¡Descarga Mitocondrias: Genoma, Síntesis y Translocación de Proteínas - Prof. Antón y más Apuntes en PDF de Biología Celular solo en Docsity!
TEMA 12:
MITOCONDRIAS
CARACTERISTICAS COMUNES DE MITOCONDRIAS Y CLOROPLASTOS
- Función común: Coger energía y transformarla
- Especialización en conversión de energía
- CLOROPLASTOS: Utilizan la energía del sol para CREAR enlaces—compuestos organicos (Gliceraldehido-3-P)
ENERGIA DE LA LUZ—ENERGIA QUÍMICA DE LOS ENLACES
- (^) MITOCONDRIAS: ROMPEN los enlaces—la energía que surge la utilizan para formar ATP
ENERGIA QUÍMICA DE LOS ENLACES—ATP
- (^) Son semiautónomos: tienen DNA y ribosomas propias—rRNA, tRNA, mRNA(prot. MMI)
- Los dos tienen un origen endosimbiotico
- Los dos tienen une doble membrana y diversos subcompartimientos(entre 4-6)
- (^) Membrana externa(debido a su origen endosinbiotico)
- Espacio intermembranoso
- Membrana interna
- En la cavidad interior 4.a. MITOCONDRIAS: Matriz 4.b.CLOROPLASTOS: Estroma
SOLO LOS CLOROPLASTOS:
- Tilacoides
- Espacio tilacoidal
ESTRUCTURA Y COMPOSICIÓN DE LAS MITOCONDRIAS
BIOGENESI DE LAS MITOCONDRIAS
- Se fabrican a través de las mitocondrias ya existentes—FISIÓN
- Tiene lugar mientras la célula está en interfase—pero no es como en el núcleo no tiene fases (G1, S…) puede ocurrir en cualquier momento de la interfase
- Modelo de partición de la mitocondria:
- Crecimiento, para que la mitocondria crezca: ■ Replicación del DNA mitocondrial ■ Sintesi de proteínas de la membrana mitocondrial interna (MMI) ■ Importación de proteínas del citosol—Las mitocondrias son SEMIAUTONOMAS—tienen proteínas propias pero también proteínas que se sintetizan en el citosol(estas últimas son el 96%del total) ■ Importación de lípidos del RE—mediante LA PROTEINA INTERCAMBIADORA DE FOSFOLÍPIDOS: Cogen fosfolípidos de la membrana externa del RE y las pasan a la membrana externa de las mitocondrias
- División para partición (fusión de crestas). Las mitocondrias se parten en dos para aumentar el número de dichos orgánulos. Para que esta partición ocurra, las crestas deberán de fusionarse *Es un orgánulo bastante versátil—Se puede fusionar y fisionar
GENOMA MITOCONDRIAL Y SINTESIS DE PROTEINAS
CARACTERÍSTICAS:
- El DNA de las mitocondrias es CIRCULAR
- Su medida es variable dependiendo la especie
• EPILEPSIA MIOCLÓNICA CON FIBRAS ROJAS ESTRIPADAS(MERRF)
- Causado por una mutación en un gen que codifica tRNA
- Consecuencias ■ Disminuye la síntesis proteica ■ La CTE (Cadena de transporte de electrones) se ve afectada ■ La producción de ATP disminuye
- Afecta a los tejidos musculares y nirviosos
- Consecuencias/síntomas: Sebilidad muscular, trastornos cardiacos, epilepsia, demencia…)
• NEUROPATIA ÓPTICA HEREDITARIA DE LEBER
- Causado por una mutación en los genes que codifican D4, ND1, ND6 (Son proteínas, subunidades,que forman parte del COMPLEX I de CTE
- La mayoría de células puede compensar dicha mutación, excepto las células del nervio—produce la muerte celular de nervio óptico
- CONSECUENCIA: Pérdida indolora de la visión central en los 2 ojos con12-30 años
• ENCEFOLOMIOPATIA MITOCONDRIAL
- Causado por una mutación en un gen que codifica tRNA—disminuye la síntesis proteica—disminuye la producción de ATP
- Consecuencias: Disfunción de la fosforilación oxidativa en células de parénquima cerebral (están en la región funcional)
- Consecuencias/síntomas: Retraso en el desarrollo y crecimiento, dificultades de aprendizaje/atención, convulsiones…
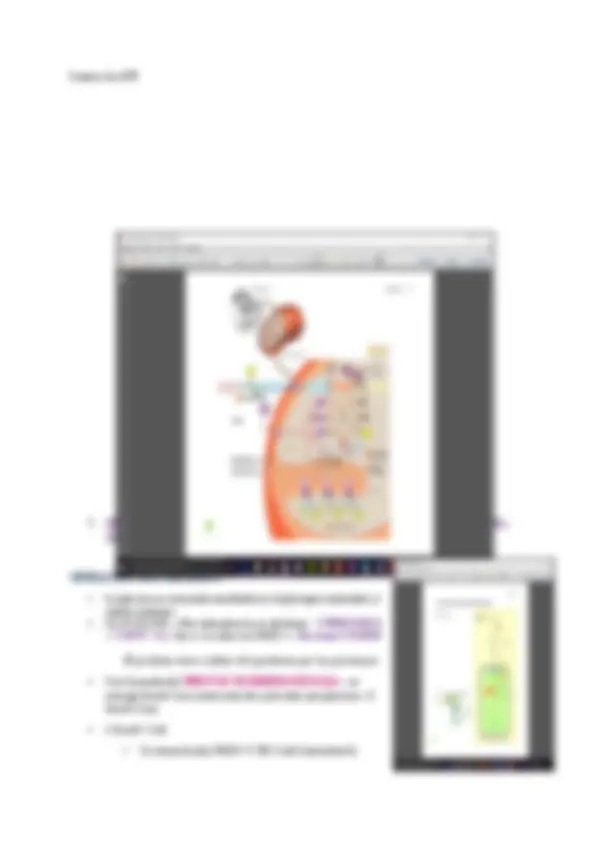
IMPORTACIÓN DE PROTEINAS DEL CITOSOL
La translocación es postraduccional—las proteínas se sintetizan en el citosol y posteriormente son translocadas
- La secuencia señal se encuentra en el extremo N-terminal(se elimina en la matriz)
- Se llama secuencia de DIRECCIONAMIENTO a la matriz—esta señal hace que todas las proteínas vayan a la matriz y luego se redistribuyen
- La señal no es una secuencia de aá concreta
- Su característica principal: Cuando la proteína se pliega—forma una hélice— las cargas se ordenan de manera que esta hélice sea anfipática(todas las proteínas con carga positiva a un lado/ todas las proteínas hidrofóbicas al otro lado)
- Cada proteína aparte de esta secuencia de direccionamiento consta de más secuencias para que la proteína sea redistribuida a su zona correspondiente
TRANSLOCACIÓN DE PROTEINAS DEDE EL CITOSOL
- Tiene lugar en los puntos de contacto—los puntos donde las dos membranas se unen
Se hace de forma post-traduccional—requiere gasto de energía (no se utiliza la energía
de la síntesis)
- Los complejos proteicos necesarios:
PROYECCIÓ
N
(4-6 aá por vuelta)
se asocia a las chaperonas—TIM 44
¿POR QUE TIENE RECEPTORES EN LA MEMBRANA Y NO RECEPTORES SOLUBLES
COMO EN EL CASO DEL RE?
En una proteina que se crea en el RE tiene que entrar rapidamente al RE--es imprescindible que haya receptores solubles—no pueden esperar que por casualidad se una al transloco (en mitocondrias no es tan necesario) Proteina plegada en el citosol
PROTEINAS DE LA MEMBRANA MITOCONDRIAL INTERNA
Dependiendo del tipo de proteina siguen caminos diferetes (A,B,C)
CAMINO A (PROTEINA UNIPAS CON SEÑAL DE DIRECCIONAMIENTO)
- El proceso es igual que en las proteinas de la matriz mitocondrial
- (^) Siguen las proteinas transmembranales de un solo paso (unipas)
- Estas proteinas ademas de la secuencia de direccionamiento en el extremo N-terminal tienen otra secuencia—ST (Stop transfering secuence)
- (^) La proeina pasa el primer canal y se adentra en la siguiente TIM
- Las chaperonas van estirando la proteina hasta que esncuentran una secuencia hidrofóbica—el TIM se va - Mientras se va estirando la MPP corta la secuencia de direccionamiento
- (^) La proteina se queda anclada
CAMINO B( PROTEINA MULTIPAS CON SEÑAL DE DIRECCIONAMIENTO)
- (^) El proceso es igual excepto:
- Entra la proteina entera a la matriz
- La MPP corta la secuencia de direccionamiento
- Las chaperonas (HSP 70)reconocen una secuencia interna y se unen pero no la pliegan —la mantiene estirada
- OXA1, mediante un mecanismo desconocido reconoce las secuencias hidrofóbicas y colocan la proteina en la membrana
- Siguen la mayoria de las proteinas multipass
CAMINO C (ALGUNAS PROTEINAS MULTIPAS, SIN SECUENCIA DE
DIRECCIONAMIENTO)
- Muy pocas proteinas siguen este camino
- Estas proteinas no tienen la secuencia de direccionamiento (no helice anfipática)
- Se desconoce que reconoce el receptor
- Despues de pasar el primer canal (TOM 22 /70 ) una TIM 9/10 (un tipo de chaperonas) asociada a la membrana interna—ayuda a que la proteina se inserte en el segundo canal (TIM 22/54 )—supone un gasto de energia
- La TIM (mediante un metodo desconocido), reconoce algo y inserta la proteina dentro de la membrana (no le deis más bueltas, se desconoce todo lo demás, solo intentar entender más o menos el proceso) *Son proteinas multipas sin secuencia
PROTEINAS DEL ESPACIO INTERMEMBRANOS
CAMINO A
- La mayoria de las proteinas del espacio intermembranoso sigen el camino A
- El proceso es el mismo que hemos visto antes hasta que la proteina llega al punto 2
- Aparece una secuencia hidrofobica—se engancha
- (^) La proteasa reconoce una diana—la corta
- La proteina final queda liberada en el espacio intermembranoso
*LA proteina lleva una secuencia más a parte de la secuencia que la lleva hasta la matriz
CAMINO B
- Lo siguen unas cuantas proteinas que no tienen una secuencia anfipatica (como las demás)en el extremo N-terminal
- No se sabe que secuencia se utiliza ni quien dirige la entrada total
PROTEINAS DE LA MEMBRANA MITOCONDRIAL EXTERNA
Siguen dos vias distinas -Las proteinas que tienen los dominios hidrofobicos en helice α
-Las proteinas que tienen los dominios hidrofobicos en lamina β
-Tienen una secuencia que satura construida por aá hidrofóbicos
-La proteína entra completamente -Se necesitan chaperonas— gasto de E—
Sintesis de ATP
1. OXIDACIÓN DEL PIRUBATO, ACIDOS GRASOS HASTA CONSEGUIR ACETIL-
CoA
1.a.
OXIDACIÓN DEL PIRUBATO
- La glucosa se encuentra reserbada en el glucogen (animales) y midón (plantas)
- GLUCOLISIS—Por cada glucosa se obtienen: 2 PIRUVATO + 2 ATP + 4 e -^ (los e-^ se unen con NAD +-- Se crean 2 NADH
El pirubato entra a fabor del gradiente por las permeasas
- Con la ayuda del PIRUVAT DESHIDROGENASA— se consige Acetil-Coa (como eran dos piruvatos por glucosa— Acetil-Coa)
- 1 Acetil- CoA
- Se necesita más NAD+ Y HS-CoA(Coenzima A)
1.b. OXIDACION DE LOS ACIDOS GRASOS
- Más eficiente que el piruvato
- Se utiliza Acil-Coa (conseguimos por la comida o por las reservas de los trigliceridos—se cogen los ACIDOS GRASOS
- Dos fases
Con gasto de ATP se consiguen Acilgrasos-Coa (n=C)
*Gracias a las permeasas entran dentro de la matriz
1.c. OXIDACIÓN DE AMINOACIDOS
- Normalmetne los aá son reciclados para crear nuevas proteinas
- Pero se pueden utilizar
- (^) El proceso es espécifico para cada aá
- Solo se utilizan en casos extremos
- Condiciones de agotamiento de carbohidratos y acidos grasos(por inanición)
- Consumo excesivo de proteinas en la dieta
OXIDACIÓN EN EL CICLO
DE KREBS
PARTICIPAN LAS
SIGUIENTES ENZIMAS:
-Enzimas solubles de la matriz
Los dos C añadidos se eliminan en forma de CO 2
*La energia qu ese desprende en la CTE impulsara los H+--ayudan a la ATP SINTASA
El aceptor de electornes:
- NADH: COMPLEJO Nº
- FADH 2 : COMPLEJO Nº *Los dos complejos son grandes complejos proteicos—les cuesta mover—por eso en estas cadenas hay elementos moviles como UBIQUINONA
Cuando más abancemos en la cadena—los electrones tienen menos energia + la afinidad de los
complejos hacia los electrones es mayor—condicionan el recorrido de los electrones—mantienen una direción
*Los complejos formados por muchas proteinas tienen diferentes grupos
TABLA RESUMEN
PROETINAS Y COMPLEJOS QUE PARTICIPAN
• CITOCROMO
- Captan 1 e -
- Hay de 5 tipos en CTE: a, a 3 ,b 566 …)
- En CTE más de 100
- Pueden estar libres o ancladas a otros complejos
- FUNCIONAMIETO: 4 atomos de Nitrogeno anclan Fe— se oxida o se reduce
PROTEINAS FERROSULFURADAS
- Captan 1 e -
- La hay de 7 tipos en la CTE
- Tienen atomos de Fe y S que se oxidan y se reducen
UBIQUINONA O COENZIMA Q
- (^) Transporta H(como maximo 2)
- Se mueve
- Es hidrofóbica
• FLAVOPROTEINAS
- Transportan H+ (igual que la ubiquitina captan e- en forma de H
- FAD: Flavina Adenina Dinucleotido
- (^) FMN: Flavina mononucleotido
- ATOMOS DE COURE (Captan 1 e - )
- Captan 1 e -
- Se oxidan o se reducen
MODELO DE TRANSPORTE DE PROTONES QUE REALIZAN LOS COMPLEJOS PROTEICOS
DE LA CTE
Los cambios alostericos que sufren—afectan a las subunidades por separadoo
- A medida que los electrones pasan, los complejos sufren cambios alostericos—solo
Cola hidrofóbica— por eso se mantienen en la membrana
EJEM: La ubiquinona no interacciona con el COMPLEJO IV
H+ sale—no se acidifica—las porinas hacen que el espacion intermembranoso este en contacto con el citoosol
SINTESIS DE ATP
ATPsintasa:
- Bomba de tipo F—son como los de tipo P pero con la diferencia de que estan orientados en sentidos opuestos
- Misma estructura que las bombas de tipo P
■ Tres subunidades—ABC
■ Subunidades α y β—captan ADP y P—crean ATP
■ Subunidades γ, δ y ε—giran el eje de rotación
- El 15% de las proteinas de la MMI
- Se utiliza la energia de gradiente para el sintesis—por 3 H+--se obtiene energia mecanica—la subunidad 0 2 6 3rota—provoca cambios de conformación en la subunidad β(subunidad que cataliza)—se obtiene 1 ATP
- Por 1 vuelta de 360º--se obtienen 3 ATPs—una por cada unidad de subunidad β
- Se crean 100ATPs por segundo
PROCESO
- Entran H+ -- ENERGIA QUIMICA
- Gira el eje -- ENERGIA MECÁNICA
H+ sale—no se acidifica— las porinas hacen que el espacion intermembrano
GRADIENTE
ELECTROQ
UÍMICO —
DIFERENCIA
DE
POTENCIAL
Basidifica la matriz
- Cambio de comformación de la subunidad β —se sintetiza ATP – ENERGÍA QUIMIBA (Se libera energia para crear enlaces)
*In-vivo, el proceso no pasa en el sentido opuesto *A la ATPsintasa le unieron actina a la ATPsintasa para ver como funcionava
Como minimo
EJEMPLOS DE RENDIMIENTO MÁXIMO
Teniendo en cuenta que el gasto es mínimo
De la ATP sintasa
Por cada P (fosforo) que entra en la matriz, necesitamos sacar un H+
ANTIPORT TA2 (Utiliza el gradiente electrico)
- La carga propia del OH---hace que esta salga fuera de la matriz por consecuencia de la diferencia de potencial de la MM
- Instantaneamente a la salida de OH-, este se junta con H+ creando una molécula de H20— GASTAMOS UN H+
*Con cualquiera de los dos métodos gastamos 1H
PRODUCCIÓN DE CALOR
- TERMOGENINA (UCP1= Proteina Desacopladora 1)— transporta H+ a favor del gradiente—transforma la energia del paso en energia termica
- Solo lo tienen unas mitocondrias concretas de unos tejidos concretos
- En el greix bru (la grasa parda)—hay más mitocondrias con termogenina (15%de las proteinas de MMI)
En los tejidos de los siguientes mamíferos-
■ Neonatos ■ (^) Animales que hivernan ■ Animales de las regiones árticas
Tiene un aspecto más oscuro(más ATPsintasas con termogenina
Es necesario un aporte extra de calor para mantener la Tª corporal de los niños Cuando nacemos, tenemos grasa parda recubriendo toda la espina dorsal y la parte de apajo Porque no pueden mantener la temperatura:
- Porque en los adultos las venas se contraen, pero en los bebes no
- (^) Nosotros nos podemos tapar
- Nos podemos mover para entrar en calor
- Tenemos escalofríos—los bebes no (el sistema neuronal no esta tan desarrollado)
- Su superficie es mayor que su masa (la superficie es por donde se escapa el calor)
Se encuentra más en: