¡Descarga Inmune conceptos básicos y más Apuntes en PDF de Fisiología solo en Docsity!
INMUNE, BÁSICO
Resistencia del organismo a la infección: I. Leucocitos, granulocitos, sistema
monocitomacrofágico e inflamación
Nuestros organismos tienen un sistema especial para combatir los diferentes microorganismos infecciosos y sustancias tóxicas. Este sistema está compuesto de células blancas sanguíneas (leucocitos) y células tisulares derivadas de los leucocitos. Estas células trabajan juntas de dos formas para evitar la enfermedad: 1) destruyendo las bacterias o virus invasores mediante fagocitosis, y 2) formando anticuerpos y linfocitos sensibilizados , que, por separado o juntos, pueden destruir o inactivar al invasor. Leucocitos: son las unidades móviles del sistema protector del organismo. Se forman en parte en la médula ósea ( granulocitos y monocitos y unos pocos linfocitos ) y en parte en el tejido linfático ( linfocitos y células plasmáticas ). Tras su formación, son transportados en la sangre a diferentes partes del organismo donde son necesarios. La mayoría de ellos se transportan específicamente a zonas de infección e inflamación intensas, lo que constituye una defensa rápida y potente frente a los microorganismos infecciosos.
1. los neutrófilos polimorfonucleares. 2. los eosinófilos polimorfonucleares. 3. los basófilos polimorfonucleares.
- los monocitos.
- los linfocitos.
- en ocasiones las células plasmáticas.
- Megacariocito: Un gran número de plaquetas , que son fragmentos de otro tipo de célula similar a los leucocitos que se encuentra en la médula ósea. Nota: Los granulocitos y monocitos protegen el organismo frente a los microorganismos invasores sobre todo ingiriéndolos, es decir, mediante fagocitosis. Los linfocitos y las células plasmáticas actúan sobre todo en conexión con el sistema. Finalmente, la función de las plaquetas es en concreto activar el mecanismo de coagulación de la sangre.
Genesis de los leucocitos
Los granulocitos y los monocitos: se forman sólo en la médula ósea. Los linfocitos y las células plasmáticas: se producen, sobre todo: Estos tres primeros poseen un aspecto granulado y múltiples núcleos. Por esto se llaman granulocitos.
pueden atacar y destruir bacterias incluso en la sangre circulante. Por el contrario, los macrófagos tisulares comienzan la vida como monocitos sanguíneos, que son células inmaduras mientras están en la sangre y tienen poca capacidad de luchar contra los microorganismos infecciosos en ese momento. Pero una vez que entran en los tejidos, comienzan a aumentar de tamaño. Estas células se llaman ahora macrófagos y son muy capaces de combatir los microorganismos que están en los tejidos.
Los leucocitos entran en los espacios tisulares mediante diapédesis.
Los neutrófilos y los monocitos pueden exprimirse a través de los poros de los capilares sanguíneos por diapédesis. Es decir, aunque el poro sea mucho menor que la célula, una pequeña porción de la misma se desliza a través del poro; esta porción se constriñe momentáneamente al tamaño del poro. ¿Los leucocitos se mueven a través de los espacios tisulares por movimiento ameboide?
Los leucocitos son atraídos a las zonas de tejido inflamado mediante
quimiotaxia.
Muchas sustancias químicas diferentes en los tejidos hacen que los neutrófilos y los macrófagos se muevan hacia la fuente de las sustancias químicas. Se conoce como quimiotaxia. Cuando un tejido se inflama, se forman al menos una docena de productos diferentes que pueden producir quimiotaxia hacia la zona inflamada. Entre ellas están:
- algunas toxinas bacterianas o víricas.
- productos degenerativos de los propios tejidos inflamados.
- varios productos de reacción del «complejo del complemento» activados en los tejidos inflamados.
- varios productos de reacción causados por la coagulación del plasma en la zona inflamada, así como otras sustancias. La quimiotaxia depende de un gradiente de concentración de la sustancia quimiotáctica. La concentración es mayor cerca de la fuente, que dirige el movimiento unidireccional de los leucocitos.
Fagocitosis
Significa ingestión celular de agente ofensivo. Los fagocitos deben seleccionar el material que fagocitan; de otro modo podrían ingerir células y estructuras normales del cuerpo. El que tenga lugar la fagocitosis depende en especial de tres intervenciones selectivas.
- En primer lugar, la mayoría de las estructuras naturales en los tejidos tienen superficies lisas que se resisten a la fagocitosis, Pero si la superficie es rugosa, aumenta la probabilidad de fagocitosis.
- En segundo lugar, la mayoría de las sustancias naturales del cuerpo tiene cubiertas proteicas protectoras que repelen a los fagocitos. En cambio, la mayoría de los tejidos muertos y partículas extrañas no tiene cubiertas protectoras, lo que las hace susceptibles a la fagocitosis.
- En tercer lugar, el sistema inmunitario del cuerpo produce anticuerpos frente a los microorganismos infecciosos como las bacterias. Los anticuerpos se adhieren entonces a las membranas bacterianas y por tanto hacen a las bacterias especialmente susceptibles a la fagocitosis. Para ello, la molécula de anticuerpo se combina también con el producto C3 de la cascada del complemento, que es una parte adicional del sistema inmunitario. Las moléculas de C3 se unen a su vez a receptores situados en la membrana del fagocito, lo que inicia la fagocitosis. Esta selección y proceso de fagocitosis se llama opsonización.
Fagocitosis por los neutrófilos.
Los neutrófilos que entran en los tejidos son ya células maduras que pueden comenzar inmediatamente la fagocitosis. Al acercarse a una partícula que va a fagocitar, el neutrófilo se une en primer lugar a la partícula y después proyecta seudópodos en todas las direcciones alrededor de la partícula. Los seudópodos se encuentran entre sí en el lado opuesto y se fusionan. Esto crea una cámara cerrada que contiene la partícula fagocitada. Después la cámara se invagina hacia el interior de la cavidad citoplasmática y se separa de la membrana celular externa para formar una vesícula fagocítica (también conocida como
Sistema monocitomacrofágico (sistema reticuloendotelial)
Una gran proporción de monocitos se une a los tejidos y permanece así meses o incluso años hasta que es requerida para realizar funciones protectoras locales específicas. Tienen las mismas capacidades que los macrófagos móviles de fagocitar grandes cantidades de bacterias, virus, tejidos necróticos u otras partículas extrañas en el tejido. Y, cuando se les estimula adecuadamente, pueden romper sus inserciones y convertirse de nuevo en macrófagos móviles que responden a la quimiotaxia y a todos los otros estímulos relacionados con el proceso inflamatorio. De este modo, el organismo tiene un «sistema monocitomacrofágico» amplio en casi todos los tejidos. La combinación total de monocitos, macrófagos móviles, macrófagos tisulares fijos y unas pocas células endoteliales especializadas en la médula ósea, el bazo y los ganglios linfáticos se denomina sistema reticuloendotelial.
Macrófagos tisulares en la piel y en los tejidos (histiocitos)
Aunque la piel es prácticamente impermeable a los microorganismos infecciosos, esto no es cierto cuando la piel se rompe. Cuando la infección comienza en un tejido subcutáneo y surge la inflamación local, los macrófagos tisulares locales pueden dividirse en el mismo sitio y formar todavía más macrófagos.
Macrófagos en los ganglios linfáticos.
Prácticamente ninguna partícula que entre en los tejidos, como pueden ser por ejemplo las bacterias, puede pasar directamente a través de las membranas capilares hacia la sangre. Pero si no se destruyen las partículas que entran en los tejidos, entran en la linfa y fluyen hacia los ganglios linfáticos localizados de modo intermitente a lo largo del trayecto del flujo linfático. Las partículas extrañas quedan entonces atrapadas en estos ganglios en una red de senos recubiertos por macrófagos tisulares. Un gran número de macrófagos recubren los senos linfáticos, y si entra cualquier partícula en los senos a través de la linfa, los macrófagos la fagocitan e impiden su diseminación general por todo el cuerpo.
Macrófagos alveolares en los pulmones.
Otra vía por la que los microorganismos invasores entran con frecuencia en el cuerpo es a través de los pulmones. Hay un gran número de macrófagos tisulares formando parte integral de las paredes alveolares. Pueden fagocitar partículas que quedan atrapadas en los alvéolos. Si las partículas son digeribles, los macrófagos pueden digerirlas también y liberar los productos digeridos en la linfa. Si la partícula no es digerible, los macrófagos forman a menudo una cápsula de «células gigantes» alrededor de la partícula hasta el momento en que puedan disolverla lentamente, si es que este momento llega.
Macrófagos (células de Kupffer) en los sinusoides hepáticas. (Hígado)
Otra vía favorita por medio de la cual las bacterias invaden el cuerpo es el aparato digestivo. A través de la mucosa intestinal y hacia la sangre portal pasa constantemente un número alto de bacterias presentes en los alimentos ingeridos. Antes de que esta sangre entre en la circulación general, pasa a través de los sinusoides hepáticos, que están recubiertos de macrófagos tisulares llamados células de Kupffer. Estas células forman un sistema de filtración de partículas eficaz que hace que casi ninguna de las bacterias del aparato digestivo pase de la sangre portal a la circulación sistémica general.
Macrófagos en el bazo y en la médula ósea
Si un microorganismo invasor consigue entrar en la circulación general, hay otras líneas de defensa del sistema macrofágico tisular, especialmente los macrófagos del bazo y de la médula ósea. En estos dos tejidos, los macrófagos se quedan atrapados en la trama reticular y, cuando la partícula extraña entra en contacto con estos macrófagos, es fagocitada.
Inflamación: participación de los neutrófilos y los macrófagos
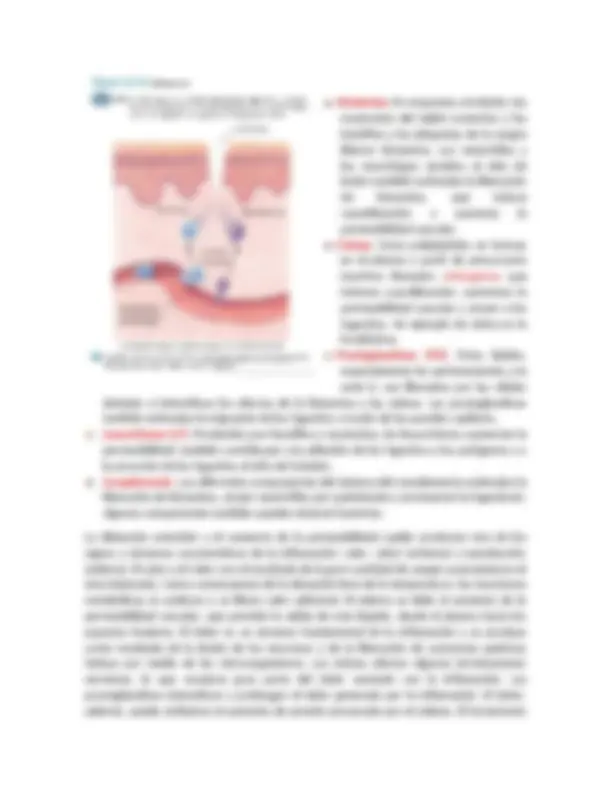
Inflamación Cuando se produce una lesión tisular, ya sea debida a bacterias, traumatismos, sustancias químicas, calor o cualquier otro fenómeno, los tejidos lesionados liberan múltiples sustancias que dan lugar a cambios secundarios espectaculares en los tejidos vecinos no lesionados. Este complejo de cambios tisulares se llama inflamación. La inflamación se caracteriza por: 1 ) La vasodilatación de los vasos sanguíneos locales, con el consiguiente exceso de flujo sanguíneo local. 1 ) La vasodilatación de los vasos sanguíneos locales, con el consiguiente exceso de flujo sanguíneo local.
- El aumento de la permeabilidad de los capilares, lo que permite la fuga de grandes cantidades de líquido hacia los espacios intersticiales. 2 ) El aumento de la permeabilidad de los capilares, lo que permite la fuga de grandes cantidades de líquido hacia los espacios intersticiales.
- A menudo la coagulación del líquido en los espacios intersticiales por un aumento en las cantidades de fibrinógeno y otras proteínas que salen de los capilares. 3 ) A menudo la coagulación del líquido en los espacios intersticiales por un aumento en las cantidades de fibrinógeno y otras proteínas que salen de los capilares.
- La migración de un gran número de granulocitos y monocitos al tejido. 4 ) La migración de un gran número de granulocitos y monocitos al tejido.
- La tumefacción ( Hinchazón que se forma en una parte del cuerpo)de las células tisulares.
- La tumefacción ( Hinchazón que se forma en una parte del cuerpo)de las células tisulares.
La invasión por neutrófilos de la zona inflamada es una segunda línea de
defensa.
Alrededor de la primera hora siguiente a la infección, un gran número de neutrófilos comienza a invadir la zona inflamada desde la sangre. Esto se debe a citocinas inflamatorias (p. ej., TNF, IL-1) y otros productos bioquímicos producidos por tejidos inflamados que inician las siguientes reacciones:
La segunda invasión de macrófagos del tejido inflamado es una tercera
línea de defensa.
Junto a la invasión de los neutrófilos, los monocitos procedentes de la sangre entran en el tejido inflamado y aumentan de tamaño hasta convertirse en macrófagos. Pero el número de monocitos en la sangre circulante es bajo: además, la reserva de monocitos en la médula ósea es mucho menor que la de neutrófilos. Luego el aumento de macrófagos en Neutrofilia : que significa aumento del número de neutrófilos en la sangre. La neutrofilia se debe a los productos de la inflamación que entran en el torrente sanguíneo, llegan a la médula ósea y allí actúan sobre los neutrófilos almacenados para movilizarlos hacia la sangre circulante. Esto deja incluso más neutrófilos disponibles para la zona tisular inflamada.
la zona del tejido inflamado es mucho más lento que el de los neutrófilos y necesita varios días para ser eficaz. Además, incluso después de invadir el tejido inflamado, los monocitos todavía son células inmaduras que necesitan 8 h o más para adquirir tamaños mucho mayores y desarrollar cantidades tremendas de lisosomas; sólo entonces adquieren la capacidad plena de los macrófagos tisulares para la fagocitosis.
La mayor producción de granulocitos y monocitos en la médula ósea es una
cuarta línea de defensa.
La cuarta línea de defensa es una mayor producción de granulocitos y monocitos en la médula ósea. Esto se debe a la estimulación de las células precursoras de granulocitos y monocitos en la médula. Pero transcurren 3-4 días antes de que los granulocitos y monocitos recién formados alcancen la fase de dejar la médula ósea. Si el estímulo procedente del tejido inflamado continúa, la médula ósea puede continuar produciendo estas células en cantidades tremendas durante meses e incluso años, a veces 20-50 veces con respecto a lo normal.
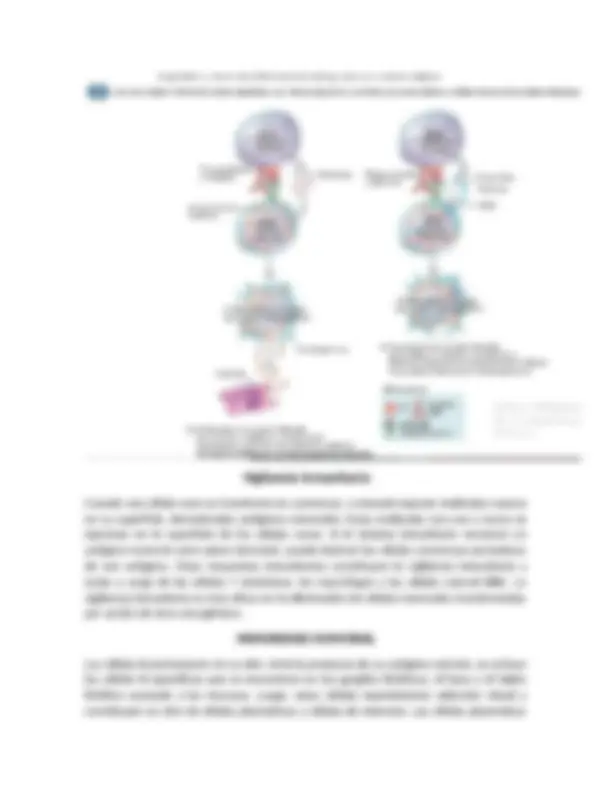
Control por retroalimentación de las respuestas del macrófago y del
neutrófilo
Las causas de esta mayor producción de granulocitos y monocitos en la médula ósea son sobre todo los tres factores estimulantes de colonias: GM-CSF, estimula la producción de granulocitos y monocitos. G-CSF y M-CSF, estimulan la producción de granulocitos y monocitos, respectivamente. Esta combinación de TNF, IL-1 y factores estimuladores de colonias constituye un mecanismo de retroalimentación poderoso que comienza con la inflamación tisular y conduce a la formación de un gran número de leucocitos defensivos que ayudan a eliminar la causa de la inflamación. Estos factores los forman los macrófagos activados en los tejidos inflamados y en menores cantidades las células tisulares inflamadas. Factores en el control de la respuesta del macrófago a la inflamación.
- El factor de necrosis tumoral (TNF).
- La interleucina 1 (IL-1).
- El factor estimulador de colonias de granulocitos-monocitos (GM-CSF).
- el factor estimulador de colonias de granulocitos (G-CSF)
- El factor estimulador de colonias de monocitos (M-CSF).
- Probablemente liberando también formas muy reactivas del oxígeno que son especialmente mortales para los parásitos.
- Liberando de los gránulos un polipéptido muy larvicida llamado proteína principal básica. Los eosinófilos también tienen una especial tendencia a acumularse en los tejidos en que se producen reacciones alérgicas, como los tejidos peribronquiales de los pulmones en las personas con asma y en la piel después de las reacciones alérgicas cutáneas. Los mastocitos y los basófilos liberan un factor quimiotáctico de eosinófilos que provoca la migración de los eosinófilos hacia el tejido con una inflamación alérgica. Se cree que los eosinófilos detoxifican algunas de las sustancias inductoras de la inflamación liberadas por los mastocitos y los basófilos y probablemente también fagociten y destruyan complejos antígeno-anticuerpo, evitando así una diseminación excesiva del proceso inflamatorio local.
Basófilos
Los basófilos que están en la sangre circulante son similares a los mastocitos tisulares grandes localizados inmediatamente por fuera de muchos de los capilares del cuerpo. Los mastocitos y los basófilos liberan heparina a la sangre, una sustancia que puede impedir la coagulación de la sangre. Los mastocitos y los basófilos también liberan histamina, así como pequeñas cantidades de bradicinina y serotonina. De hecho, son sobre todo los mastocitos de los tejidos inflamados los que liberan estas sustancias durante la inflamación. Los mastocitos y los basófilos desempeñan una función destacada en algunos tipos de reacciones alérgicas porque el tipo de anticuerpo que provoca las reacciones alérgicas, la inmunoglobulina E (IgE), tiene una tendencia especial a unirse a los mastocitos y los basófilos. Después, cuando el antígeno específico del anticuerpo IgE específico reacciona después con el anticuerpo, la unión resultante del antígeno al anticuerpo hace que el basófilo o el mastocito se rompan y liberen cantidades elevadas de histamina, bradicinina, serotonina, heparina, sustancia de reacción lenta de la anafilaxia y varias enzimas lisosómicas. Estas desencadenan reacciones vasculares locales y tisulares que a su vez provocan muchas, si no la mayoría, de las manifestaciones alérgicas.
Resistencia del organismo a la infección: II. Inmunidad y alergia. Inmunidad
innata
Inmunidad: Capacidad de resistir casi todos los tipos de microorganismos y toxinas que tienden a lesionar los tejidos y órganos.
Gran parte de ella es inmunidad adquirida que no aparece hasta que el cuerpo es
atacado por primera vez por una bacteria, un virus o una toxina, y a menudo precisa semanas o meses para desarrollarse. Una parte adicional de la inmunidad se debe a Antígeno: Sustancia que al introducirse en el organismo induce en este una respuesta inmunitaria, provocando la formación de anticuerpos.
procesos generales en lugar de a procesos dirigidos a microorganismos específicos. A esta
se le llama inmunidad innata. Comprende lo siguiente:
Inmunidad adquirida (adaptativa)
El cuerpo humano tiene la capacidad de desarrollar una inmunidad específica extremadamente potente frente a microorganismos invasores individuales como bacterias, virus y toxinas mortales, e incluso a sustancias extrañas procedentes de otros animales. La inmunidad adquirida se debe a un sistema inmunitario especial que forma anticuerpos, linfocitos activados o ambos que atacan y destruyen los microorganismos invasores específicos o las toxinas.
Tipos básicos de inmunidad adquirida: humoral y mediada por células.
Inmunidad humoral o inmunidad del linfocito B (porque los linfocitos B producen los anticuerpos): el cuerpo produce anticuerpos circulantes, que son moléculas de globulinas presentes en el plasma sanguíneo capaces de atacar al micro-organismo invasor. Inmunidad celular o inmunidad del linfocito T (porque los linfocitos activados son linfocitos T): inmunidad adquirida se consigue mediante la formación de un gran número de linfocitos T activados que se habilitan especialmente en los ganglios linfáticos para destruir el microorganismo extraño.
Los dos tipos de inmunidad adquirida los inician los antígenos
1. Fagocitosis de bacterias y otros
invasores por los leucocitos y las
células del sistema macrofágico
tisular.
2. Destrucción de microorganismos
ingeridos por las secreciones ácidas
del estómago y las enzimas
digestivas.
3. Resistencia de la piel a la invasión
por microorganismos.
4. Presencia en la sangre de ciertos
compuestos químicos que se unen a
microorganismos o toxinas extraños y
los destruyen. Algunos de estos
compuestos son:
1) La lisozima, un polisacárido
mucolítico que ataca a las bacterias y
las disuelve.
2) Polipéptidos básicos, que
reaccionan con ciertos tipos de
bacterias grampositivas y las
inactivan.
3) El complejo del complemento un
sistema de unas 20 proteínas que
puede activarse por diversas vías para
destruir las bacterias.
4) Los linfocitos asesinos naturales
que pueden reconocer y destruir
célula extrañas, células tumorales e
incluso algunas células infectadas.
Tanto los anticuerpo s como los linfocitos activados se forman en los tejidos linfáticos del
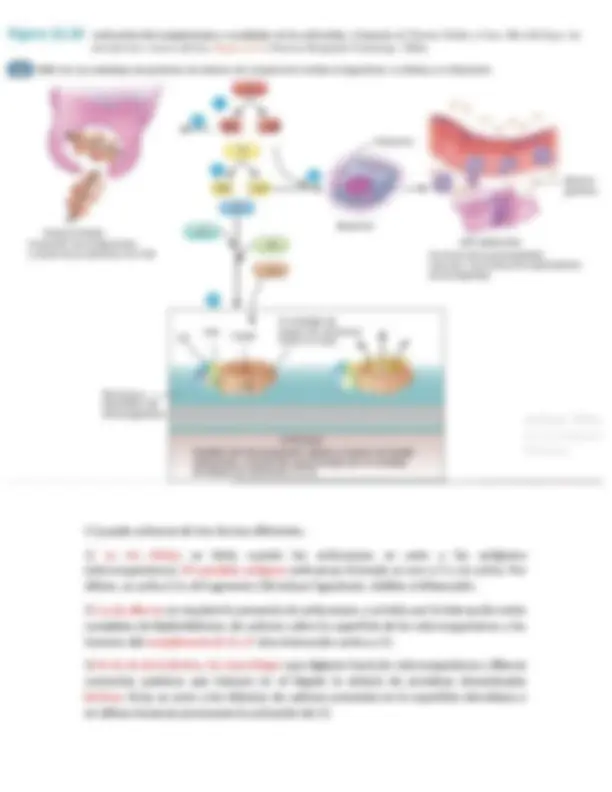
Preprocesamiento de los linfocitos T y B
Todos los linfocitos del cuerpo se originan de las células precursoras comprometidas en la línea linfocitaria del embrión, estas células progenitoras son incapaces por sí mismas de formar directamente linfocitos T activados ni anticuerpos. Antes de poder hacerlo deben diferenciarse más en zonas de procesamiento adecuadas.
El timo preprocesa los linfocitos T.
Los linfocitos T, tras originarse en la médula ósea, migran primero al timo. Aquí se dividen rápidamente y al mismo tiempo forman una diversidad extrema de capacidad de reacción frente a antígenos específicos diversos. Es decir, que un linfocito tímico desarrolla una especificidad específica frente a un antígeno. Después, el siguiente linfocito desarrolla una especificidad frente a otro antígeno. Esto continúa hasta que hay miles de tipos diferentes de linfocitos tímicos con reactividades específicas frente a muchos miles de antígenos diferentes. Estos tipos diferentes de linfocitos T preprocesados dejan ahora el timo y se diseminan a través de la sangre por todo el cuerpo para alojarse por todo el tejido linfático.
Las células progenitoras linfoides
comunes destinadas finalmente a
formar linfocitos T activados migran
primero al timo y son
preprocesados, y por ello reciben el
nombre de linfocitos «T» para
designar la función del timo.
Los linfocitos B destinados a formar
anticuerpos, es preprocesada en el
hígado durante la mitad de la vida
fetal y en la médula ósea al final de
la vida fetal y tras el nacimiento. Se
llaman linfocitos «B», para designara
la bolsa.
El timo se asegura de que los linfocitos T que abandonan el timo no reaccionen frente a proteínas u otros antígenos que estén presentes en los tejidos propios; de otro modo los linfocitos T serían mortales para la propia persona en unos días. El timo selecciona qué linfocitos T se liberarán primero mezclándolos con casi todos los «autoantígenos» de los tejidos propios del cuerpo. Si un linfocito T reacciona, es destruido y fagocitado en lugar de liberado. Luego las únicas células liberadas finalmente son las que no reaccionan con antígenos propios: sólo reaccionan frente a antígenos de una fuente externa, como una bacteria, una toxina o incluso un órgano trasplantado de otra persona.
El hígado y la médula ósea preprocesan los linfocitos B.
En el ser humano los linfocitos B se preprocesan en el hígado durante la etapa intermedia de la vida fetal y en la médula ósea durante la última etapa de la vida fetal y tras el nacimiento. Los linfocitos B son diferentes de los linfocitos T en dos aspectos: primero, en lugar de que toda la célula desarrolle la reactividad frente al antígeno, como ocurre en los linfocitos T, los linfocitos B secretan activamente anticuerpos que son las sustancias reactivas. Estas sustancias son proteínas grandes capaces de combinarse con la sustancia antigénica y de destruirla. En segundo lugar, los linfocitos B tienen una diversidad incluso mayor que los linfocitos T, con lo que forman muchos millones de tipos de anticuerpos con diferentes reactividades específicas. Tras el preprocesamiento, los linfocitos B, migran al tejido linfático de todo el cuerpo, donde se alojan cerca, pero ligeramente separados, de las zonas de los linfocitos T.
En el caso de los linfocitos T, moléculas similares a los anticuerpos llamadas proteínas receptoras de superficie (o marcadores del linfocito T ), están en la superficie de la membrana del linfocito T, y son también muy específicos de un antígeno activador específico. Por tanto, un antígeno estimula sólo aquellas células que tengan receptores complementarios para el antígeno y ya estén comprometidas para responder a él.
Función de los macrófagos en el proceso de activación.
La mayoría de los microorganismos invasores son en primer lugar fagocitados y digeridos en parte por los macrófagos, y los productos antigénicos se liberan al citosol del macrófago. Después, los macrófagos pasan estos antígenos por contacto célula a célula directamente a los linfocitos, lo que activa clones linfocitarios específicos. Además, los macrófagos secretan una sustancia activadora especial, denominada interleucina 1, que favorece un mayor crecimiento y reproducción de los linfocitos específicos.
Función de los linfocitos T en la activación de los linfocitos B
La mayoría de los antígenos activa a los linfocitos T y B al mismo tiempo. Algunos de los linfocitos T que se forman, llamados linfocitos colaboradores , secretan sustancias específicas (llamadas en conjunto linfocinas ) que activan a los linfocitos B específicos. De hecho, sin la ayuda de estos linfocitos T colaboradores, la cantidad de anticuerpos formada por los linfocitos B suele ser pequeña.
Atributos específicos del sistema del linfocito B: la inmunidad humoral y los
anticuerpos
Formación de anticuerpos por las células plasmáticas
Aquellos linfocitos B específicos frente al antígeno aumentarán de tamaño inmediatamente y adquirirán el aspecto de linfoblastos. Algunos linfoblastos se diferencian hasta formar plasmoblastos, que son los precursores de las células plasmáticas. En los plasmoblastos , el citoplasma se expande y prolifera mucho el retículo endoplásmico. Los plasmoblastos comienzan entonces a dividirse. Cada célula plasmática madura produce entonces anticuerpos gammaglobulínicos. Después los anticuerpos se secretan hacia la linfa y luego a la sangre circulante. Este proceso continúa varios días o semanas hasta que las células plasmáticas se agotan o mueren.
Formación de linfocitos de «memoria»: diferencia entre respuesta primaria
y respuesta secundaria.
Algunos linfoblastos formados por la activación de un clon de linfocitos B no forman células plasmáticas sino un número moderado de linfocitos B nuevos similares a los del Al entrar el antígeno extraño, los macrófagos del tejido linfático fagocitan el antígeno y lo presentan a los linfocitos B adyacentes.
clon original. Circulan a través del cuerpo para poblar todo el tejido linfático; pero desde un punto de vista inmunológico permanecen durmientes hasta que una nueva cantidad del mismo antígeno los activa. Estos linfocitos se llaman linfocitos de memoria. La exposición posterior al mismo antígeno dará lugar a una respuesta de anticuerpos mucho más potente y rápida esta segunda vez, porque hay muchos más linfocitos de memoria que linfocitos B originales del clon específico. Respuesta primaria para formar anticuerpos: Aparece ante la primera exposición a un antígeno específico. Débil potencia y su corta vida. Respuesta secundaria: Se produce después de la segunda exposición al mismo antígeno. Comienza rápidamente después de la exposición al antígeno (a menudo en horas), es mucho más potente y forma anticuerpos durante muchos meses en lugar de sólo unas semanas. La mayor potencia y duración de la respuesta secundaria explica por qué puede conseguirse la inmunización inyectando múltiples dosis de antígeno con períodos de semanas o meses entre las inyecciones.
Naturaleza de los anticuerpos
Los anticuerpos son gammaglobulinas llamadas inmunoglobulinas (abreviadas Ig ). Todas las inmunoglobulinas están compuestas de combinaciones de cadenas de polipéptidos pesadas y ligeras. La mayoría es una combinación de dos cadenas ligeras y dos pesadas. Pero algunas de las inmunoglobulinas tienen combinaciones de hasta 10 cadenas pesadas y 10 ligeras, lo que origina inmunoglobulinas con un peso molecular alto. Sin embargo, en todas las inmunoglobulinas cada cadena pesada lleva paralela una cadena ligera en uno de sus extremos, lo que forma parejas de cadenas pesadas y ligeras, y siempre hay al menos 2 y como mucho 10 de estas parejas en cada molécula de inmunoglobulina. La porción variable une específicamente a un tipo de antígeno en particular. La porción constante del anticuerpo determina otras propiedades del anticuerpo, lo que determina factores como la capacidad de difusión del anticuerpo en los tejidos, la adherencia del anticuerpo a estructuras específicas dentro de los tejidos, la unión al complejo del complemento, la facilidad con la que los anticuerpos atraviesan las membranas y otras propiedades biológicas del anticuerpo.