¡Descarga procesos biológicos 2 y más Resúmenes en PDF de Biología solo en Docsity!
Resumen de PB
SEMANA 1
-resumen de lecturas Receptores acoplados a proteína G: estructura y mecanismo La clase más numerosa de receptores son los receptores acoplados a proteína G (GPCR). En los seres humanos, los GPCR se utilizan para detectar y responder a diferentes tipos de señales (neurotransmisores, hormonas). Todas las vías de transducción de la señal de los GPCR tienen los siguientes elementos en común: (1)un receptor que contiene 7 hélices x que atraviesan la membrana, (2)una proteína G trimérica (también se le puede llamar heterotrimérica porque tiene 3 tipos de subunidades distintas: x,b,y), (3)una proteína efectora unida a la membrana y (4)proteínas que participan en la regulación por retroalimentación y desensibilización de la vía de señalización. En muchas vías GPCR, también se produce un segundo mensajero. Las vías GPCR suelen tener efectos a corto plazo en la célula al modificar rápidamente proteínas existentes, ya sea enzimas o canales de iones. Estas vías permiten a las células responder con rapidez a una variedad de señales, ya sea estímulos ambientales (luz) u hormonales (adrenalina). Todos los receptores acoplados a proteína G tienen la misma orientación en la membrana y contienen 7 regiones x helicoidales transmembrana, 4 segmentos extracelulares y 4 segmentos citosólicos. El extremo N-terminal está sobre el lado exoplasmático y el extremo C-terminal está sobre el lado citosólico. El segmento carboxilo terminal C4, el lazo C3, y en algunos receptores también el lazo C2 están involucrado con las interacciones de la proteína G trimérica. Los GPCR están anclados en el centro hidrófobo de la membrana plasmática por aa hidrófobos sobre las superficies más externas de los 7 segmentos que atraviesan la membrana. Un grupo de GPCR cuya estructura molecular es conocida en detalle son los receptores B- adrenérgicos , que unen hormonas como la adrenalina y noradrenalina. En estos y otros receptores, los segmentos de hélices x embebidas en la membrana y lazos extracelulares forman el sitio de unión al ligando que se abre hacia la superficie exoplasmática. Mientras que todos los GPCR comparten la misma estructura básica, diferentes subtipos de GPCR pueden unir la misma hormona con diferentes efectos celulares. La hormona adrenalina se activa en momentos de lucha o huida. En los mamíferos, la liberación de glucosa y ácidos grasos es desencadenada por la unión de adrenalina (o sus derivados de noradrenalina) a los receptores B-adrenérgicos sobre la superficie de células hepáticas y adipocitos. Además, unida a los receptores B-adrenérgicos sobre las células musculares cardiacas aumenta la velocidad de contracción; la adrenalina estimula los receptores B-adrenérgicos sobre las células del musculo liso del intestino y provoca la relajación; y al unirse con los receptores B-adrenérgicos del músculo liso de los vasos sanguíneos en el tracto intestinal, la piel y los riñones causa la constricción de los vasos sanguíneos y se corte la circulación a dichos órganos.
Las proteínas G triméricas contienen 3 subunidades: x, b y y. La subunidad (^) B y (^) Y siempre están unidas juntas y suelen mencionarse como la subunidad GBY. En el estado de reposo, la subunidad GX tiene un GDP unido y forma un complejo con GBY. La unión de un ligando a un GPCR cambia la conformación de sus lazos citosólicos y permite al receptor unirse a la subunidad GX. Esta unión libera el GDP unido; así el receptor unido a ligando funciona como un factor de intercambio de nucleótido de guanina (GEF) para la subunidad GX. Luego el GTP se une al sitio de nucleótido de guanina “vacío”, lo que causa un cambio conformacional de sus segmentos interruptores. Estos cambios debilitan la unión de GX con el receptor y la subunidad GBY. En la mayoría de los casos GX GTP que permanece anclado en la membrana, luego interacciona con una proteína efectora y la activa (en algunos GX GTP inhibe al efector). El estado GX GTP activo es de corta duración debido que el GTP es hidrolizado a GDP en minutos (la velocidad de la hidrólisis de GTP es a veces
potenciada aún más mediante la unión del complejo GX GTP al efector, por lo tanto, el efector funciona como una proteína de activación GTPasa), catalizado por la actividad GTPasa intrínseca de la subunidad GX. Por ende, la conformación de la GX vuelve a cambiar al estado inactivo GX GDP y bloquea cualquier activación posterior de proteínas efectoras y se vuelve a unir con el complejo GBY. *GAP: Proteína aceleradora de GTPasa, activa el dominio GTPasa. Se encuentra a nivel citosólico.
En el cuadro se resumen las principales clases de proteína G con diferentes subunidades GX. Los receptores B-adrenérgicos, llamados B1 Y B2 están acoplados a una proteína G estimuladora cuya subunidad alfa activa enzimas efectoras unidas a membrana llamadas adenilcicclasas. Una vez activada, esta enzima cataliza la síntesis del segundo mensajero (^) CAMP. Por el contrario, el subtipo X2 del receptor adrenérgico está acoplado a una proteína G inhibitoria cuya subunidad alfa inhibe a la adenilciclasa (proteína transmembrana con dominio citosólico enzimático).
Los transportadores que se encuentran dentro del grupo de bombas impulsadas por ATP, canales iónicos, transportadores (uniporte, simporte, antiporte) facilitan el movimiento de pequeñas moléculas específicas o iones. La mayoría de nuestras células utilizan glucosa como sustrato para síntesis de energía: glucólisis ● Ejemplo 1 – GLUT1: a nivel de tejido nervioso, eritrocitos y tejido fetal. Transporte Uniporte de glucosa: transportador (GLUT1) sufre un cambio de conformación que permite el paso de glucosa – aminoácidos se reorganizan de forma diferente; implica reordenamiento de cargas y aminoácidos.
● Ejemplo 2 – Enterocito a nivel de intestino: se da la absorción de glucosa de la luz intestinal/parte apical de la célula por el Simporte Na/Glu – SGLT 1: necesita de 2 moléculas de Na (260 moléculas de H ingresan acompañando al Na+) para que 1 glu ingrese, parte de la glucosa se queda para el metabolismo de la célula, mayor parte pasa por Glut2 uniporte (liver, pancreas, intestine); sodio se queda en la célula y sale por acción de la bomba Na/K – ATPasa.
● Ejemplo 3 – Eritrocito: ● Capilares sistémicos (sangre desoxigenada): ingresa CO 2 por difusión simple, actúa enzima anhidrasa carbónica utilizando OH del H formando bicarbonato (sale bicarbonato e ingresa cloro a través de la proteína AE1 o antiportador Cl-/HCO3-), H+^ del H20 se queda en la Hb haciéndola más ácida. Se modifican los residuos de histidina (a que se ve afectado por la presencia de H +), se desacopla el O2.
● Capilares pulmonares (sangre oxigenada): bicarbonato ingresa por mismo antiportador (proteína AE1: cataliza el intercambio reversible de iones Cl-^ y HCO3-), anhidrasa carbónica es capaz de degradar bicarbonato a CO 2 y OH- formando agua a partir del OH- volviendo el medio intracelular más básico. Residuos de histidina no se ven afectados, el hierro se vuelve más afín con el O 2 y lo atrae (entra). El CO 2 sale.
Bombas impulsadas por ATP: se les denomina ATPasas de Transporte
● Bombas asociadas a ATP: ● Ejemplo Bomba Na/K: clase P , ingresa 2K+^ y sale 3Na+: Mg se utiliza como cofactor de ATPasa. ● Ejemplo Bombas en lisosomas/vacuolas/endosomas: clase V , permite el ingreso de ácido que activa las enzimas de los lisosomas que degradan las moléculas. Apertura de canal de Cl hace del medio más ácido.
● Ejemplo Bombas Cl/bicarbonato: (antiportador o proteína AE1) en células parietales (pH 7.2) del estómago permiten la formación del ácido clorhídrico (Cl sale de la célula por el canal de Cl) para acidificación del estómago (pH 1: permite eliminar patógenos ingeridos y desnaturalizar numerosas proteínas). (1)El CO 2 ingresa por difusión simple, la anhidrasa carbónica(2) utiliza el CO 2 , y el OH-^ del H 2 0 para formar bicarbonato y sale de la célula a través del antiportador Cl-/HCO3-. Queda el(4) H + del agua que sale a la luz del estómago (para aumentar el pH de la célula: formación de HCl) por la bomba ATPasa H/K( clase P: estómago de mamíferos ). *Para lograr acidez en el estómago: H+, CO 2 , Cl-
Luz del estomago(1.0)
● Omeprazol: inhibe la bomba ATPasa H +/K+ no se genera el ácido clorhídrico, disminuye el nivel de acidez en el estómago.
● ● Glut asociado a músculo e hígado: Receptor de insulina capta insulina (sintetizado en células beta del páncreas), interaccionan y activa una cascada de señalización que permite que los Glut-4 se transloca a la membrana para que pueda transportar glucosa dentro de la célula (interacción receptor de insulina-insulina + Glut-4 favorecen la formación de glucógeno en el músculo). ● Glut-2 en el hígado junto con receptor insulina-insulina favorecen la síntesis de glucógeno y lipogénesis (formación de nuevos lípidos: relacionado en el transporte de lípidos – síntesis de bilis/ácidos biliares por hidroxilación del colesterol; formación de lipoproteínas y ácidos grasos) en el hígado.
● Transportadores tipo Glut: Glucosa de la digestión se conduce por el torrente sanguíneo a células del cerebro a través de la barrera hematoencefálica, el transportador Glut-1 permite el transporte de glucosas dentro de las células nerviosas. Glucosa utilizada para sinapsis.
● Señalización Celular: ● Proteína efectora: genómica (ingresa al núcleo, se modifica la expresión génica); no genómica (síntesis de proteínas, metabolismo, desarrollo, crecimiento celular - citosol). ● Una proteína sustrato puede activarse mediante fosforilación (no necesariamente debe estar fosforilada para estar activa; puede estar inactiva siendo fosforilada); se puede activar cuando pierde el grupo fosfato a través de la enzima proteína fosfatasa. ● Génica: se da en el núcleo (activador de genes, supervivencia celular) – No génica: en el citosol (metabolismo, desarrollo, crecimiento celular).
TIPOS DE RECEPTORES:
● Receptor tirosinas-quinasa necesita de un dominio tirosinas-quinasa intrínseco (dentro de él); se fosforila a sí mismo (autofosforilación) ● Receptor asociado a proteína G (ejm: x y B-adrenérgicos, glucagón, quimiocinas, rodopsina, muscarinico) ● Receptores-canales iónicos (neurotransmisores, iones, nucleótidos, IP 3 ) ● Receptores con actividad tirosina cinasa intrínseca: Es parte del receptor. (ejm: Factores de crecimiento, insulina) ● Receptores asociados a tirosina cinasa: No parte del receptor. (receptores antigénicos) ● Ligandos agonistas: cumplen una función // Antagonistas: no cumplen una función
Tipos de receptores: Receptor asociado a un canal Receptor con actividad enzimática: (1)Receptor tirosina cinasa intrínseca (2)Receptor JAK-STAT (3)Receptor Serina-treonina quinasa Receptor asociado a proteína G y sistema efector
Efectores asociados con los receptores acoplados a proteína G: ● Fosfolipasa C: función– enzimática; degrada fosfolípidos (fosfatidilinositol-4,5-bifosfato o PIP 2 ) ● Hidroliza y obtiene: DAG (diacilglicerol – se queda en la membrana) + IP 3 ● IP 3 activa canal de Ca y permite su salida al citosol ● DAG interactúa con la PKC (dependiente de Ca) + Ca y activa la vía de la proteína quinasa dependiente de Ca (PKC): la interacción sucede en la membrana ● DAG, PIP 2 , IP 3 son segundos mensajeros de la proteína Gq ● Ca+2 y derivados fosfatidilinositol unidos a membrana son considerados como segundos mensajeros. ● Termolábil: no resiste cambios de temperatura
Propiedades de las enzimas: ● Mayor velocidad de reacción ● Condiciones de reacción moderada: Tº->Si la temperatura es baja, las enzimas tienen baja actividad. Al aumentar la temperatura (>37º) la actividad enzimática se incrementa, pero si es más de 50º la estructura terciaria de las proteínas se destruye. // pH-> las enzimas son + activas cuando se encuentran en el pH óptimo (pH al que se mantiene la estructura terciaria de la proteína). ● Mayor especificidad de reacción: capacidad de actuar de manera selectiva ante un sustrato. ● Capacidad de regulación: la enzima puede ser regulada por inhibidores, etc.
*La anhidrasa carbónica es una metaloenzima (proteína que contiene un ion metálico como cofactor), abundante en las paredes de los alvéolos y túbulos renales. CO 2 + H 2 O -> H 2 CO 3 Clasificación de las enzimas:
- Oxido Reductasas: Reacciones de oxidación – reducción. (deshidrogenasas)
- Transferasas: transferencia de grupos funcionales. (transaminasas)
- Hidrolasas: reacciones de hidrólisis (proteasas)
- Liasas: Eliminación de grupos para formar enlaces dobles. (carboxilasas->descarboxilaciòn).
- Isomerasas: Isomerización. (triosa fosfato isomerasa)
- Ligasas: Formación de enlaces acoplado con hidrólisis de ATP. (ADN ligasas) Cofactor: Ayuda a que encaje mejor el sustrato en la enzima: Coenzima: Mayoría derivan de vitaminas. Se encuentran unidas covalentemente al sitio activo de la enzima o próximo a él. Cosustrato : No está asociado a la enzima Grupo Prostético: Asociadas a la estructura proteica de las enzimas mediante uniones covalentes o no covalentes. Cofactores:
Especificidad de la enzima: Complementariedad geométrica y electrónica (atracción de cargas) entre enzima y sustrato->Fuerzas NO covalentes: interacciones Van der Waals, electrostáticas, puentes de hidrógeno.
Estado de transición:
Laboratorio:
Factores que afectan la actividad enzimática: ● Dependiente del pH ● Dependiente de la Tº ● Concentración de sustrato ● Inhibidores: suicidas, competitivos, no competitivos.
Sesión 2: Cinética enzimática
-Temperatura óptima para las enzimas: 36º-(36, 5º)-37º. A mayor temperatura, mayor número de colisiones. Si se da una mayor temperatura del rango habitual la proteína se desnaturaliza. Conforme disminuye la temperatura, la enzima se vuelve más lenta hasta que llega un máximo y se inactiva (si aumenta, pueden volver a realizar sus funciones catalíticas).
-pH óptimo: Debemos tener un pH ligeramente alcalino. En casi todas las enzimas celulares, la actividad enzimática ocurre con un pH de alrededor de 7,4 (7-8). La pepsina (pepsinógeno->pepsina) trabaja en un pH de 2.5 aprox (enzima extracelular: es llevada hacia la luz del estómago). La tripsina trabaja con un pH de
2.Regulación por retroalimentación: El producto actúa como inhibidor de la primera enzima esto impide: utilización innecesaria del primer sustrato y se evita la acumulación de intermedios y del producto final.
3.Regulación alostérica: (1) Control de la disponibilidad de la enzima (2) Control de la actividad enzimática
*Regulación alostérica negativa: regulador alostérico negativo cambia la conformación del sitio activo cuando se coloca en el sitio alostérico, no permite la unión de enzima-sustrato.
*Regulación alostérica positiva: el regulador alostérico positiva cambia la conformación del sitio activo para que el sustrato.
4.Regulación por modificación covalente: Enzimas que están inactivas hasta que se activan por unión covalente con otras moléculas y viceversa.
(a) Fosforilación y desfosforilación (b) Activación por ruptura proteolítica
un cimógeno debe perder una parte polipeptídica para que se active
Importancia médica: Citocromo C: síntesis de ácidos biliares Lactato deshidrogenasa: linfoma, infarto al miocardio.
SEMANA 3
Sesión 1: Carbohidratos CHO
-Forman el 65-75% de nutrientes -Principal fuente energética en la mayoría de células (energía inmediata) -Principal fuente de energía para músculos, cerebro. -La absorción de nutrientes en los enterocitos comienza a partir de moléculas asimilables, en este caso serían los monosacáridos. -Monosacáridos: azúcares simples se dividen: según su grupo funcional (cetosa: cetona, aldosa: aldehído) como la glucosa (aldohexosa), galactosa (aldohexosa) y fructosa (cetohexosa). -Isómeros: misma composición química pero diferente distribución. -El aldehído está más expuesto al ciclarse. -Azúcar reductor: aquel que puede sufrir una oxidación (debe tener un H y OH disponible) Disacáridos: sacarosa x (1, 2) no es un azúcar reductor. Lactosa b (1, 4) es un azúcar reductor. Maltosa x (1, 4) es un azúcar reductor. Polisacáridos: Almidón: amilosa y amilopectina // Unión lineal( x 1, 4) y unión ramificada(x 1, 6)
Digestión de carbohidratos Digestión: Proceso mecánico, químico y/o enzimático en el cual las macromoléculas van a pasar por procesos hasta convertirse en partículas asimilables (los carbohidratos serán convertidos en monosacáridos). (1)Amilasa salival o ptialina (digestión mecánica y enzimática): masticación y ayudado por la enzima x amilasa: rompe el enlace x 1, 4 del almidón liberándose glucosa, maltosa y dextrinas (2)Se forma el bolo alimenticio, llega al istmo de las fauces. (3)comienzan los movimientos peristálticos hacia llegar al estómago (digestión mecánica ayudado por el cambio de pH).para los carbohidratos no hay una digestión enzimática en el estómago porque no hay enzimas que degraden los carbohidratos se dan unos movimientos que generan la digestión mecánica y también puede generar la ruptura de enlaces(4)Pasa por el píloro, ingresa al duodeno y se forma el quilo/ampolla de
Sorbitol y glucosa: Efectos en el organismo El ATP y el citrato inhibe a la fosfofructoquinasa para que deje de producir piruvato. El citrato se encarga de la síntesis de lípidos. Galactosemia: La deficiencia de galactosa 1 fosfato uridiltransferasa trae muchas problemas renales, y es peor que la deficiencia galactoquinasa.
Monosacáridos de importancia médica: Fructosa Glucosa 6 fosfato puede irse a formar glucosa y luego glucógeno. Piruvato a partir de fosfoenolpiruvato. Se puede obtener Acetil COA y AcilCOA que es un precursor de lípidos. VLDL: se sintetiza en el hígado La vía del hígado graso no alcohólico: se obtienen lípidos en el hígado a partir de fructosa. 2 vías: obtener glucosa o almacenarlo en glucógeno. Si se inhibe la señalización de glucosa en células insulina independientes, no captaría glucosa, sintetizaría glucagon que favorece la gluconeogénesis. Lactosa Dispepsia: gases en el estómago Glicógeno Si no puede degradar glucógeno, genera una hipoglucemia y daño hepático.
Sesión 2: Metabolismo de carbohidratos
Lípidos: energía a largo plazo. Son utilizados después de utilizar TODOS los carbohidratos. Carbohidratos: energía corto plazo. Las proteínas proporcionan la misma energía que los carbohidratos. La glucolisis: -No requiere de O -Redox
-2 etapas: (a)inversión de energía (b)compensación de energía. Productos: 4ATPbrutos(2ATPnetos), 2NADH, 2 piruvato, 2H -3 puntos de control (irreversibles): hexoquinasa, fosfofructoquinasa, fosfopiruvato quinasa. -Ocurre en todas las células. -Después de obtener el piruvato puede hacer un proceso aeróbico o anaeróbico. -En el caso del eritrocito, solo hace glucólisis porque no tiene organelas.
Regulación de la glucolisis Si no tengo AMP, significa que no tengo ATP. A nivel de la hexoquinasa: va inhibir glucosa 6 fosfato. A nivel de la fosfofructoquinasa inhibe ATP y citrato A nivel de fos piruvato quinasa inhibe ATP y acetil CoA
Destino anaeróbico del piruvato Fermentación alcohólica y láctica: no podemos hacer la fermentación alcohólica (bacterias, levaduras: hongo unicelular), sola la láctica. Fermentación alcohólica: el ácido pirúvico se descarboxila a través de la piruvato descarboxilasa para obtener acetaldehído, luego un alcohol deshidrogenasa lo convierte a etanol a través del paso de NADH a NAD+. Fermentación láctica(músculos): la lactato deshidrogenasa hidrogena al ácido
es permeable, por lo tanto el NADH no puede transportar los electrones, entonces los transporta a partir de las lanzaderas. La dihidroxiacetona fosfato y el gliceraldehído 3 fosfato ambas son polares e ingresan por purinas (canales) El glicerol 3 fosfato deshidrogenasa (porque pierde sus hidrogeniones) mitocondrial actúa sobre el gliceraldehido 3 fosfato y transfiere sus electrones al fad oxidado y se vuelve fad reducido, este fad se va al segundo complejo de la cadena transportadora de electrones. Malaton aspartato y x cetoglutarato: pueden ingresar a través de la membrana interna mitocondrial El oxalacetato no es permeable a la membrana, por lo tanto no puede ingresar. Recibe 2 electrones de la enzima malato deshidrogenasa citoplasma y se convierte en malato. El malato solo transporta electrones al nad oxidado para convertirlo en NADH reducido y se va al complejo 1 mitocondrial (proteínas transmembranales que reciben los electrones en Cu+2 o\y Fe+2 porque son metales). El malato se convierte nuevamente en oxalacetato al transportar sus electrones al NADH, para esto tiene que transaminarse y convertirse en aspartato para recién salir de la mitocondria, el aspartato le da su grupo amino al x cetoglutarato y se convierte en glutamato. Cadena transportadora de electrones Los complejos actúan como canales de hidrogeniones, tienen sitios de Cu y fe para poder transportar electrones. Complejo 2 y citocromo c no son transmembrana pero igual tiene sitios de cu y fe para transportar electrones. El ATP sintasa y el último aceptor de electrones regulan la concentración de hidrogeniones. ATP sintasa: permite el ingreso de hidrogeniones para formar ATP. El O2: forma agua a partir de hidrogeniones El agua que se forma a partir del último aceptor de electrones se ubica en el intersticio. Especies reactivas de oxigeno (como OH) dañan lípidos, proteínas, hasta ácido nucleico. El complejo 3 es inhibido por una antimicina (antibióticos) porque antes era una bacteria (teoría endosimbiótica). El cianuro (CN) es transportado por la hemoglobina y afecta al complejo 4 de la mitocondria, generando que se detenga la cadena transportadora de electrones. La rotenona o amital inhibe al complejo 1. En los complejos se producen aniones superóxido que es regulado por la superoxido dismutasa que convierte en peróxido de hidrógeno.
Importancia Infarto: privación de oxígeno. Para que el corazón pueda contraerse los cardiomiocitos necesitan alta cantidad de energía. Lo que ocurre en la matriz mitocondrial es oxidativa. Si la cadena transportadora de electrones está afectada no se puede obtener la energía suficiente para contraerse, no hay buena estabilidad en el potencial. Los cardiomiocitos realizan necrosis en mayor cantidad. Hay privación de oxígeno porque no hay suficiente hemoglobina que transporte oxígeno. Cardiolipina, fosfolípido a nivel de membrana mitocondriales, también se ve afectado y por ende favorece a la disfunción de la mitocondria.
Síntesis y degradación del glucógeno Glucogenesis: la glucosa 1 fosfato se junta con UTP (uridil trifofasto), el UTP libera 2 foasfatos y me quedan 2 fosfatos, se forma uridil difosfato glucosa(UDP). El glucógeno sintasa le quita el grupo UDP y queda
glucosa, ese glucosa va aumentando y aumentado. El glucógeno tiene ramificaciones formadas por enzima ramificadoras( enlace 1,6: unión de ramificaciones y la lineal 1,4). Glucogenolisis: el glucógeno debe degradar los enlaces x 1, 4, luego una enzima desramificadora rompe las ramificaciones. Una glucógeno fosforilasa, fosforila y cambia a glucosa 1 fosfato, y por una enzima fosfoglucomutasa forma glucosa 6-fosfato y puede ingresar a glucolisis o puede obtener glucosa. GLUCOSA 6 FOSFATASA: EL MÚSCULO NO LA TIENE En la degradación de glucógeno se obtiene una glucosa libre cuando interviene la x1, glucosidasa. Control hormonal de glucógeno Células hepaticas: DAG e IP3 están relacionados en la activación de vía de la PKc, esta vía disminuye la síntesis de glucógeno. La glucógeno sintasa cuando está fosforilada está inactiva y activa cuando no tiene un grupo fosfato Músculo liso: estimulación hormonal: AMPc relacionada a proteína Gs, la PCA inhibe a la glucógeno sintasa. Control Hormonal de glucógeno Los hepatocitos no dependen de insulina para que ingrese glucosa. Si hay deificiencia enzimática de enzimas que degradan glucógeno, este no puede degradar por eso es normal. Enfermedad de pompe deficiencia de x 1, glucosidasa. Lisosomas contienen enzimas digestivas, las nazis a hidroliticas se liberan y esa hará un cambio de pH.
Glucógeno en cerebro A nivel del cerebro, se encuentran la laforina y la Malina, ellas forman un complejo que realizan una ubiquitinizacion, regulan a la glucógeno sintasa. Malina es una ubiquitina que tiene la función de degradación proteolitica. Al formarse el complejo, se une a una enzima, lo marca y lo degrada. Para que no se sintetice más glucógeno. Enfermedad de lafora: Deficiencia genética en esas proteínas, la glucógeno sintasa producirá más glucógeno y se habrá más a nivel de neuronas y se afecta el potencial electro químico a nivel de las neuronas y esto genera convulsiones. Viven alrededor de 10 años.
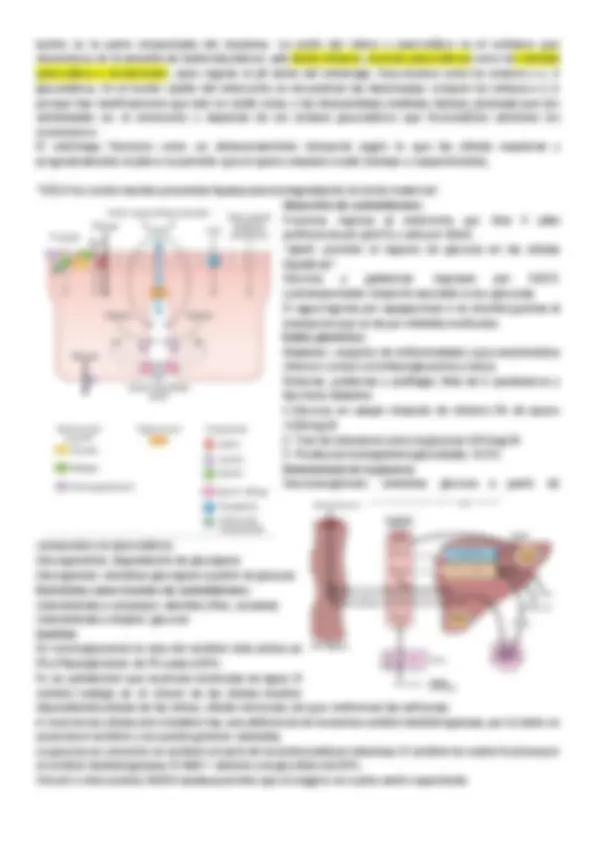
Gluconeogenesis : obtención de glucosa a partir de compuestos no glucosidicos. El hígado transporta glucosa hacia músculo. del músculo sale el lactato hacia el hígado y de lactato puede formarse glucógeno. Ciclo de la urea: elimina grupos aminos en exceso. -Alanina al perder su grupo amino pasa a ser piruvato, el grupo amino va hacia el ciclo de la urea. El piruvato pasa por gluconeogenesis y se convierte en glucosa. -En el tejido adiposo gracias a una lipasa degrada los triglicéridos a ácidos grasos y glicerol. El glicerol va hacia el hígado y puede convertirse en glucosa.
Regulación del metabolismo de carbohidratos