Certas espécies são naturalmente capazes de assumir o DNA exógeno (tais espécies são consideradas competentes),
incluindoHaemophilusinfluenzae,Streptococcuspneumoniae,Bacillusspp.eNeisseriaspp.Essa competênciase desenvolveaté
ofinaldafasedecrescimentologarítmico.E.colieamaioriadasoutrasbactériascarecemdeumacapacidadenaturalparaa
incorporaçãodeDNA,eessacompetênciadeveserinduzidapormétodosquímicosoueletroporação(usodepulsosdealta
tensão)parafacilitaraabsorçãodeplasmídeoseoutrasmoléculasdeDNA.
Conjugação
Aconjugação resultanatransferênciaem umúnicosentidode DNA a partir deumacéluladoadora (ou macho) a uma
receptora(oufêmea),atravésdopilisexual.Conjugaçãoocorrecomamaioria,senãotodasaseubactérias,geralmenteentre
os membros da mesma espécie ou de espécies relacionadas, mas também foi demonstrado que pode ocorrer entre
procariontesecélulasdeplantas,animaisefungos.
Otipo demating (sexo)dacélula depende dapresença(macho) ou ausência(fêmea)deum plasmídeoconjugativo,tal
comoo plasmídeoF deE. coli.OplasmídeoFé definidocomoconjugativo porquetransporta todososgenes necessários
para a sua própria transferência, incluindo a capacidade de fazerpili sexuais e iniciar a síntese de DNA na origem de
transferência(oriT)doplasmídeo.OpilussexualéumdispositivodesecreçãodotipoIVespecializado.Apósatransferência
doplasmídeoF,os receptores se tornam células‑machoF+.Seum fragmento de DNA cromossômico foiincorporadono
plasmídeo, ele édenominado plasmídeo prime F (F’). Quando ele é transferido para a célula receptora, ele carrega esse
fragmentoconsigoeconverteamesmaemumacélula‑machoF’.SeasequênciadoplasmídeoFéintegradanocromossomo
bacteriano,acélulaédenominadacélulaHfr(recombinaçãodealtafrequência).
ODNAqueétransferidoporconjugaçãonãoéumadupla‑hélice,masumamoléculadecadeiasimples.Suamobilização
começaquando umaproteínacodificada porplasmídeosfaz umaclivagemponto‑específica decadeiasimples naoriT.O
corte(nick)iniciaareplicaçãoaolongodoanel,eacadeialinearobtidaédirecionadaparaacélulareceptora.ODNAdefita
simplestransferido érecircularizadoesuafita complementarésintetizada.Aconjugação resultanatransferênciade uma
partedasequênciadoplasmídeoeuma porçãodoDNAcromossômicobacteriano.Devidoà fragilidadedaconexãoentre
asduascélulas,atransferênciaégeralmente interrompidaantes deser completadade talmodo queapenas assequências
cromossômicasadjacentesaoFintegradosãotransferidas.AinterrupçãoartificialdarelaçãoentreumparHfreF‑temsido
útilnaconstruçãodeum mapa consistente do DNA cromossômico deE. coli. Em tais mapas, a posição de cada gene é
indicadaemminutos(combasenovalorde100minutosparaatransferênciacompleta,a37°C),deacordocomoseutempo
deentradanumacélulareceptora,emrelaçãoaumaorigemfixa.
Transdução
Atransferênciagenéticaportransduçãoémediadaporvírusbacterianos(bacteriófagos),quecaptamfragmentosdeDNAe
empacotam‑nos em partículas de bacteriófagos. O DNA é entregue a células infectadas e é incorporado nos genomas
bacterianos.Atransdução podeserclassificada comoespecializada,se osfagosemquestão transferem genes específicos
(geralmenteaquelesadjacentesaosseuslocaisdeintegraçãonogenoma),ougeneralizada,seaincorporaçãodesequências
deDNA éaleatóriadevido aoempacotamentoacidentaldoDNA dohospedeirono capsídeodo fago.Porexemplo, uma
nuclease do fago P1 degrada o DNA cromossômico da E. colihospedeira, e alguns dos fragmentos do seu DNA são
empacotadosempartículasdefago.ODNAencapsulado,emvezdoDNAdofago,éinjetadoumanovacélulahospedeira,
onde pode recombinar‑se com o DNA homólogo do hospedeiro. Partículas de transdução generalizada são valiosas
nomapeamento genéticodos cromossomos bacterianos. Os doisgenesmaispróximos no cromossomo bacteriano serão,
muitoprovavelmente,cotransduzidosnomesmofragmentodeDNA.
Recombinação
AincorporaçãodeDNAextracromossômico (estrangeiro)nocromossomoocorreporrecombinação.Existemdoistiposde
recombinação: homóloga e não homóloga. A recombinaçãohomóloga (legítima) ocorre entre sequências de DNA
estreitamenterelacionadasegeralmentesubstituiumasequênciapor outra. O processo requer um conjunto de enzimas
produzidas(em E. coli) pelos genes rec. A recombinação não homóloga (ilegítima) ocorre entre sequências distintas de
DNAe, geralmente,produzinserções ou deleções,ouambas. Esteprocessogeralmente requerenzimasde recombinação
especializadas(porvezesponto‑específicas),taiscomoasproduzidaspormuitostranspósonsebacteriófagoslisogênicos.
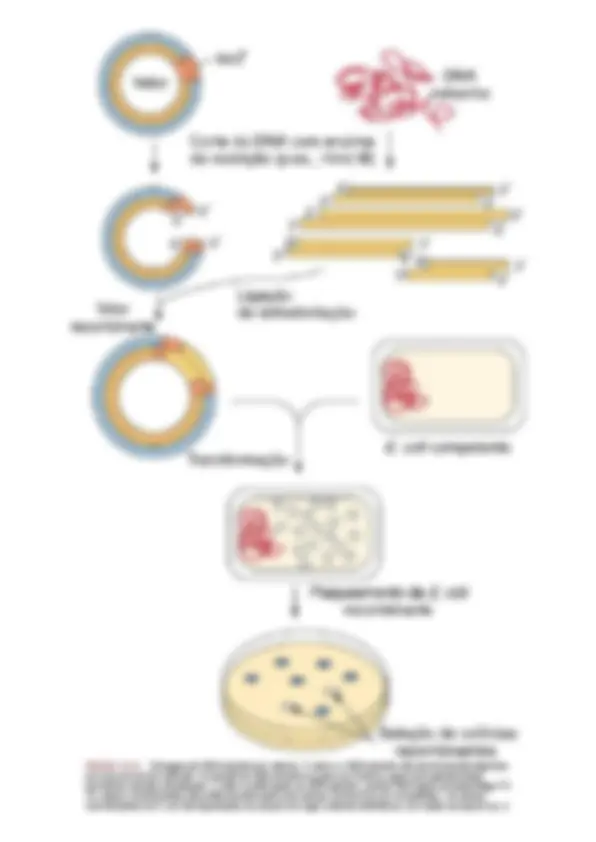
EngenhariaGenética
A engenharia genética, também conhecida como tecnologia do DNA recombinante, utiliza as técnicas e ferramentas
desenvolvidas pelos geneticistas bacterianos para purificar, amplificar, modificar e expressar sequências de genes
específicos.Ousodaengenhariagenéticaeda“clonagem”revolucionouabiologiaemedicina.Oscomponentesbásicosde
engenhariagenética são(1) vetoresde expressãoe clonagem,quepodemserusados parainserir sequênciasdeDNAem
bactériasreceptivaseamplificarasequênciadesejada;(2) asequência deDNA aser amplificadae expressa;(3) enzimas,
como enzimas de restrição, que são utilizadas para clivar o DNA de forma reprodutível em sequências definidas
(Tabela13‑1);e(4)aDNAligase,aenzimaqueligaofragmentodovetordeclonagem.