¡Descarga guia tema 23 y más Guías, Proyectos, Investigaciones en PDF de Genética solo en Docsity!
Epigenética
L
as células somáticas del cuerpo humano contienen de 20.000 a 25.000 genes. Sin embargo, en cualquie- ra de estas células, solo un porcentaje relativamente pequeño de todos los genes está activo. En los más de 200 tipos de células diferentes presentes en el cuer- po, se transcriben distintos conjuntos de genes específicos de cada célula, mientras que el resto del genoma es transcrip- cionalmente inactivo. Además, los programas de expresión génica se hacen cada vez más restrictivos durante el desarro- llo y la diferenciación, a medida que las células embrionarias se convierten gradualmente en células adultas especializa- das, con fenotipos diferenciados. La visión predominante ha sido que la regulación de la expresión génica está coordina- da por los promotores, promotores proximales, intensifica- dores y otros elementos reguladores de actuación en cis, así como por proteínas de unión al ADN y factores de trans- cripción. Esta regulación puede tener lugar en cualquiera de los pasos de la expresión génica. Por ejemplo, las fases de la regulación transcripcional, así como el procesamiento del ARNm y otros estadios de la regulación postranscripcional, controlan la cantidad de producto génico sintetizado a par- tir de un molde de ADN. Sin embargo, a medida que apren- demos más cosas acerca de la organización del genoma y de la regulación de la expresión génica, queda claro que los me- canismos genéticos clásicos no pueden explicar cómo surgen algunos fenotipos. Por ejemplo, los gemelos monocigóticos tienen genotipos idénticos, pero no siempre son fenotípica- mente idénticos. Aunque para todo gen, un alelo se hereda de la madre y el otro del padre, en algunos casos solo se ex- presa el alelo materno o el paterno, mientras que el otro es transcripcionalmente silencioso. El nuevo campo emergente de la epigenética nos está proporcionando las bases para comprender cómo pueden influir en la variación fenotípica ciertos cambios heredita- rios, distintos de los determinados por la secuencia de ADN. Estos avances amplían enormemente nuestro conocimiento sobre las bases moleculares de la regulación génica y se apli- can a un amplio rango de áreas, entre las que se incluyen las enfermedades genéticas, el cáncer y el comportamiento. Un carácter epigenético es un fenotipo estable, mitóti- ca y meióticamente, heredable, que resulta de cambios en la
CAPÍTULOS ESPECIALES
SOBRE GENÉTICA MODERNA
C A P Í T U L O E S P E C I A L 3
El nuevo campo emergente de la epigenética nos está proporcionando las bases para comprender cómo pueden influir en la variación fenotípica ciertos cambios hereditarios, distintos de los determinados por la secuencia de ADN.
CAPÍTULO ESPECIAL 3 ● EPIGENÉTICA 571
expresión génica que tienen lugar sin que se produzcan alte- raciones en la secuencia del ADN. La epigenética es el estu- dio de las formas en que estos cambios alteran los patrones de la expresión génica específicos de cada célula y cada te- jido. La regulación epigenética de la expresión génica utili- za modificaciones reversibles del ADN y de la estructura de la cromatina para mediar la interacción del genoma con una diversidad de factores ambientales, y para generar cambios en los patrones de expresión génica en respuesta a estos fac- tores. El epigenoma es el término con el que se designa al es- tado epigenético de una célula. Durante su tiempo de vida, un organismo tiene un solo genoma, pero este genoma pue- de modificarse en los diversos tipos de células en diferentes instantes temporales, para producir muchos epigenomas. Los actuales esfuerzos de investigación se centran en va- rios aspectos de la epigenética: cómo surge un epigenoma durante el desarrollo y la diferenciación celular, qué meca- nismos mantienen estos estados y cómo se transmiten a tra- vés de la mitosis y la meiosis, convirtiéndose en caracteres hereditarios. Además, puesto que ciertas alteraciones del genoma controladas epigenéticamente están asociadas con el cáncer y con algunas enfermedades comunes como la dia- betes y el asma, los esfuerzos de investigación también se es- tán dirigiendo hacia el desarrollo de fármacos que puedan modificar o invertir en las células los cambios epigenéticos asociados con enfermedades. Se han descubierto varios sistemas y rutas que dan lu- gar al establecimiento, mantenimiento y herencia del estado epigenético. Estas rutas están organizadas en tres categorías, aunque indudablemente quedan por elucidar otros meca- nismos y categorías. La primera categoría incluye las seña- les ambientales denominadas generadores epigenéticos que la célula recibe y que estimulan una respuesta a través de una ruta intracelular. Las respuestas a las señales generado- ras se denominan iniciadores epigenéticos. Los componentes de esta segunda categoría producen cambios epigenéticos. Estos iniciadores incluyen rutas de transducción de seña- les proteína-proteína, proteínas de unión al ADN y ARN no codificante. Las acciones de los iniciadores definen la localización en la que tienen lugar los cambios epigenéti- cos en la cromatina. El reconocimiento de la secuencia de ADN es una parte necesaria de esta respuesta. Una vez que las modificaciones epigenéticas se han producido, estas son mantenidas por unos elementos moleculares denominados mantenedores epigenéticos. Los componentes de esta tercera categoría conservan y mantienen las modificaciones epige- néticas en la generación actual y en las futuras. Estos man- tenedores no son específicos de la secuencia, operan en cualquier lugar del genoma y dependen de los iniciadores para especificar los loci en los que tendrán lugar las modifi- caciones de la cromatina. Además, los mantenedores garan- tizan que las modificaciones epigenéticas sean transmitidas
por mitosis a las células hija, o por meiosis a los gametos y a las subsiguientes generaciones. Entre los mantenedores se incluyen la metilación del ADN y las modificaciones de las histonas. La Figura CE3.1 muestra estos componentes de la ruta epigenética, junto con algunos ejemplos. Debe ob- servarse que las modificaciones de las histonas forman par- te del proceso general de activación transcripcional; esto significa que algunas de las modificaciones de las histonas, pero no todas, son sucesos epigenéticos. La modificación epigenética del genoma proporciona una respuesta clara y definida a las señales ambientales, median- do cambios en la expresión génica que persisten a lo largo de varias generaciones. Se ha determinado que la epigenética in- fluye en muchos aspectos de la biología, incluyendo la restric- ción progresiva de la expresión génica durante el desarrollo, la expresión específica de los alelos típica de la impronta ge- nética y las interacciones entorno-genoma durante el desarro- llo prenatal que afectan a los fenotipos adultos. La regulación anormal del epigenoma lleva a trastornos genéticos humanos, como el síndrome de Prader-Willi, el síndrome de Angelman y el síndrome de Beckwith-Weidemann. La pérdida o altera- ción de otros estados epigenéticos puede resultar en cáncer. El conocimiento de los mecanismos de las modificacio- nes epigenéticas del genoma, de cómo estas modificaciones se mantienen y transmiten y de sus relaciones con los pro- cesos biológicos básicos, son importantes para comprender el desarrollo, los procesos de las enfermedades, la reproduc- ción y la evolución de la adaptación al entorno.
Transducción de señal
Receptor
Proteínas de unión al ADNARN pequeños Citoplasma
Generadores
Iniciadores
ADN metilasas Histona acetilasas Histona deacetilasas
Mantenedores
Núcleo Genes activados
Genes silenciados
Carcinógenos Radiación UV Inflamación
Estrés Nicotina Infección
FIGURA CE3.1 Una ruta epigenética implica la recepción y el procesamiento de señales externas que resultan en alteraciones que cambian y mantienen los patrones de expresión génica.
CAPÍTULO ESPECIAL 3 ● EPIGENÉTICA 573
genes de estos nucleosomas modificados estén disponibles para la transcripción [ Figura CE3.4 (a)]. Esta modificación es reversible y los grupos acetilo pueden ser eliminados por otra enzima, la histona deacetilasa (HDAC), cambiando la modificación de la cromatina a «cerrada» y silenciando los genes, al hacerlos inaccesibles para la transcripción [Figu- ra CE3.4(b)]. Sin embargo, el programa de modificación de las histonas depende de otros sucesos, como la presencia o ausencia de grupos metilo en las histonas. El programa de modificación aminoacídica es complejo y a menudo el mismo tipo de modificación puede tener resul- tados transcripcionales opuestos. Por ejemplo, la metilación de la lisina 4 o 27 de la histona H3 da lugar a la activación de la transcripción, mientras que la metilación de la lisina 9 da lugar al silenciamiento de la transcripción. Se ha descubier- to que combinaciones específicas de modificaciones de las histonas controlan el estado transcripcional de una región
la replicación de elementos transponibles tales como las se- cuencias LINE y SINE (véase el Capítulo 12), que constitu- yen una fracción muy importante del genoma humano. La metilación heterocromática también mantiene la estabili- dad del cromosómica, impidiendo la translocación y otras anormalidades cromosómicas. Como parte de la compensación de dosis génica, los cromosomas X de las hembras de mamífero se inactivan, convirtiéndolos en heterocromatina. Estos cromosomas in- activados tienen patrones alterados de metilación del ADN (consulte el Capítulo 7 para ver una explicación detalla- da sobre la inactivación del cromosoma X). Como hemos mencionado anteriormente, la metilación CpG en las regio- nes eucromáticas da lugar a un patrón de transcripción gé- nica específico del progenitor.
Modificación de las histonas
Además de la metilación del ADN, la modificación de las histonas constituye un importante mecanismo epigenético de regulación génica. Recuerde que la cromatina está com- puesta por ADN espiralizado alrededor de un octámero de proteínas histonas, para formar nucleosomas. Los aminoá- cidos de la región N-terminal de estas histonas puede ser modificados covalentemente de varias formas, entre las que se incluyen la acetilación, la metilación y la fosforilación ( Figura CE3.3 ). Estas modificaciones se producen en se- cuencias aminoacídicas conservadas en las colas de la re- gión N-terminal de las histonas, las cuales sobresalen del nucleosoma. La modificación química de las histonas altera la estructura de la cromatina, haciendo accesibles o inacce- sibles los genes para la transcripción. Normalmente, cuando las histonas son modificadas por acetilación, una reacción catalizada por la enzima histona acetiltransferasa (HAT), la estructura de la cromatina «se abre», haciendo que los
(a)
(b)
Isla CpG Gen
El promotor no está metilado y el gen se puede transcribir.
El promotor está metilado y el gen está silenciado.
Isla CpG
Isla CpG no metilada Isla CpG metilada
Gen
FIGURA CE3.2 Las islas de CpG suelen localizarse corriente arriba de las regiones promotoras.
ADN
Me
P Ac
Ac
Histonas
FIGURA CE3.3 Los nucleosomas, que son agrupaciones de histonas, son el foco de las modificaciones epigenéticas. Ac = grupos acetilo, Me = grupos metilo, P = grupos fosfato.
574 LOS CONCEPTOS DE LA GENÉTICA
de ARNm, el ARNm se corta y se destruye, silenciando de forma efectiva al gen. Recientemente se ha descubierto que ARNsi pueden si- lenciar a los genes interfiriendo directamente con la inicia- ción de la transcripción. Esto no implica ningún cambio en las modificaciones epigenéticas del promotor ya existentes, ni tampoco requiere nuevas modificaciones. En su lugar, ARNsi complementarios de regiones promotoras se unen a un promotor. La unión bloquea el ensamblaje del comple- jo de preiniciación impidiendo la unión del factor de trans- cripción TFIIB y de la ARN polimerasa. Las moléculas de ARNsi también pueden asociarse con complejos proteicos para formar complejos de silenciamien- to transcripcional inducidos por ARN (RITS, RNA-Indu- ced Transcriptional Silencing). Los complejos RITS inician la formación de heterocromatina facultativa que silencia los genes localizados dentro de estas regiones heterocromáticas recién creadas. A diferencia de la heterocromatina de los te- lómeros y centrómeros, que es constitutiva, el estado hete- rocromático de la heterocromatina facultativa es reversible y puede convertirse en eucromatina, estando los genes de esta región de nuevo accesibles para la transcripción. En resumen, las modificaciones epigenéticas alteran la estructura de la cromatina, sin cambiar la secuencia de ADN, mediante varios mecanismos, entre los que se inclu- yen la metilación del ADN, la acetilación de histonas y la in- terferencia del ARN. Estos cambios epigenéticos crean un epigenoma que a su vez puede regular el desarrollo normal o generar respuestas a señales ambientales.
de cromatina. Por ejemplo, el que la lisina 9 de la histona H esté o no metilada está controlado por modificaciones reali- zadas en otros lugares de esta proteína. Por un lado, si se fos- forila la serina 10, se inhibe la metilación de la lisina 9. Por el contrario, si la lisina 14 se desacetila, se facilita la metila- ción de la lisina 9. La suma de los complejos patrones e in- teracciones de las modificaciones de las histonas que alteran la organización de la cromatina y la expresión génica se co- noce como código histónico. Los detalles de cómo funcio- na este nivel de regulación están empezando a vislumbrarse, pero todavía no se comprenden completamente.
Interferencia del ARN
Además de la metilación del ADN y de la modificación de las histonas, las moléculas de ARN pequeño no codi- ficante (tema del que hemos hablado en el Capítulo 17) también participan en la regulación epigenética de la ex- presión génica. Después de la transcripción, estas molécu- las de ARNsi (ARN pequeño de interferencia) se asocian con complejos proteicos para formar complejos de silen- ciamiento inducidos por ARN (RISC, RNA-Induced Silen- cing Complexes). Los complejos RISC se unen a moléculas de ARNm que transportan secuencias complementarias de ARNsi contenidos en los RISC. Si ARNsi no son perfecta- mente complementarios de los ARNm, la unión interfiere con la traducción, dando lugar a una sub-regulación de la expresión génica. Sin embargo, si los ARNsi del complejo RISC son perfectamente complementarios de las secuencias
Configuración «abierta». El ADN no está metilado y las histonas están acetiladas. Los genes pueden ser transcritos.
Configuración «cerrada». El ADN está metilado en las islas CpG (círculos negros) y las histonas están deacetiladas. Los genes no pueden ser transcritos.
(b)
(a)
Ac Ac Ac Ac
H1 H1 H
FIGURA CE3.4 Modificaciones epigenéticas del genoma.
576 LOS CONCEPTOS DE LA GENÉTICA
y no codificante, que es un represor del crecimiento. La ex- presión de estos genes suele estar controlada por una región de control de impronta (ICR) localizada dentro de su domi- nio cromosómico. La impronta materna anormal en uno o ambos domi- nios puede llevar a desarrollar el síndrome de Beckwith– Wiedemann, pero muchos individuos afectados tienen una pérdida de impronta del alelo materno IGF2. Esto hace que tanto el alelo materno como el paterno sean transcripcio- nalmente activos, lo que lleva al sobrecrecimiento tisular ca- racterístico de esta enfermedad. El número de genes impresos conocido solo representa una fracción pequeña del genoma, pero desempeñan fun- ciones importantes en el control del crecimiento durante el desarrollo embrionario y prenatal. Puesto que actúan en una etapa tan temprana de la vida, los factores internos o externos que perturban el patrón epigénetico de la impron- ta o la expresión de los genes impresos, pueden tener con- secuencias fenotípicas serias. Después de la introducción de la fecundación in vi- tro (IVF) y de la clonación de embriones en la producción de animales de granja, se ha observado una alta inciden- cia de nacimientos de individuos con defectos y de muer- te de recién nacidos. Algunos estudios han demostrado que muchos de estos problemas estaban relacionados con alte- raciones epigenéticas anormales durante el crecimiento y el desarrollo embrionarios. Estos hallazgos en animales apun- taron la posibilidad de que el uso de IVF y de otras tecno- logías de reproducción asistida (ART) en humanos puedan provocar problemas con los genes impresos. Diversos estudios han demostrado que los niños nacidos gracias a la fecundación in vitro tienen el riesgo de nacer con un peso bajo o muy bajo, una condición que puede ser cau- sada por una impronta anormal. También el uso de IVF y de otros procedimientos de reproducción asistida se ha asocia- do con un riesgo de tres a seis veces mayor de sufrir el síndro- me de Beckwith-Wiedemann (BWS). Uno de estos estudios examinó el epigenoma de niños con BWS que habían nacido gracias a un proceso denominado inyección intracitoplasmá- tica de espermatozoides (ICSI), en el que se inyecta esperma en el oocito. Los resultados mostraron que la mayoría de es- tos niños presentaban cambios específicos tanto en los patro- nes de metilación impresos, como en los epigenéticos. Otros estudios han demostrado que más del 90 por ciento de los niños nacidos con BWS después de un proceso de ART pre- sentan defectos de impronta. Dado que los trastornos de im- pronta no son comunes (BWS se produce con una frecuencia de aproximadamente 1 de entre 15.000 nacimientos), será necesario llevar a cabo estudios a gran escala y longitudina- les para poder llegar a establecer una relación causal entre las anormalidades de la impronta, los trastornos de crecimiento y los procedimientos de reproducción asistida.
mientras que los genes silenciados por metilación epigené- tica pueden ser reactivados por señales externas durante o después de la diferenciación. La mayoría de las enfermedades de la especie humana asociadas con la impronta tienen sus orígenes durante el cre- cimiento y el desarrollo fetales. Los defectos de impronta causan el síndrome de Prader-Willi, el síndrome de Angel- man, el síndrome de Beckwith-Wiedemann y otras enferme- dades ( Tabla CE3.1 ). Sin embargo, dado el número de genes candidatos y la posibilidad de que se descubran genes impre- sos adicionales, el número total de enfermedades genéticas relacionadas con la impronta puede ser mucho más alto.
TABLA CE3.1 Algunos trastornos de la impronta en los seres humanos Enfermedad Locus Osteodistrofia hereditaria de Albright Síndrome de Angelman Síndrome de Beckwith-Wiedemann Síndrome de Prader-Willi Síndrome de Silver-Russell Disomía uniparental 14
20q 15q11-q 11p 15q11-q Cromosoma 7 Cromosoma 14
En los seres humanos, la mayoría de los genes impresos co- nocidos codifican factores de crecimiento u otros genes que regulan el crecimiento. Generalmente, los alelos expresados maternalmente de los genes impresos suprimen el crecimien- to, mientras que los alelos paternos potencian el crecimien- to. Una enfermedad dominante autosómica de la impronta, el síndrome de Beckwith-Wiedemann (BWS), proporciona in- formación sobre como los desajustes de los genes impresos epi- genéticamente conducen a un fenotipo anormal. BWS es una enfermedad prenatal de sobrecrecimiento con defectos en las paredes abdominales, órganos agrandados, un peso excesivo al nacer y predisposición al cáncer. BWS no está causada por una mutación ni está asociada con ninguna aberración cromosómi- ca, sino que es un trastorno debido a la impronta que está cau- sado por la presencia de patrones de metilación anormales. Los genes relacionados con el BWS están localizados en una agrupación de genes impresos ubicada en el bra- zo corto del cromosoma 11. Esta agrupación contiene más de una docena de genes impresos, algunos de ellos expre- sados paternalmente y otros maternalmente, encargándose todos ellos de regular el crecimiento durante el desarrollo prenatal. La región impresa se subdivide en dos dominios regulados por separado, conteniendo uno de ellos los genes estrechamente ligados IGF2 (factor de crecimiento de la in- sulina 2) y H19. Normalmente, se expresa el alelo paterno de IGF2 y el alelo del homólogo materno se imprime y se silencia. En el caso del H19, la situación suele ser la inver- sa: el alelo materno se expresa y el alelo paterno se impri- me y silencia. La proteína codificada por IGF2 es un factor de crecimiento y el producto del gen H19 es un ARN largo
CAPÍTULO ESPECIAL 3 ● EPIGENÉTICA 577
otros cambios epigenéticos, incluyendo la hipermetilación selectiva y el silenciamiento de genes, también están presen- tes en las células cancerosas. Actualmente, el cáncer se con- templa como una enfermedad que implica tanto cambios epigéneticos como genéticos, que conducen a alteraciones en la expresión génica ( Figura CE3.6 ). La hipometilación del ADN invierte la inactivación de los genes, llevando a una transcripción no restringida de muchos grupos de genes, incluyendo los oncogenes. Tam- bién relaja el control sobre los genes impresos, haciendo que las células adquieran nuevas propiedades de crecimiento. La hipometilación de las secuencias de ADN repetitivas en las regiones heterocromáticas está asociada con un incremen- to de las reorganizaciones cromosómicas y de los cambios en el número de cromosomas, cuestiones ambas que son características de las células cancerosas. Además, la hipo- metilación de secuencias repetitivas conduce a la activación transcripcional de secuencias de ADN transponibles tales como las secuencias LINE y SINE, incrementando aun más la inestabilidad genómica. Aunque la hipometilación generalizada es un distintivo de las células cancerosas, la hipermetilación en las islas CpG y la inactivación de ciertos genes, incluyendo los genes supre- sores de tumores ( Tabla CE3.2 ), también se encuentran en muchos cánceres, a menudo según un patrón específico de cada tumor. Por ejemplo, el BRCA1 se hipermetila e inactiva en el cáncer de mama y de ovario, y el MLH1 se hipermetila en algunas formas de cáncer de colon. Se cree que la inacti- vación por hipermetilación de los genes supresores de tumo- res desempeña un papel importante y complementario de los cambios mutacionales que acompañan a la transformación de las células normales en células malignas. Por ejemplo, en una línea celular de un cáncer de vejiga, un alelo del gen de control del ciclo celular CDKN2A muta y el otro, el alelo nor- mal, queda inactivado por hipermetilación de su isla CpG. La
Epigenética y cáncer
Los estudios epidemiológicos investigan el papel de los facto- res ambientales en la variación fenotípica normal y también como factores de riesgo en el desarrollo de enfermedades. En muchos casos, es difícil obtener medidas cuantitativas de los efectos de las influencias ambientales multidimensiona- les sobre un organismo. En el caso de algunas enfermedades complejas, existen vínculos fuertes con factores ambienta- les, como por ejemplo la asociación entre el tabaco y el cán- cer de pulmón. El descubrimiento de que la epigenética hace de mediador en el proceso de modificación de los patro- nes de expresión génica en respuesta a señales ambientales ofrece un nuevo método potencialmente más directo, para comprender las interacciones existentes entre el genoma y el entorno en enfermedades como el cáncer. Después del descubrimiento de genes asociados con el cáncer, incluyendo los genes supresores de tumores y los pro- tooncogenes, las investigaciones sobre la base genética del cáncer se centraron principalmente en los alelos mutantes de genes implicados en diversas funciones celulares, incluyen- do el ciclo celular, la diferenciación, la adhesión y la apopto- sis. Hasta hace poco tiempo, el punto de vista convencional era que el cáncer es clonal en su origen y que se inicia en una célula que ha acumulado una serie de mutaciones dominan- tes y recesivas que la permiten escapar de los mecanismos de control del ciclo celular. Las mutaciones subsiguientes per- miten a las células tumorales adquirir propiedades metás- ticas y que el cáncer se extienda a otros lugares del cuerpo, donde se crean nuevos tumores malignos. Algunas líneas de investigación convergentes están clarificando el papel que desempeñan los cambios epigenéticos en la iniciación y el mantenimiento de la malignidad. Estos hallazgos ayudan a explicar propiedades de las células cancerosas que son difíci- les de explicar únicamente por la acción de alelos mutantes. Ahora hay pruebas del papel que juegan los cambios epige- néticos en el cáncer las cuales desafían al paradigma con- vencional acerca del origen del cáncer y establecen que los cambios epigenómicos son una de las rutas principales para la formación y difusión de las células malignas. La relación entre la epigenética y el cáncer fue obser- vada por primera vez en 1980 por Feinberg y Vogelstein, quienes vieron que las células del cáncer de colon tenían ni- veles mucho más bajos de metilación que las células norma- les obtenidas del mismo tejido. Investigaciones posteriores realizadas por muchos investigadores demostraron que la hipometilación global es una propiedad de todos los cánce- res examinados hasta la fecha. En los años siguientes, quedó claro que los estados epigenéticos de las células normales se ven enormemente alterados en las células cancerosas y que
Genes supresores de tumores
Oncogenes
Desactivación
Activación
Hipermetilación de islas CpG
Mutaciones genéticas
Hipometilación global
Translocaciones cromosómicas
Inestabilidad cromosómica genómica
(a)
(b)
FIGURA CE3.6 Cáncer: cambios epigenéticos y mutacionales en el genoma.
CAPÍTULO ESPECIAL 3 ● EPIGENÉTICA 579
ser componentes importantes de los fenotipos comporta- mentales. En los ratones, dos regiones del cerebro muestran expresión preferencial de los genes de los progenitores. En una de estas regiones (el hipotálamo), la impronta especí- fica del sexo era más prominente en la descendencia feme- nina, lo que sugiere efectos derivados del progenitor en la función de esta región del cerebro en las hijas. En general, se han demostrado efectos dependientes del progenitor en más de 800 genes, lo que apoya la idea de que la impronta en diferentes regiones del cerebro puede representar una forma importante de regulación epigenética.
epigenéticas del genoma en las células cancerosas permitirán diseñar fármacos específicos más potentes dirigidos a los su- cesos epigenéticos, como forma de terapia contra el cáncer.
Epigenética y comportamiento
Un conjunto creciente de pruebas demuestra que los cam- bios epigenéticos, incluyendo las alteraciones en los patro- nes de metilación y la modificación de las histonas, pueden
Células madre Células madre con cambios epigenéticos
La célula madre comienza a dividirse
Célula madre alterada epigenéticamente con mutaciones genéticas
Crecimiento descontrolado
Metástasis
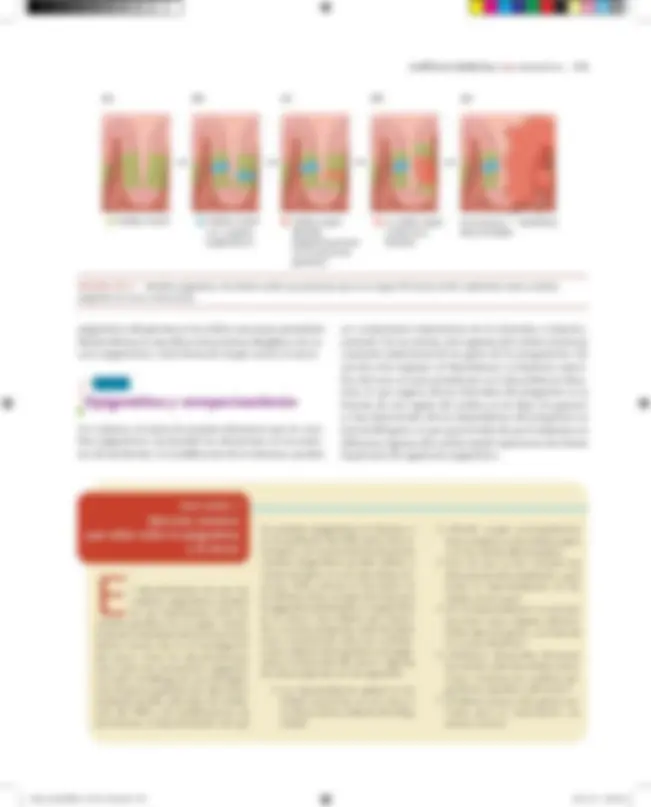
(a) (b) (c) (d) (e)
FIGURA CE3.7 Modelo epigenético de células madre que propone que en el origen del cáncer están implicados tanto cambios epigenéticos como mutaciones.
E
l descubrimiento de que los cambios epigenéticos pueden ser tan importantes como los cambios genéticos en el origen, mante- nimiento y metástasis de los cánceres ha abierto nuevas vías en la investigación del cáncer. Entre los descubrimientos clave sobre los mecanismos epigenéti- cos están el hallazgo de una desregula- ción de genes específica de cada tumor mediante perfiles alterados de metila- ción del ADN y de modificaciones de las histonas, el descubrimiento de que
los cambios epigenéticos en histonas o en la metilación del ADN están interco- nectados, y el reconocimiento de que los cambios epigenéticos pueden afectar a cientos de genes en una sola célula can- cerosa. Estos avances se han hecho en los últimos años y aunque está claro que la epigenética desempeña un papel clave en el cáncer, hay todavía que respon- der a muchas preguntas antes de poder sacar conclusiones sobre las contribu- ciones relativas de la genética y la epige- nética al desarrollo del cáncer. Algunas de estas preguntas son las siguientes: ¿La hipometilación global en las células cancerosas es una causa o un efecto de la condición de malig- nidad?
¿Dónde surgen principalmente estos cambios, en las células madre o en las células diferenciadas? Una vez que se han iniciado las alteraciones de la metilación, ¿qué activa la hipermetilación en las células cancerosas? ¿Es la hipermetilación un proceso que tiene como objetivo determi- nados tipos de genes, o se trata de un suceso aleatorio? ¿Podemos desarrollar fármacos que actúen sobre las células cance- rosas e inviertan los cambios epi- genéticos específicos del tumor? ¿Podemos actuar sobre genes con- cretos para su reactivación, sin activar a otros?
Qué más tenemos
que saber sobre la epigenética
y el cáncer
RECUADRO 2
580 LOS CONCEPTOS DE LA GENÉTICA
metilación y una transcripción reducida del gen del recep- tor GR. Los menores niveles del receptor GR no amortiguan la respuesta al estrés de estas ratas. Las ratas que recibieron cuidados adecuados tenían un 90 por ciento menos de me- tilación en un sitio de unión de factores de transcripción en la región promotora GR, permitiendo así mayores nive- les de transcripción de GR, generando más receptores GR y disminuyendo los efectos del estrés. En los adultos que ha- bían recibido menos cuidados de sus madres, los fármacos que invierten los altos niveles de metilación en el promotor GR permitían una mayor transcripción de GR. Esto, a su vez, invertía el efecto de los malos cuidados al principio de su vida y disminuía la respuesta al estrés. Trabajos posterio- res demostraron que las ratas hembra con madres más solí- citas eran más solícitas con sus propias crías, demostrando así el papel de los mantenedores epigenéticos en la transmi- sión de estos cambios a la descendencia. Después de esta investigación, otros estudios sobre ra- tas y ratones demostraron que el entorno que rodea a los animales preñados o a los recién nacidos afecta al compor- tamiento y a la salud de la descendencia en su vida adulta. Estos experimentos también demostraron que dichos cam- bios comportamentales estaban mediados por la metilación epigenética del ADN y la modificación de las histonas, que altera la configuración de la cromatina, conduciendo a mo- dificaciones en el nivel de expresión génica. La cuestión de si estos hallazgos pueden extrapolarse a los humanos ha sido fuente de algunas controversias. Se ha establecido que la adversidad en las primeras fases de la vida, como por ejemplo los malos tratos en la infancia, aumenta el riesgo de sufrir depresiones y de suicidio. ¿Puede la epigené- tica estar implicada en esto? En la especie humana, hay pocas o ninguna prueba directa de que los cambios epigenéticos en el cerebro durante etapas tempranas de la vida afecten al comportamiento posterior a lo largo de la misma. Por ejem- plo, algunos estudios demuestran que existe una correlación entre el estatus económico bajo (pobreza en la infancia) y las enfermedades del corazón en la etapa adulta. La cuestión es si los cambios epigenéticos están implicados en estas y otras situaciones similares. Dado que no es posible obtener y ana- lizar muestras del cerebro de los individuos a intervalos de tiempo distribuidos entre la infancia y la madurez, los in- vestigadores han utilizado otras fuentes de material, como la sangre del cordón umbilical y los leucocitos. Los resulta- dos obtenidos con estos tipos de células no han sido conclu- yentes en ningún caso. Que los cambios epigenéticos puedan invocarse como explicación del papel de las condiciones am- bientales en el comportamiento adulto y en las enfermedades dependerá de los resultados que se obtengan de los estudios realizados con los tejidos humanos de interés, no extrapolan- do los hallazgos obtenidos en experimentos con animales o en estudios efectuados usando tipos de células sustitutas.
En la especie humana, se han documentado cambios epi- genéticos en la evolución de trastornos neurodegenerativos y enfermedades neuropsiquiátricas con fenotipos compor- tamentales alterados. Los cambios epigenéticos en el siste- ma nervioso se han documentado en casos de la enfermedad de Alzheimer, la enfermedad de Parkinson, la enfermedad de Huntington y en enfermedades neuropsiquiátricas, entre las que se incluyen la esquizofrenia y el trastorno bipolar. Ya que la aparición de estos trastornos implica la predisposición gené- tica, sucesos en el desarrollo, efectos ambientales y la existen- cia de efectos epigenéticos transgeneracionales, ha sido difícil definir el papel preciso de los cambios epigenómicos en los trastornos neurológicos. Además, las diferentes regiones del cerebro de los individuos no afectados presentan distintos per- files epigenéticos, añadiendo una complejidad adicional a la interpretación de los resultados. Quizá el problema más desta- cable sea la dificultad de obtener tejido cerebral para su análi- sis. La mayor parte de los datos sobre alteraciones epigenéticas se han obtenido a partir de muestras de autopsias, una situa- ción que hace imposible establecer directamente una correla- ción entres estas alteraciones y el origen o evolución de estos trastornos. Sin embargo, a pesar de estas dificultades, existen bastantes pruebas que sugieren que, en el futuro, la epigenéti- ca debería también considerarse como parte del diagnóstico y tratamiento de estos complejos trastornos neurológicos. Una de las teorías relacionadas con la epigenómica que más rápidamente se está extendiendo, y quizá la más con- trovertida, tiene que ver con la idea de que hay alteraciones epigenéticas ligadas a señales ambientales, durante el desa- rrollo o en fases tempranas de la vida influyen en el compor- tamiento (y la salud física) posterior durante la vida adulta. Uno de los primeros estudios sobre epigenómica compor- tamental, publicado por Szyf y Meaney y sus colaboradores en 2004, demostró que las variaciones en el comportamien- to materno de las ratas actúan como generadores epigené- ticos, cambiando la forma en la forma en la que el cerebro de las crías responde al estrés durante la vida adulta. Los ra- tones con madres solícitas se adaptaban mejor a las situa- ciones estresantes que los ratones que no habían recibido tantos cuidados. Tanto en las ratas como en los humanos, el estrés activa el eje hipotalámico-hipofisario-adrenal (eje HHA), que amortigua la reacción al estrés aumentando los niveles de glucocorticoides en la sangre. En las ratas adultas que habían recibido cuidados ade- cuados en fases tempranas de la vida, la expresión de los receptores de glucocorticoides (GR) en las células de la re- gión del hipotálamo del cerebro se veía incrementada. Es- tos receptores detectan los niveles de glucocorticoides en la sangre y forman parte del lazo de realimentación que redu- ce la cantidad de hormonas de estrés liberadas por las glán- dulas suprarrenales. Las ratas adultas que recibieron menos cuidados durante la crianza presentan un nivel más alto de
582 LOS CONCEPTOS DE LA GENÉTICA
Atlas es uno de los programas del proyecto Roadmap Pro- ject del NIH. El atlas recopila y cataloga datos sobre un con- junto de epigenomas humanos que sirvan como estándares de referencia; se está utilizando para proporcionar infor- mación detallada acerca de las modificaciones epigenómi- cas en loci, tipos de células, estados fisiológicos y genotipos específicos. Este atlas permite a los investigadores llevar a cabo análisis integradores y comparativos de datos epige- nómicos de regiones genómicas determinadas o de geno- mas completos. Otro proyecto, denominado Proyecto del Epigenoma Humano, es un consorcio multinacional de carácter públi- co y privado que se ha creado para identificar, cartografiar y establecer el significado funcional de todos los patrones de metilación del ADN en el genoma humano. El análisis de estos patrones de metilación puede mostrar que las res- puestas genéticas a las señales ambientales mediadas por cambios epigenéticos son una ruta de aparición de enferme- dades. Aunque estos proyectos se encuentran en sus fases iniciales de trabajo, la información de la que ya se dispone apunta fuertemente a que estamos en el umbral de una nue- va era de la genética, una en la que podremos estudiar el im- pacto de los factores ambientales sobre el genoma en el nivel molecular. Los resultados de estos proyectos pueden ayudar a explicar cómo los parámetros medioambientales en fases tempranas de la vida pueden afectar a la predisposición a enfermedades en la edad adulta.
del alelo Avy^ procedentes de tejidos que representan los tres tipos de células principales del cuerpo (ectodermo, meso- dermo y endodermo) también se vieron alterados, mostran- do que la metilación en el alelo Avy^ tenía lugar en una fase temprana del desarrollo embrionario, antes de la diferencia- ción de las células madre embrionarias. Estos hallazgos tie- nen aplicación de cara a las enfermedades epigenéticas de la especie humana. Por ejemplo, el riesgo de cáncer colorrec- tal está relacionado directamente con la deficiencia de fola- to en la dieta y las diferencias de actividad en enzimas que llevan a la síntesis de donantes metilo.
Proyectos sobre el epigenoma
Vamos a terminar hablando de varios proyectos que se han puesto en marcha para cartografiar el epigenoma huma- no, incluyendo el proyecto Roadmap Epigenomics del NIH. Este proyecto está basado en la idea de que muchos aspec- tos de la salud y de la susceptibilidad a las enfermedades están relacionados con la regulación epigenética o con la in- adecuada regulación de la actividad génica. El programa se centra en cómo los mecanismos genéticos que controlan la diferenciación de las células madre y la formación de órga- nos generan respuestas biológicas a los estímulos externos e internos que provocan enfermedades. Human Epigenome
Artículos de revistas
Burdge, G.C. y Lillicrop, K.A. 2010. Nutrition, epigenetics, and deve- lopmental plasticity: implications for understanding human di- sease. Ann. Rev. Nutr. 30: 315–339. Butler, M.G. 2009. Genomic imprinting disorders in humans: a mini- review. J. Assist. Reprod. Genet. 20: 477–486. Dulac, C. 2010. Brain function and chromatin plasticity. Nature 465: 728–735. Iacobuzio-Donahue, C.A. 2009. Epigenetic changes in cancer. Ann. Rev. Pathol. Mech. Dis. 4: 229–249. Petronis, A. 2010. Epigenetics as a unifying principle in the aetiology of complex traits and diseases. Nature 465: 721–727.
Relton, C.L. y Smith, G.D. 2010. Epigenetic epidemiology of com- mon complex disease: prospects for prediction, prevention, and treatment. PLoS Medicine 7: (10) e1000356.
Sitios web National Institutes of Health Roadmap for Epigenomics. http://www. nihroadmap.nih.gov/epigenomics/initiatives.asp Human Epigenome Project. www.epigenome.org Human Epigenome Atlas. http://www.genboree.org/epigenomeatlas/ index.rhtml Computational Epigenetics Group. http://www.computational-epige- netics.de
Lecturas y recursos seleccionados