposón) y luego intervienen la ligasa y la ADN polimerasa. -‐ Transposones: descubiertos por primera vez en el genoma de maíz por Barbara McClintock's, Premio Nobel 1983, son elementos que pueden moverse de manera autosuficiente a diferentes partes del genoma que puede causar mutaciones por inserción. Existen varias clases en función de su estrategia de transposición: -‐ Retrotransposones (transposones clase I): -‐ Un ejemplo son los LINEs y los SINEs, que utilizan una maquinaria retrotranscriptiva. -‐ Al utilizar retrotranscripción → aumenta la cantidad de ADN -‐ No está relacionado con la repetitividad de secuencias. -‐ La frecuencia de retrotranscripción en humanos es bajísima -‐ Transposones de ADN (transposones clase II): -‐ Los transposones de ADN completos pueden autopropagarse sin mediar la retrotranscripción. Un transposón contiene el gen de una enzima transposasa, flanqueado por repeticiones invertidas. -‐ Al no utilizar retrotranscripción → no aumenta la cantidad de ADN -‐ La transposasa codificada por el propio transposón lo extrae realizando dos cortes flanqueantes en la hebra de ADN, generando extremos cohesivos, y lo inserta en la secuencia diana en otro punto del genoma. Una ADN polimerasa rellena los huecos generados por los extremos cohesivos y una ADN ligasa restablece los enlaces fosfodiéster, recuperando la continuidad de la secuencia de ADN -‐ Hay múltiples familias, de las que cabe destacar los elementos mariner (así como las familias MER1 y MER2) por su importancia en medicina ya que pueden generar reordenaciones cromosómicas -‐ Transposones tipo III: -‐ Existe una última clase de transposones (clase III o MITEs). Son transposones cortos que se caracterizan por su alto número de copias y homogeneidad en tamaño. Son secuencias de unas 400 pares de bases flanqueadas por repeticiones invertidas de 15 pares de bases -‐ Son demasiado cortas para codificar una proteína
CICLO CELULAR: REGULACIÓN E INTERFASE -‐ Fases: -‐ Todas las células del cuerpo humano derivan de un óvulo fecundado gracias a miles de mitosis -‐ El período entre dos mitosis sucesivas se denomina interfase ( fases G1, S y G2 ) y es el estado en el que la célula pasa la mayor parte de su ciclo vital -‐ Algunos tipos celulares como las neuronas o los eritrocitos no se dividen en absoluto una vez plenamente diferenciados (entran en fase G0 ) -‐ Tras la mitosis la célula entra en la fase G1 , en la que posee una copia del genoma. -‐ La fase G1 se continúa con la fase S (6-‐8h), en la que tiene lugar la síntesis de ADN: cada cromosoma se duplica y pasa a estar formado por dos cromátidas hermanas unidas físicamente en el centrómero , una región de ADN satélite α que se asocia con una serie de proteínas específicas para formar el cinetocoro (estructura que sirve para acoplar cada cromosoma los microtúbulos del huso acromático). -‐ Tras la fase S, la célula entra en una breve etapa (2-‐4h) denominada G2 que termina cuando entra en mitosis (cuando los cromosomas se condensan y se hacen visibles al microscopio). -‐ Regulación: -‐ El ciclo celular está regulado por distintas moléculas reguladoras: -‐ Factores de crecimiento: estimulan el abandono de la fase G0 y el crecimiento celular de la fase G1. -‐ Factores de transcripción: como el E2F, indispensables para la transición de G1 a S. -‐ Factores promotores: son señales internas involucradas especialmente en la división celular. Destaca el factor promotor de la mitosis (MPF) y el complejo promotor de la anafase (APC) -‐ Ciclinas: proteínas que junto con las quinasas dependientes de ciclinas (CDK) forman complejos reguladores de las fases del ciclo celular. Su concentración varía de forma cíclica a lo largo del ciclo. -‐ Actúan en diversos puntos de control: -‐ Control de la fase G1 : si la célula ha adquirido un buen crecimiento, está en un entorno favorable y no existe daño en el ADN comienza la fase S de duplicación del ADN. -‐ Control de la fase S: si se ha duplicado el ADN con éxito y sin ningún daño la célula entra en la fase G2, deteniendo su crecimiento y preparándose para la división celular. -‐ Control de la fase G2: si la maquinaria de replicación del ADN no ha fallado y no está dañado y el crecimiento celular y el ambiente es óptimo, comienza la mitosis. -‐ Control de la metafase : si todos los cromosomas se encuentran alineados en el huso y sin ningún daño comienza la anafase y se predispone a la finalización de la mitosis. -‐ Funcionamiento de las ciclinas: -‐ Las ciclinas, que por sí solas no tienen actividad enzimática, interaccionan con las CDK. -‐ Las CDK se autofosforilan inducidas por la unión a las ciclinas. -‐ Ese complejo fosforilado tiene la capacidad de fosforilar sustratos y por tanto de activar e inactivar enzimas y factores reguladores. -‐ En eucariotas superiores existen diferentes quinasas de la familia de Cdc2 (Cdk’s) que intervienen en distintas fases del ciclo celular.
CICLO CELULAR: MEIOSIS -‐ Concepto: La meiosis es un tipo de división celular de las células germinales que da lugar a gametos haploides. Consiste en una ronda de síntesis de ADN seguida de dos rondas de segregación cromosómica y división celular -‐ Las dos divisiones consecutivas se denominan meiosis I y meiosis II -‐ Los cromosomas X e Y no son homólogos en sentido estricto pero tienen segmentos homólogos en los extremos de los brazos por donde se aparean -‐ Fases: -‐ La meiosis I es crucial porque en ella se produce la recombinación genética (entrecruzamiento meiótico) -‐ En este proceso se intercambian segmentos homólogos del ADN entre cromátidas no hermanas de las parejas de cromosomas, lo que asegura que ningún gameto es idéntico a otro -‐ La recombinación es fundamental para el mapeo de genes responsables de trastornos heredados -‐ Implica un proceso de entrelazamiento físico (quiasma, tétrada) y es también crucial para asegurar una segregación cromosómica correcta durante la meiosis. -‐ Si no se produce correctamente pueden aparecer errores en la segregación de los cromosomas, que es una causa frecuente de anomalías cromosómicas, como el síndrome de Down. -‐ También se llama división reduccional , pues se caracteriza por la reducción del número de cromosomas de diploide a haploide mediante el apareamiento de los homólogos en la profase y su segregación a diferentes células en la anafase de este ciclo -‐ La meiosis II se produce tras la meiosis I tras una breve pausa (intercinesis) en la que no hay síntesis de ADN. Como en una mitosis, las cromátidas de cada cromosoma se separan y pasan a cada célula hija. -‐ Meiosis I -‐ Profase I : La profase I de la meiosis I es un complicado proceso que difiere de la profase mitótica en varios aspectos cruciales. Se definen varias etapas en las cuales los cromosomas se van condensando: -‐ Leptoteno : Los cromosomas, que se han replicado durante la fase S precedente, se hacen visibles como finos filamentos que empiezan a condensarse. Las dos cromátidas hermanas están tan estrechamente alineadas que no pueden distinguirse. -‐ Cigoteno : Los cromosomas homólogos se emparejan a lo largo de toda su longitud. El emparejamiento o sinapsis es preciso y completo. Los cromosomas permanecen juntos gracias a un complejo sinaptonémico (estructura en forma de cinta que contiene proteínas) -‐ Paquiteno : Durante esta etapa, los cromosomas se enrollan de manera mucho más estrecha. La sinapsis es completa y cada par de homólogos aparece como un bivalente (también llamado tétrada). Se produce el entrecruzamiento meiótico -‐ Diploteno : Desaparece el complejo sinaptonémico y se separan los bivalentes. Al final de esta fase, los bivalentes sólo aparecen unidos por los quiasmas (puntos de entrecruzamiento). En espermatocitos humanos se aprecian 50 quiasmas de media (es decir varios por bivalente) -‐ Diacinesis : En esta etapa los cromosomas alcanzan su máxima condensación -‐ Metafase I : Como en la mitosis, la metafase I empieza cuando desaparece la membrana nuclear. Se ha formado un huso y los cromosomas apareados se alinean en el plano ecuatorial, con sus centrómeros orientados hacia polos diferentes.
-‐ Anafase I : Los dos miembros de cada bivalente se separan y sus respectivos centrómeros son dirigidos a los polos opuestos de la célula, un proceso denominado disyunción: -‐ Los distintos bivalentes se reparten aleatoriamente entre las células hijas, de manera que los conjuntos maternos y paternos originales se distribuyen al azar. Combinaciones posibles 223 (> 8 millones, prácticamente 100% de gametos distintos). La variación aumenta por el entrecruzamiento. -‐ Casi todos los errores en la división celular llevan a la muerte celular pero algunos causan alteraciones en la agregación de cromosomas durante la anafase (dos homólogos se desplazan al mismo polo). Este proceso conocido como no disyunción tiene consecuencias genéticas graves -‐ Telofase I : Los dos conjuntos haploides de cromosomas se encuentran agrupados en los polos opuestos -‐ Citocinesis: Después de la telofase I la célula se divide en dos células hijas haploides y entra en una breve interfase meiótica llamada intercinesis. -‐ En la espermatogénesis, el citoplasma se divide en partes más o menos iguales entre las dos células hijas -‐ En la ovogénesis, una célula hija (el ovocito secundario) recibe casi todo el citoplasma mientras que el otro se convierte en el primer corpúsculo polar -‐ La interfase meiótica no tiene fase S, es decir, no hay síntesis de ADN entre las dos divisiones meióticas -‐ Meiosis II: Es similar a una mitosis normal excepto en que el número de cromosomas de la célula que entra en meiosis II es haploide. Se obtienen cuatro células hijas haploides. -‐ Algunas consecuencias de la no disyunción: (es mucho más frecuente en cromosomas sexuales) -‐ Síndrome de Turner: X. Mujeres estériles y con escaso desarrollo de los caracteres sexuales. -‐ Síndrome de Klinefelter: XXY. Hombres estériles, testículos poco desarrollados y características sexuales secundarias con tendencia a la feminidad. -‐ Síndrome del triplo X: XXX. Frecuentemente son mujeres normales y en unos pocos casos presentan menor desarrollo físico y mental. -‐ Un hombre con 44 autosomas + Y no es viable debido a que la información genética contenida en el cromosoma X en muchísimo mayor que la contenida en Y y lo hace imprescindible para la vida
PROBLEMAS -‐ Se sabe que existe una serie de 4 alelos de un determinado gen en humanos; ¿Cuántos estarían presentes en: a) ¿un cromosoma haploide? b) ¿un par cromosómico (diploide)? c) ¿un individuo de la especie? d) con la misma base:¿cuántas combinaciones diferentes de alelos se puede esperar en población completa? a) 1 b) 1 ó 2 c) 1 ó 2 d) 10. AA, AB, AC, AD, BB, BC, BD, CC, CD, DD -‐ Si dos pares de genes A:a y B:b se transmiten independientemente y se sabe que A es dominante sobre a y B dominante sobre b, ¿cuál es la probabilidad de obtener: a) un gameto AB a partir de un individuo AaBb? b) un gameto Ab a partir de un individuo AABb? c) un descendiente AABB a partir de un cruzamiento aabb X AABB? d) un fenotipo AB a partir de un cruzamiento AaBb X AaBb? e) un fenotipo AB a partir de AABB X aabb? f) un fenotipo aB a partir de AaBb X AaBB? a) 25% b) 50% c) 0% d) 9 / e) 100% f) 2 / 8 -‐ En los siguientes pedigríes humanos los símbolos negros representan una condición fenotípica anormal, heredada de manera mendeliana simple. En cada pedigrí: a) Indique si la condición anormal es dominante o recesiva b) Describa los genotipos de todos los individuos que pueda
-‐ La fenilcetonuria (PKU) es una enfermedad hereditaria humana que impide al organismo metabolizar la fenilalanina contenida en las proteínas que ingerimos. La PKU se debe a un alelo recesivo mendeliano. Una pareja desea tener hijos, pero consulta a un consejero genético porque el hombre tiene una hermana con PKU y la mujer tiene un hermano con PKU. No se conocen otros casos en sus familias. Piden al consejero genético que determine la probabilidad de que su primer hijo padezca de PKU. ¿Cuál es esta probabilidad? Como máximo del 25% -‐ La enfermedad de Tay-‐Sachs es una enfermedad humana muy poco frecuente en la que sustancias tóxicas se acumulan en las células nerviosas. El alelo recesivo responsable de la enfermedad es heredado en una forma Mendeliana simple. Una mujer sana tenía una hermana que murió de la enfermedad está preocupada por la posibilidad de que su hijo, aún por nacer, esté afectado. ¿Cómo la aconsejaría? En primer lugar deber averiguar los antecedentes del padre, si es posible -‐ Si un hombre de grupo sanguíneo AB se casa con una mujer de grupo sanguíneo A, cuyo padre era O. ¿Qué grupos sanguíneos pueden esperar entre sus hijos? Si el padre de la mujer A era O (homocigoto) ella es AO. ABxAO: -‐ Genotipo: 25% AA, 25% AO, 25% AB, 25% BO -‐ Se podrá esperar que el 50% de los niños sea A, el 25% AB y el 25% B. -‐ En un centro de maternidad han confundido a cuatro recién nacidos. Se sabe que los tipos ABO de los cuatro niños son O, A, B y AB. Se analizan entonces los ti pos ABO de las cuatro parejas y se trata ahora de determinar qué niño pertenece a cada una, a saber: a. AB x O. b. A x O. c. A x AB. d. O x O. Determine qué niño corresponde a cada pareja La pareja d) sólo pueden ser padres del niño 0 El niño AB tiene que ser hijo de la pareja c) De los que quedan, la pareja b) tienen que ser padres del niño A El B es de la pareja a)
EL DOGMA CENTRAL -‐ Generalidades: -‐ En organismos eucariotas, el hecho de que el ADN esté en el núcleo para protegerlo, complica mucho la transferencia de información al citoplasma, ya que la transcripción tiene lugar en el núcleo y la traducción en el citoplasma. -‐ El ARNm es el encargado de realizar esta transferencia. el ADN se transcribe en una copia de ARN, que tras madurar, saldrá al citoplasma en forma de ARNm. Una vez allí, junto con ribosomas y ARNt se podrá llevar a cabo la traducción del ARN a proteínas. -‐ La estructura química del ARN es similar a la del ADN salvo que tiene un azúcar de ribosa en lugar de desoxirribosa. Además, el uracilo (U) remplaza a la timina. Este cambio impide la formación de estructuras superiores. El ARN de la mayoría de organismos suele encontrarse en cadena simple. -‐ Definición: Se conoce como dogma central a las relaciones de información entre el ADN, ARN y las proteínas: el ADN controla la síntesis y secuencia del ARN y éste último, a su vez, controla la síntesis y secuencia de las proteínas CARACTERÍSTICAS ESTRUCUTRALES DE UN GEN HUMANO TÍPICO -‐ Definición molecular: Un gen es una secuencia de ADN necesaria para la elaboración de un producto funcional, ya sea un polipéptido o una molécula de ARN.
codificantes, los propios genes también incluyen otras secuencias de nucleótidos adyacentes necesarias para la adecuada expresión del gen; los elementos o zonas reguladoras. -‐ Elementos reguladores: -‐ En cada tejido concreto se expresa un subgrupo de genes del genoma. Los distintos promotores van a ser lo que especifiquen qué genes se van a expresar en cada tejido. -‐ Existen distintas zonas reguladoras de la expresión génica, como por ejemplo: el promotor región 5’; La cola de poli A, en la región 3’, e incluso los intrones: -‐ La región promotor del extremo 5’, tiene secuencias para que se dé el inicio de la transcripción. -‐ En el extremo 3’ de cada gen se encuentra una importante región no traducida que contiene una señal para añadir una secuencia de residuos de adenosina (cola poli A) al extremo del RNA maduro. -‐ Tanto la región promotora del extremo 5’ como la cola de poliadenina tienen como principal función estabilizar y proteger la información genética, y no son traducidas. -‐ Existen distintos niveles de regulación: Potenciadores (o enhancers), silenciadores y regiones de control. -‐ Algunos elementos reguladores pueden estar situados en posiciones alejadas de la secuencia codificadora.
FUNDAMENTOS DE LA EXPRESIÓN GÉNICA: TRANSCRIPCIÓN -‐ Definición: es el conjunto de procesos por el cual se produce el paso de una secuencia de ADN (hebra molde) a una secuencia de ARN mensajero, transferente o ribosómico, usando siempre para ello, ribonucleótidos trifosfatados (de A,C,G,U), que proporcionarán, al romper dos de sus enlaces fosfato, la energía necesaria para el proceso. -‐ Síntesis del transcrito primario de ARN: se desarrolla en sentido 5’ a 3’, mientras que la cadena del gen que está siendo transcrito se lee en sentido 3’ a 5’, del producto final de ARN. Teniendo la cadena codificante puedo saber qué ARN voy a tener. -‐ Los elementos cis son secuencias específicas del ADN que regulan la transcripción de uno o más genes. -‐ Los factores que actúan en trans son proteínas que reconocen y se unen (directa o indirectamente) a los elementos cis para regular su actividad. -‐ ARN polimerasas: Los organismos eucariotas tienen 3 clases de ARN polimerasas, cada una de ellas compuestas por diferentes subunidades y que son responsables de la transcripción de diferentes ARN: -‐ ARN pol I: Transcribe uno de los tipos del ARN ribosómico. No se une al promotor directamente. -‐ ARN pol II: es un complejo multienzimático de 12 subunidades. Se trata de una nucleotidil transferasa capaz de añadir, en ausencia de cebador, un nuevo residuo a un ribonucleótido en su extremo 3'. -‐ Lleva a cabo la transcripción, es la más abundante. -‐ Se asocia con 6 factores de transcripción TFII A -‐ TFII H. -‐ Roger Kornberg consiguió el Nobel en 2006 por desvelar su estructura. -‐ ARN pol III: transcribe el resto del ARN. -‐ Las ARN polimerasa requiere varios cofactores que se unen al ADN en el momento de iniciarse la transcripción llamados también factores de transcripción. -‐ La amanitina es un inhibidor específico de la ARN pol. -‐ Fases de la transcripción: Se distinguen 3 fases en el proceso: -‐ Iniciación. En este primer paso, la síntesis de ARN no ocurre al azar, sino que es necesaria la presencia de un promotor y de regiones reguladoras. Aquí se unirá la ARNpol II y comenzará la síntesis. Es muy importante que el promotor no interaccione con nada pues quedaría bloqueado y no se daría la síntesis. -‐ Elongación, cuando la enzima se ancla, se irá desplazando por la hebra de ADN en sentido 3-‐ 5, mientras que la cadena de ARNm se irá formando antiparalelamente, en sentido 5-‐ 3. Se trata de un rápido proceso. -‐ La mayor parte de los FT se “sueltan” de la ARN-‐pol al iniciar la fase de elongación. -‐ Cuando se han transcrito unas 30 bases del gen, al ARNm en formación se le añade en el extremo 5una “caperuza” de metil-‐guanosina, unida por un grupo trifosfato (extremo CAP). -‐ **Finalización**. Termina con el codón STOP. La terminación de la transcripción ocurre en la secuencia AATAAA seguida de repeticiones de GT. -‐ Ocurre de forma coordinada con las modificaciones postranscripcionales. -‐ Tras esto, actúa otra enzima, la poli-‐A-‐polimerasa, que adiciona en el extremo 3 la “cola poli-‐ A”.