¡Descarga Lectura sobre gluconeogenesis y más Apuntes en PDF de Bioquímica Médica solo en Docsity!
8.2 Gluconeogénesis 255
8.2 GLUCONEOGÉNESIS
La gluconeogénesis, la formación de moléculas nuevas de glucosa a partir de precur-
sores que no son carbohidratos, ocurre principalmente en el hígado. Los precursores
son el lactato, el piruvato, el glicerol y determinados � -cetoácidos (moléculas que
derivan de los aminoácidos). En determinadas situaciones (p. ej., acidosis metabó-
lica o inanición) el riñón puede producir pequeñas cantidades de glucosa. Entre las
comidas se mantienen concentraciones sanguíneas adecuadas de glucosa por medio
de la hidrólisis del glucógeno hepático. Cuando se agota el glucógeno hepático (p.
ej., por un ayuno prolongado o por ejercicio vigoroso), la vía de la gluconeogénesis
proporciona al organismo la cantidad de glucosa adecuada. El cerebro y los eritroci-
tos dependen exclusivamente de la glucosa como fuente de energía.
Reacciones de la gluconeogénesis
La secuencia de reacciones de la gluconeogénesis es, en gran medida, la inversa de la
glucólisis. Sin embargo, es importante recordar que tres reacciones glucolíticas (las
reacciones catalizadas por la hexocinasa, la PFK-1 y la piruvato cinasa) son irreversi-
bles. En la gluconeogénesis, para evitar estos obstáculos, se utilizan reacciones alter-
nativas catalizadas mediante enzimas diferentes. Después se resumen las reacciones
únicas de la gluconeogénesis. En la figura 8.9 se presentan la vía gluconeogénica
completa y sus relaciones con la glucólisis. Las reacciones de circunvalación de la
gluconeogénesis son las siguientes:
1. Síntesis de PEP. La síntesis de PEP a partir de piruvato requiere dos enzimas: la piruvato carboxilasa y la PEP carboxicinasa. La piruvato carboxilasa, que se en- cuentra dentro de las mitocondrias, convierte el piruvato en oxaloacetato (OAA):
La transferencia de CO 2 para formar el producto OAA está mediada por la coen- zima biotina , que se une de manera covalente dentro del sitio activo de la enzima. El OAA se descarboxila y se fosforila por medio de la PEP carboxicinasa en una reacción impulsada por la hidrólisis del trifosfato de guanosina (GTP):
Piruvato
C
CH 3
O
C O−
O
Oxaloacetato (OAA)
C
CH 2
O
C O
O−
C O−
O
Piruvato carboxilasa (biotina)
Pi
CO 2 H 2 O
ATP ADP
OAA
C
CH 2
O
C O
O−
C O−
O
PEP
C
CH 2
O
C O−
O
P O−
O−
O
PEP
carboxicinasa
GTP GDP
CO
256 CAPÍTULO 8 Metabolismo de los carbohidratos
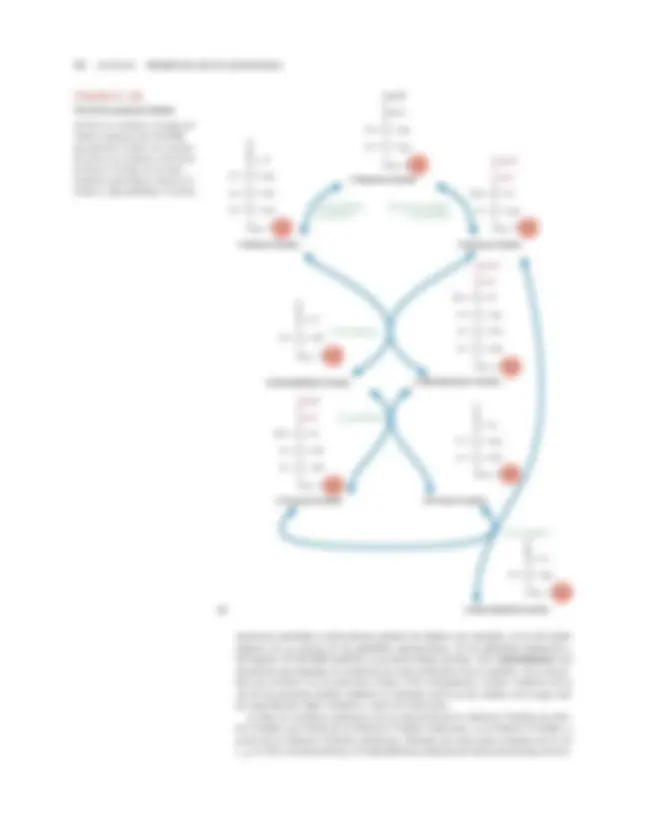
FIGURA 8.
Metabolismo de los carbohidratos: gluconeogénesis y glucólisis
En la gluconeogénesis, que tiene lugar cuando la concentración sanguínea de azúcar es baja y está agotado el glucógeno hepático,
se invierten 7 de las 10 reacciones de la glucólisis. Tres reacciones glucolíticas irreversibles se evitan mediante otras reacciones.
Los principales sustratos de la gluconeogénesis son determinados aminoácidos (que proceden de los músculos), el lactato (que se
forma en los músculos y en los eritrocitos) y el glicerol (que se produce en la degradación de los triacilgliceroles). En cambio con las
reacciones de la glucólisis, que sólo ocurren en el citoplasma, las reacciones de la gluconeogénesis catalizadas por la piruvato carboxilasa
y en algunas especies por PEP carboxicinasa se producen en las mitocondrias. La reacción catalizada mediante glucosa-6-fosfatasa ocurre
en el retículo endoplásmico. Nótese que la gluconeogénesis y la glucólisis no suceden al mismo tiempo. En la glucólisis, el piruvato se
convierte en acetil-CoA (no se muestra) o en lactato.
Glucosa
Glicerol
Gliceraldehído-3-fosfato Fosfato de dihidroxiacetona
Glicerato-1,3-difosfato
Glicerato-3-fosfato
Pi
Determinados aminoácidos
Glicerato-2-fosfato
Fosfoenolpiruvato
Lactato
Piruvato
Piruvato carboxilasa
Piruvato cinasa PEP carboxicinasa
Oxaloacetato
Determinados aminoácidos
Glucosa-1-fosfato
Glucógeno UDP-glucosa
Glucosa-6-fosfato
Fructosa-1,6-difosfato
Fructosa-6-fosfato
Hexocinasa
Glucosa-6-fosfatasa
PFK-
Fructosa difosfato fosfatasa
Pi H 2 O Pi
PPi
H 2 O H 2 O
H 2 O
CO 2
CO 2
ATP Pi ADP
UTP
NAD
NAD
NADH
GDP
GTP
NADH
NADH NADH +
NAD
NAD
ADP
ADP ADP
ADP
ADP
ATP
ATP ATP
ATP
ATP
H+ H+
H+^ H+
258 CAPÍTULO 8 Metabolismo de los carbohidratos
2. Conversión de la fructosa-1,6-difosfato en fructosa-6-fosfato. La reacción irreversible de la glucólisis catalizada por la PFK-1 se evita por la fructosa-1,6- difosfatasa:
Esta reacción exergónica (∆ G ˚′ = −16.7 kJ/mol) es también irreversible en condiciones celulares. El ATP no se regenera, y también se produce fosfato in- orgánico (Pi). La fructosa-1,6-difosfatasa es una enzima alostérica. Su actividad la estimula el citrato y la inhiben el AMP y la fructosa-2,6-difosfato.
3. Formación de glucosa a partir de glucosa-6-fosfato. La glucosa-6-fosfatasa, que sólo se encuentra en el hígado y el riñón, cataliza la hidrólisis irreversible de la glucosa-6-fosfato para formar glucosa y Pi. A continuación, la glucosa se libera en el torrente sanguíneo. Como se ha señalado, cada una de las reacciones anteriores está empatada con una reacción opuesta irreversible en la glucólisis. Cada conjunto de estas reacciones emparejadas se denomina ciclo de sustrato. Debido a que están re- guladas de forma coordinada (un activador de la enzima que cataliza la reacción directa sirve como inhibidor de la enzima que cataliza la reacción inversa), se desperdicia muy poca energía a pesar de que ambas enzimas pueden estar fun- cionando en cierto nivel al mismo tiempo. El control de fl ujos (la regulación del flujo de sustrato y la eliminación del producto) es más eficaz si la acumulación transitoria de un producto se encauza de vuelta al ciclo. La velocidad catalítica de la enzima en sentido directo permanecerá elevada si la concentración del sustrato se maximiza. La ganancia de eficacia catalítica compensa con creces la pequeña pérdida de energía del reciclado del producto. La gluconeogénesis es un proceso que consume energía. En lugar de generar ATP (como la glucólisis), la gluconeogénesis requiere la hidrólisis de seis enla- ces fosfato de alta energía.
La hipertermia maligna es una enfermedad hereditaria poco frecuente que se desen- cadena por determinados anestésicos durante las operaciones quirúrgicas. Un aumen- to considerable (y peligroso) de la temperatura corporal (hasta 44°C) se acompaña de rigidez muscular y acidosis. La contracción muscular excesiva inicia por una gran liberación de calcio del retículo sarcoplásmico, un organelo de almacenamiento de calcio de las células musculares. La acidosis es consecuencia de una producción ex- cesiva de ácido láctico. El tratamiento oportuno para reducir la temperatura corporal y contrarrestar la acidosis, es esencial para salvar la vida del paciente. Un factor que probablemente contribuye con esta enfermedad es el ciclo derrochador entre la glucólisis y la gluconeogénesis. Explique por qué es éste un argumento razonable.
PREGUNTA 8.
Tras examinar la vía gluconeogénica, explique cada componente de la ecuación. ( Pista: la hidrólisis de cada nucleótido libera un protón.)
PREGUNTA 8.
Fructosa-1,6- difosfatasa
HO
HO
O
OH
−O P CH 2
O
O−
CH 2 P^ O−
O
O−
Fructosa-1,6-difosfato
P (^) i
Fructosa-6-fosfato
HO
HO
O
OH
−O P CH 2
O
O−
O O O CH 2 OH
Hipertermia maligna
8.2 Gluconeogénesis 259
Los pacientes con la enfermedad de von Gierke (una enfermedad de almacenamiento de glucógeno) carecen de actividad glucosa-6-fosfatasa. Dos síntomas notables de esta enfermedad son la hipoglucemia en ayunas y la acidosis láctica. Explique por qué se producen estos síntomas.
PREGUNTA 8.
Sustratos de la gluconeogénesis
Como se mencionó antes, numerosos metabolitos son precursores gluconeogénicos. Se describen de forma breve tres de los sustratos más importantes. El lactato lo liberan los eritrocitos y otras células que carecen de mitocondrias o que tienen concentraciones bajas de oxígeno. En el ciclo de Cori , las células mus- culares liberan lactato durante el ejercicio (fig. 8.10). Después de transferir el lactato al hígado, se reconvierte en piruvato a través de la lactato deshidrogenasa y luego en glucosa por gluconeogénesis. El glicerol, un producto del metabolismo de las grasas en el tejido adiposo, se transporta al hígado en la sangre y luego se convierte en glicerol-3-fosfato por medio de la glicerol cinasa. La oxidación del glicerol-3-fosfato para formar DHAP ocurre cuando la concentración citoplásmica de NAD+^ es relativamente elevada.
FIGURA 8.
Ciclo de Cori
Durante el ejercicio extenuante se pro-
duce lactato en las células musculares
en condiciones anaerobias. Tras pasar a
través de la sangre al hígado, el lactato
se convierte en glucosa mediante glu-
coneogénesis.
Torrente sanguíneo
Glucosa
Glucosa
Piruvato
Lactato
Glucosa-6- fosfato
Piruvato
Lactato
Lactato
Glucosa
H
HPO 4
2C 3 H 4 O 3 + 4 + 2 + 2 + 2 +
1C 6 H 12 O 6 + (^4) + 2 + 2 + 6 + (^6) H+
Glucosa
Ácido pirúvico
6 H 2 O
ADP
ATP
GDP
GTP
NAD
NADH
C
CH 2
O
O O−
CH 2 OH
P
O
O−
Glicerol DHAP
C
CH 2 OH
OH H
CH 2 OH
Glicerol-3-fosfato
OH C H
CH 2 OH
CH 2 O P O−
O
O−
Glicerol cinasa
ATP ADP + H+
Glicerol fosfato deshidrogenasa
NAD
+ NADH
Enfermedad
de von Gierke
8.2 Gluconeogénesis 261
concentraciones elevadas de lactato, de glicerol y de aminoácidos. Una alimentación
abundante en grasas, la inanición y un ayuno prolongado proporcionan grandes can-
tidades de estas moléculas.
Las cuatro enzimas clave de la gluconeogénesis (la piruvato carboxilasa, la PEP
carboxicinasa, la fructosa-1,6-difosfatasa y la glucosa-6-fosfatasa) son afectadas en
grados variables por los moduladores alostéricos. Por ejemplo, a la fructosa-1,6-
difosfatasa la activa el ATP y la inhiben el AMP y la fructosa-2,6-difosfato. La acetil-
CoA activa la piruvato carboxilasa. (La concentración de acetil-CoA, un producto de
la degradación de los ácidos grasos, es especialmente elevada durante la inanición.)
La figura 8.12 presenta las generalidades de la regulación alostérica de la glucólisis
y la gluconeogénesis.
FIGURA 8.
Regulación alostérica de la glucólisis
y de la gluconeogénesis
Las enzimas clave en la glucólisis y en
la gluconeogénesis son reguladas por
efectores alostéricos. Activador, +;
inhibidor, −.
Hexocinasa
Glucosa
Glucosa-6-fosfato
Glucosa-6- fosfatasa
Fructosa-6-fosfato
Fructosa-1,6- difosfatasa (+) Citrato (−) Fructosa-2,6- difosfato
(−) AMP
Fosfofructocinasa (+) Fructosa-2,6- difosfato (+) AMP
(−) ATP
(−) Citrato
Fructosa-1,6-difosfato
Fosfoenolpiruvato
Fosfoenolpiruvato Carboxicinasa
Piruvato
Oxaloacetato
(−) Glucosa-6- fosfato
Piruvato carboxilasa
(+) Acetil-CoA
Piruvato cinasa (−) ATP (+) Fructosa-1,6- difosfato
(−) Acetil-CoA (−) Fosforilación dependiente de cAMP
262 CAPÍTULO 8 Metabolismo de los carbohidratos
Como en otras vías bioquímicas, las hormonas afectan la gluconeogénesis al modificar las concentraciones de efectores alostéricos y enzimas clave determinan- tes de la velocidad. Como se mencionó antes, el glucagon deprime la síntesis de fructosa-2,6-difosfato, lo que libera la inhibición de la fructosa-1,6-difosfatasa y desactiva la enzima glucolítica piruvato cinasa a través de una reacción de fosfori- lación iniciada por el cAMP. Las hormonas también influyen en la gluconeogénesis al alterar la síntesis enzimática. Por ejemplo, la síntesis de enzimas gluconeogénicas es estimulada por el cortisol, una hormona esteroidea producida en la corteza de las glándulas suprarrenales que facilita la adaptación del organismo a condiciones estre- santes. Por último, la acción de la insulina provoca la síntesis de nuevas moléculas de glucocinasa, de PFK-1 (inducida por la SREBP1c) y de PFK-2 (favorecida por la glucólisis). La insulina también deprime la síntesis (una vez más a través de la SREBP1c) de PEP carboxicinasa, fructosa-1,6-difosfatasa y glucosa-6-fosfatasa. La acción del glucagon conduce a la síntesis de más moléculas de PEP carboxicinasa, de fructosa-1,6-difosfatasa y de glucosa-6-fosfatasa. Estas hormonas realizan dicha función alterando el estado de fosforilación de determinadas proteínas diana de las células hepáticas, que a su vez modifican la ex- presión de los genes. El punto clave a recordar es que la insulina y el glucagon tienen efectos opuestos en el metabolismo de los carbohidratos. La dirección del flujo de metabolitos (o sea, si se activa la glucólisis o la gluconeogénesis) depende sobre todo de la relación insulina/glucagon. Tras una comida con carbohidratos, dicho cociente se eleva y predomina en el hígado la glucólisis sobre la gluconeogénesis. Tras un periodo de ayuno o luego de una comida con pocos carbohidratos y muchas grasas, el cociente insulina/glucagon es bajo y predomina en el hígado la gluconeogénesis sobre la glucólisis. El segundo regulador importante del control recíproco de la glu- cólisis y de la gluconeogénesis es la disponibilidad de ATP, ya que las cantidades elevadas de AMP, el producto de baja energía de la hidrólisis del ATP, incrementan el fl ujo a través de la glucólisis a expensas de la gluconeogénesis, y las cantidades bajas de AMP incrementan el flujo a través de la gluconeogénesis a expensas de la glucólisis. Aunque el control del ciclo PFK-1/fructosa-1,6-difosfatasa podría parecer sufi ciente para esta vía, el control en la etapa de la piruvato cinasa es clave debido a que permite la retención máxima de PEP, una molécula con un potencial de transfe- rencia de fosfato muy elevado.
8.3 VÍA DE LAS PENTOSAS FOSFATO
La ruta de las pentosas fosfato es otra vía metabólica de oxidación de la glucosa en la que no se genera ATP. Sus productos principales son el NADPH (fosfato de di- nucleótido de nicotinamida y adenina reducido), un agente reductor que se requiere en varios procesos anabólicos, y la ribosa-5-fosfato, un componente estructural de los nucleótidos y de los ácidos nucleicos. La vía de las pentosas fosfato se produce en el citoplasma en dos fases: la oxidativa y la no oxidativa. En la fase oxidativa de la vía, la conversión de la glucosa-6-fosfato en ribulosa-5-fosfato va acompañada de la producción de dos moléculas de NADPH. En la fase no oxidativa se producen la isomerización y la condensación de varias moléculas de azúcar diferentes. Tres intermediarios de este proceso que son útiles en otras vías son la ribosa-5-fosfato, la fructosa-6-fosfato y el gliceraldehído-3-fosfato. La fase oxidativa de la vía de las pentosas fosfato consta de tres reacciones (fig. 8.13a). En la primera reacción, la glucosa-6-fosfato deshidrogenasa (G-6-PD) catali- za la oxidación de la glucosa-6-fosfato. La 6-fosfogluconolactona y el NADPH son los productos de esta reacción. A continuación la 6-fosfo-d-glucono- � -lactona se hidroliza para producir 6-fosfo-d-gluconato. Durante la descarboxilación oxidativa del 6-fosfogluconato, una reacción que produce ribulosa-5-fosfato, se produce una segunda molécula de NADPH. Estas reacciones proporcionan una cantidad sustancial del NADPH que se requie- re para los procesos reductores (p. ej., la biosíntesis de lípidos) y los mecanismos antioxidantes. Por esta razón, esta vía es más activa en las células en las que se
CONCEPTOS CLAVE
- La gluconeogénesis, la síntesis de
moléculas nuevas de glucosa a partir
de precursores que no son carbohi-
dratos, ocurre principalmente en el
hígado.
- La secuencia de reacciones es la
inversa de la glucólisis, excepto por
tres reacciones que evitan los pasos
irreversibles de la glucólisis.
264 CAPÍTULO 8 Metabolismo de los carbohidratos
sintetizan cantidades relativamente grandes de lípidos, por ejemplo, en las del tejido adiposo, de la corteza de las glándulas suprarrenales, de las glándulas mamarias y del hígado. El NADPH también es un antioxidante potente. (Los antioxidantes son sustancias que impiden la oxidación de otras moléculas. En el capítulo 10 se descri- ben sus acciones en los procesos vitales.) Por consiguiente, la fase oxidativa de la vía de las pentosas fosfato también es bastante activa en las células con riesgo alto de experimentar daño oxidativo, como los eritrocitos. La fase no oxidativa comienza con la conversión de la ribulosa-5-fosfato en ribo- sa-5-fosfato, por medio de la ribulosa-5-fosfato isomerasa, o en xilulosa-5-fosfato, a través de la ribulosa-5-fosfato epimerasa. Durante las reacciones restantes de la vía (fi g. 8.13b), la transcetolasa y la transaldolasa catalizan las interconversiones de trio-
FIGURA 8.13b
Vía de las pentosas fosfato
(b) Fase no oxidativa. Cuando las
células requieren más NADPH
que pentosas fosfato, las enzimas
de la fase no oxidativa convierten
la ribosa-5-fosfato en los inter-
mediarios glucolíticos fructosa-6-
fosfato y gliceraldehído-3-fosfato.
H C OH
H C OH
HO C H
CH (^2)
H C OH
H C OH
HO C H
H C OH
H C OH
CH 2
H C OH
H C OH
H C OH
CH 2
C
O
H C OH
C H
O
H C OH
H C OH
HO C H
H C OH
H C OH
H
CH (^2)
H C OH
D -Ribulosa-5-fosfato
D-Ribosa-5-fosfato
D-Gliceraldehído-3-fosfato
D-Fructosa-6-fosfato D-Eritrosa-4-fosfato
(b) D -Gliceraldehído-3-fosfato
D -Sedoheptulosa-7-fosfato
D-Xilulosa-5-fosfato
Ribosafosfato isomerasa
Ribulosa-5-fosfato 3-epimerasa
Transcetolasa
Transaldolasa
Transcetolasa
H
C
O
CH (^2)
C
O
H
C
CH 2 OH
O
CH (^2)
C
CH 2 OH
O
CH (^2)
CH 2 OH
C O
CH (^2)
C
CH 2 OH
O
PO 32 −
PO 32 −
PO 32 −
PO 32 −
PO 32 −
PO 32 −
PO 32 −
PO 32 −
8.3 Vía de las pentosas fosfato 265
sas, pentosas y hexosas. La transcetolasa es una enzima que requiere TPP (pirofos-
fato de tiamina) que transfiere unidades de dos carbonos de una cetosa a una aldosa.
(El TPP es la forma coenzimática de la tiamina, conocida también como vitamina
B 1 .) La transcetolasa cataliza dos reacciones. En la primera, la enzima transfiere una
unidad de dos carbonos de la xilulosa-5-fosfato a la ribosa-5-fosfato, produciendo
gliceraldehído-3-fosfato y sedoheptulosa-7-fosfato. En la segunda, una unidad de dos
carbonos de otra molécula de xilulosa-5-fosfato se transfiere a la eritrosa-4-fosfato
para formar una segunda molécula de gliceraldehído-3-fosfato y fructosa-6-fosfato.
La transaldolasa transfiere unidades de tres carbonos desde una cetosa a una aldosa.
En la reacción catalizada por la transaldolasa, se transfiere una unidad de tres carbo-
nos desde la sedoheptulosa-7-fosfato al gliceraldehído-3-fosfato. Los productos que
se forman son fructosa-6-fosfato y eritrosa-4-fosfato. El resultado de la fase no oxi-
dativa de la vía es la síntesis de ribosa-5-fosfato y de los intermediarios glucolíticos
gliceraldehído-3-fosfato y fructosa-6-fosfato.
Cuando no se requieren azúcares pentosas para las reacciones de biosíntesis, los
metabolitos de la porción no oxidativa de la vía se convierten en intermediarios glu-
colíticos que pueden degradarse posteriormente para generar energía o convertirse en
moléculas precursoras para procesos de biosíntesis (fig. 8.14). Por esta razón, la vía
FIGURA 8.
Metabolismo de los carbohidratos: glucólisis y vía de las pentosas fosfato
Si la célula requiere más moléculas de NADPH que de ribosa, puede canalizar los productos
de la fase no oxidativa de la vía de las pentosas fosfato hacia la glucólisis. Como explica
esta visión general de las dos vías, el exceso de ribosa-5-fosfato puede convertirse en los
intermediarios glucolíticos fructosa-6-fosfato y gliceraldehído-3-fosfato.
Fructosa-1,6-difosfato
Ribulosa-5-fosfato
Fructosa-6-fosfato
Gliceraldehído-3-fosfato
Glucosa-6-fosfato
CO 2
NADH
ATP
ATP
Glucólisis
NADPH NADPH
Ribosa-5-fosfato
Xilulosa-5-fosfato
ATP
Vía de las pentosas fosfato
Piruvato
Glucosa
2 ATP
8.4 Metabolismo de otros azúcares importantes 267
FIGURA 8.
Metabolismo de los carbohidratos: otros azúcares importantes
La fructosa entra en la vía glucolítica por dos caminos. En las células hepáticas, donde se metaboliza la mayoría de
las moléculas de fructosa, la fructocinasa convierte el azúcar en fructosa-1-fosfato, que luego se divide en DHAP
y gliceraldehído. En los músculos y en el tejido adiposo, la fructosa es fosforilada por la hexocinasa para formar el
intermediario glucolítico fructosa-6-fosfato. La galactosa se convierte en galactosa-1-fosfato, que luego reacciona
con UDP-glucosa para formar UDP-galactosa. Esta última es convertida en su epímero, UDP-glucosa, el sustrato
para la síntesis de glucógeno. La manosa es fosforilada por la hexocinasa para formar manosa-6-fosfato, que luego
se isomeriza a fructosa-6-fosfato.
Galactosa-1-fosfato
Galactosa
Glucosa-1-fosfato
Glucógeno
Glucosa-1-fosfato
Glucosa-6-fosfato
Fructosa-6-fosfato
Fructosa-1,6-difosfatasa
Fructosa Manosa-6-fosfato Manosa
Fructosa-1,6- difosfatasa
Fosfomanosa-isomerasa Hexocinasa
UDP-glucosa pirofosforilasa
UDP-galactosa- 4-epimerasa
Gliceraldehído-3-fosfato DHAP
Gliceraldehído Gliceraldehído cinasa
Aldolasa
Fructosa-1-fosfato
Fructocinasa (hígado)
Fructosa
ATP
Galatocinasa
Galactosa-1-fosfato uridililtransferasa
Fructosa-1- fosfato aldolasa
Hexocinasa (^) Fosfoglucomutasa
Glucosa-6-fosfatasa
Hexocinasa (músculo y tejido adiposo) PFK-
Piruvato
ADP
ATP
Glucosa
PP (^) i
UDP- galactosa
UDP-glucosa
ATP
ATP
ATP
ADP ATP
ADP
ADP
ADP
UDP-glucosa
UTP
268 CAPÍTULO 8 Metabolismo de los carbohidratos
La conversión de la fructosa-1-fosfato en intermediarios glucolíticos evita dos pasos reguladores (las reacciones catalizadas por la hexocinasa y por la PFK-1); de esta forma, en comparación con lo que ocurre con la glucosa, la entrada de la fructosa en la vía glucolítica es esencialmente no regulada. En los músculos y en el tejido adiposo, la fructosa se convierte en el intermedia- rio glucolítico fructosa-6-fosfato por conducto de la hexocinasa. Debido a que las hexocinasas tienen baja afinidad por la fructosa, esta reacción tiene una importancia menor a no ser que el consumo de fructosa sea excepcionalmente elevado.
8.5 METABOLISMO DEL GLUCÓGENO
El glucógeno almacena la glucosa. La síntesis y la degradación del glucógeno se regulan con precaución para que pueda disponerse de sufi ciente glucosa para las necesidades energéticas del organismo. La glucogénesis y la glucogenólisis están controladas principalmente por tres hormonas: insulina, glucagon y epinefrina.
Glucogénesis
La síntesis de glucógeno ocurre después de una comida, cuando la concentración san- guínea de glucosa se eleva. Se sabe desde hace mucho tiempo que después de ingerir una comida con carbohidratos ocurre la glucogénesis hepática. La síntesis de glucó- geno a partir de glucosa-6-fosfato implica la siguiente serie de reacciones.
1. Síntesis de glucosa-1-fosfato. La glucosa-6-fosfato se convierte de forma re- versible en glucosa-1-fosfato a través de la fosfoglucomutasa, una enzima que contiene un grupo fosfato unido a un residuo de serina reactivo:
HO
HO
O
CH 2 CH 2
Fructosa-1-fosfato
HO
Fructosa-1- fosfato aldolasa
CH 2
C O
O−
O
CH 2 HO
O P
O−
C
C H
HO
CH 2 OH
H
O
O−
O
O P
O−
C
C H
HO
CH 2
H
O Triosa fosfato isomerasa
DHAP
Gliceraldehído
Gliceraldehído cinasa
Gliceraldehído-3-fosfato
P O−
O−
O
O
OH
ADP
ATP
Hexocinasa
Fructosa Fructosa-6-fosfato
OH
O
HO
HOCH 2 CH 2 OH P
−O
O−
O
CH 2 OH
OH
O
HO
O CH 2
OH OH
ATP ADP
270 CAPÍTULO 8 Metabolismo de los carbohidratos
FIGURA 8.
Síntesis de glucógeno
(a) La enzima glucógeno sintasa rompe el enlace éster del UDP-glucosa y forma un enlace glucosídico � (1,4) entre la glucosa
y la cadena creciente de glucógeno. (b) La enzima ramificante es la causal de la síntesis de enlaces � (1,6) en el glucógeno.
O−
OH
HO
OH
O
HOCH 2
O
P O P O
O−
O O
Uridina
HOCH 2
OH
HO
OH
O
O−
P O P O
O−
O O
+^ −O
HOCH 2 HOCH 2
HO
OH
O
HO
OH
O
HOCH 2 HOCH^2
HO
OH
O
HO
OH
O
OH
UDP-glucosa Cebador de glucógeno ( n residuos)
Glucógeno ( n + 1 residuos) UDP
Uridina
Glucógeno sintasa
O O O
O O
(a)
HO
HO
OH
O
O
CH 2 OH CH 2 OH
HO
OH
O
HO
OH
O
O
HO
OH
O
CH 2 OH CH^2
Se forma el enlace glucosídico α(1,6)
CH 2 OH
HO
O
OH O
HO
HO
OH O
CH (^2) OH
CH (^2) OH
O
O
OH
Enzima ramificante
CH 2 OH
HO HO
OH
O
HO
OH
CH 2 OH CH 2 OH
HO
OH
O
O
CH 2 OH CH 2 OH
HO
OH
HO
OH
HO
OH
CH 2 OH CH 2 OH
OH
O
O
O O
O
O
O O O O O O
O O O
O O
(b)
8.5 Metabolismo del glucógeno 271
gránulos grandes de glucógeno, cada uno formado por una sola molécula de glu- cógeno muy ramificada. Las enzimas causales de la síntesis y de la degradación del glucógeno recubren cada gránulo.
Glucogenólisis
La degradación del glucógeno requiere las dos reacciones siguientes:
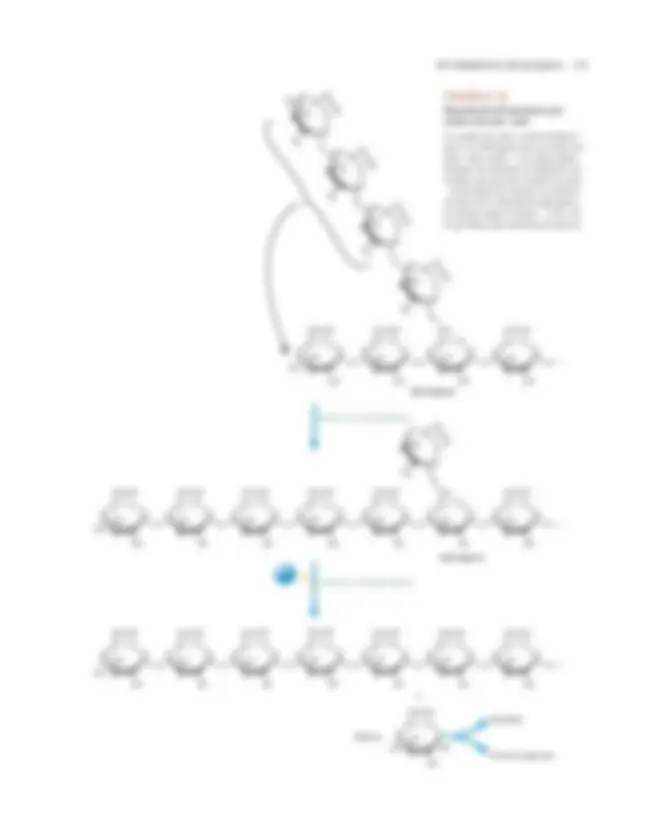
1. Eliminación de la glucosa de los extremos no reductores del glucógeno. La glucógeno fosforilasa utiliza fosfato inorgánico (Pi) para romper los enla- ces � (1,4) de las ramificaciones externas del glucógeno para formar glucosa- 1-fosfato. La glucógeno fosforilasa se detiene cuando llega a cuatro residuos de glucosa del punto de ramificación (fig. 8.17). (Una molécula de glucógeno que se ha degradado hasta estos puntos de ramificación se denomina dextrina límite .) 2. Hidrólisis de los enlaces glucosídicos � (1,6) en los puntos de ramificación del glucógeno. La amilo- � (1,6)-glucosidasa, que también se denomina enzima desramificante, comienza a eliminar los puntos de ramificación � (1,6) al trans- ferir los tres residuos de glucosa más externos de los cuatro unidos al punto de
FIGURA 8.
Degradación del glucógeno
La glucógeno fosforilasa cataliza la separación de los residuos de glucosa de los extremos no reductores de una cadena de glucóge-
no para formar glucosa-1-fosfato. En esta ilustración se retira un residuo glucosa de cada extremo no reductor. El desprendimiento
de residuos glucosa continúa hasta que en cada punto de ramificación quedan cuatro residuos.
HO
OH
O
O
CH 2 OH CH 2 OH
HO
OH
O
HO
OH
O
HO
OH
O
CH 2 OH CH 2
HO
OH
O
CH
2 OH
HO
OH
HO O
OH
CH
2 OH
CH
2 OH
OH
Glucógeno fosforilasa
HO
OH
O
O
CH 2 OH
HO PO^3
2 −
HO
OH
O
O
CH 2 OH
HO PO^3
2 −
Glucosa-1-fosfato
Glucógeno
HPO 42
−
HO
HPO 42
−
O O O
O
CH
2 OH
HO
O
CH
2 OH
HO
HO
O
HO
OH
O
O
CH 2 OH
HO
OH
O
HO
OH
O
CH 2 OH CH 2
HO
OH
O
O
CH
2 OH
HO
OH
O
O
CH
2 OH
Glucógeno
O O
HO O
OH
O
CH
2 OH
HO
OH
O
CH
OH^2 OH
OH
O
O
O
O
O
O
8.5 Metabolismo del glucógeno 273
FIGURA 8.
Degradación del glucógeno por
enzima desramifi cante
Los puntos de ramificación del glucó-
geno son eliminados por la enzima des-
ramifi cante amilo- � (1,6)-glucosidasa.
Después de transferir la unidad de tres
residuos que precede al punto de rami-
fi cación hacia un extremo no reductor
cercano de la molécula de glucógeno,
la enzima rompe el enlace � (1,6), con
lo que libera una molécula de glucosa.
HO
OH
O
CH 2 OH
HO HO
Glucólisis
Torrente sanguíneo
Glucosa
HO
OH
O
O
OH CH 2 OH CH 2 OH
HO
OH
O
HO
OH
O
HO
OH
O
CH 2 OH CH 2 OH
O O O
CH 2 OH
HO
OH
O
HO
OH
O
HO
OH
O
CH 2 OH CH 2
O O O
HO
OH
O
O
CH 2 OH CH 2 OH
HO
OH
O
HO
OH
O
HO
OH
O
CH 2 OH CH 2
O O O
CH 2 OH
HO
OH
O
HO
OH
O
HO
OH
O
CH 2 OH CH OH 2
O O O
HO
OH
O
O
CH
2 OH
O
Glucógeno
Glucógeno
OH
O
O
CH 2 OH CH 2 OH
OH
O
OH
O
OH
O
CH 2 OH CH 2
O O O
CH
2 OH
HO
OH
O
HO
OH
O
CH
2 OH
O
HO O
OHOH
O
CH
2 OH
HO
OH
O
O
CH
2 OH
Amilo-α(1,6)-glucosidasa
HO
HO
HO
OH OH OH OH
H 2 O
Amilo-α(1,6)-glucosidasa
274 CAPÍTULO 8 Metabolismo de los carbohidratos
FIGURA 8.
Degradación del glucógeno: resumen
La fosforilasa de glucógeno rompe
los enlaces � (1,4) del glucógeno para
producir glucosa-1-fosfato hasta que
llega a cuatro residuos de glucosa de
un punto de ramificación. La enzima
desramifi cante transfiere tres de estos
residuos a un extremo no reductor
cercano y libera el cuarto residuo como
glucosa libre. Las acciones repetidas de
ambas enzimas pueden conducir a la
degradación completa del glucógeno.
Enzima desramificante
Extremo reductor
Extremos no reductores
Glucógeno fosforilasa
P (^) i
Glucosa-1-fosfato
Enzima desramificante
Dextrina límite Enzima desramificante
Glucosa-1-fosfato + glucosa
Glucógeno fosforilasa
Glucosa-1-fosfato
Glucógeno fosforilasa
Pi
Glucosa