¡Descarga Psicobio 2a evidencia y más Apuntes en PDF de Psicobiología solo en Docsity!
Tema 1: Las células del sistema nervioso
1 Morfología 1.1. Soma, axón y dendritas Las neuronas son las unidades elementales en que se procesa la información en el SN. Pueden tener formas muy diversas pero la forma que tiene una neurona más común es : Soma: Cuerpo celular, envuelve al núcleo (donde se almacena la información genética). Axón: Prolongación que sale del cuerpo celular a través del cono axónico hasta la ramificación axónica donde están los denominados botones sinápticos o terminales Dendritas: Prolongaciones que salen del cuerpo celular con forma de árbol y que constituyen las principales áreas receptoras de la información que llega a la neurona. La sinapsis es la zona de transferencia de información de una neurona a otra. La mayoría de las neuronas tienen varios troncos dendríticos (dendritas primarias) que se ramifican varias veces, para así, incrementar la superficie de recepción de información de la célula. Las dendritas captan el mensaje lo conducen al soma. Algunas sinapsis se producen sobre pequeñas protuberancias de las dendritas denominadas espinas dendríticas. Las neuronas están especializadas en conducir un mensaje codificado en forma de impulso nervioso ( potencial de acción ). Esta propiedad se denomina conductividad, el axón es el que la permite. La información llega a las neuronas a través de las dendritas (entran, captan información de fuera). Se conduce la información a lo largo del axón y el botón terminal permite que este mensaje se transmita a otra célula, atravesando la sinapsis. Hay neuronas con axones muy largos, por lo tanto pueden transportar la información a puntos muy alejados del cuerpo. Tienen una resistencia muy grande porque están envueltas de nervios periféricos. 1.2. Fibras (axones) mielínicas y amielínicas: Cuando hablamos de fibras nos referimos a axones. Las axones mielínicos son axones recubiertos por un sustancia grasosa denominada mielina. Las fibras mielínicas forman la vaina de mielina (formada por lípidos = no conducen la corriente eléctrica), por lo tanto, la vaina es aislante, aísla eléctricamente de su entorno al axón. Es discontinuo, a lo largo de todo el axón hay fragmentos que no están recubiertos por mielina, los que si lo están son los internodos , y en cambio los que no lo están (están descubiertos, en contacto con el líquido celular) se denominan nodos de Ranvier. Las células que forman la vaina son diferentes en el SN periférico y el central. SN periférico: células de Schwann (internodo), forman la mielina. La célula de Schwann se alarga y se enrolla al rededor del axón, sobretodo la membrana, y la parte del núcleo quedaría a la parte más externa de la célula. SN central: oligodendroctos , células que tienen varias prolongaciones y cada una es la que formará la mielina de un segmento de axón. Por lo tanto, un oligodenrocito puede formar la parte mielina de diferentes axones. La mielina está formada por una prolongación de una célula y no para toda la célula, como las células de Schwann. En cambio, las fibras amielínicas no tienen mielina. Las fibras amielínicas también están formadas por células de Schwann y oligodendrocitos, dependiendo si es SN central o periférico. En los axones no hay mielina, por lo tanto, no están aisladas eléctricamente. La función de las células es fijar los axones en una determinada posición.
1.3 Estructura interna: Orgánulos y partículas Encontramos los mismos orgánulos que en las otras células eucariotas. Pero podemos distinguir de los orgánulos situados en el axón y otros situados en el soma y dendritas. En el soma y las dendritas encontramos los mismos orgánulos que en las otras células: RE rugoso y RE liso, aparato de Golgi, ribosomas, filamentos protéicos (forman la estructura, muy importante a las neuronas para su forma tan particular), mitocondrios ... Los filamentos proteicos de las neuronas ( citoesqueleto ) son: microtúbulos, neurofilamentos y microfilamentos. En esta parte de la neurona hay una gran acumulación de RE rugoso, denominada sustancia de Nissl , alrededor del núcleo. En cambio, en el axón no encontramos todos los orgánulos que encontramos en las dendritas y en el soma. Sí que hay mitocondrios (necesita mucha energía, más abundante que en las dendritas y soma), RE liso, filamentos proteicos... Pero en cambio, no hay RE rugoso, ni ribosomas (muy poco abundantes). Por lo tanto, en el axón prácticamente no hay síntesis de proteínas, las que necesita se han de sintetizar en el soma y transportarlos al axón, hasta donde se necesiten. Gracias al filamento proteico se puede transportar sustancias a lo largo del axón. Es su función más importante, el transporte axoplasmático, que permite transportar proteínas pero también vesículas y otros orgánulos. Este transporte puede ser: o Anterógrado : transporta sustancias del cuerpo celular hasta el botón terminal, al final del axón. o Retrógrado : transporta sustancias desde el final del axón hasta el soma (por ejemplo, sustancias viejas que se transportan a los ribosomas porque se degradan).
2. Clasificación 2.1Según su morfología Unipolares (monopolares) : Tienen una única prolongación que sale del cuerpo celular. En el caso de los mamíferos, las neuronas unipolares que encuentran son de un tipo denominado pseudo-unipolar. La única prolongación se bifurca en dos ramas, en un extremo se encuentra las dendritas y en el otro extremo se encuentran los botones terminales. También reciben el nombre de neuronas en T (por su forma). Este tipo de neuronas son sensonriales, captan la información sensorial para las dendritas y la transportan hasta los botones terminales. Bipolares : Tienen dos prolongaciones que salen del cuerpo celular. En una de las prolongaciones están las dendritas que captan la información, y en la otra prolongación está el axón con los botones terminales. También son sensoriales. Multipolares : Tienen muchas prolongaciones que salen del cuerpo celular. Una de las prolongaciones es el axón, y el resto son dendritas. Son las más abundantes y pueden ser de dos tipos: Multipolares de axón largo (Golgi tipo I) : Tienen un axón largo, generalmente mielínico. Multipolares de axón corto (Golgi tipo II) ; Tienen un axón corto, y en algunas ocasiones ni siquiera tienen axón, y normalmente el axón es amielínico. 2.2 Según su función Sensoriales (aferentes): Son neuronas que conducen la información sensorial desde los receptores sensoriales hasta el SNC. Este tipo de neuronas normalmente son neuronas en T (pseuomonopolares). Es decir, que son aferentes, llevan información de otros lugares hasta el SN central (estructura X). Motores (eferentes) : Son neuronas que conducen las instrucciones motoras dese el SNC hasta el órgano efector (músculo y glándulas, producen respuestas). Las neuronas motoras son multipolares Golgi I. Interneuronas : Se encuentran en su totalidad dentro del SNC. Son las más abundantes, se encuentran en el cerebro procesando la información.. Existen dos grupos: o Locales (multipolares Golgi II) : Comunican, transmiten su información a otras neuronas próximas, en la misma región local. o De proyección: (multipolares Golgi I) : Llevan la información de una región local hacia otra región cerebral.
Tema 2 : Fisiología de la neurona
1. Fisiología de la neurona 1.1 La membrana de la neurona La membrana neuronal es la estructura que define los límites de la neurona. Está formada por una doble capa de lípidos ( bicapa lipídica ). Esta composición hace que algunos elementos tengan más facilidad para atravesarla que otros. Aquí entran en juego las moléculas proteicas que actúan como canales iónicos que otorgan a la membrana la propiedad de semipermeabilidad. Estos canales pueden ser de dos tipos: pasivos y activos; y dentro de los activos se encuentran los dependientes de voltaje (cuya permeabilidad varía en función de cambios en el potencial de membrana) y los dependientes de ligando (que cambian su permeabilidad en respuesta a la presencia de ligandos, moléculas). La mayoría de los canales son selectivos, es decir, que permiten el paso selectivo de un único ion. Conceptos importantes: Semipermeabilidad Canales iónicos activos y pasivos (¿Cuál es la diferencia y cuando están abiertos?) 1.2 El potencial de membrana Tanto dentro como fuera de la neurona se encuentran una serie de moléculas ( iones) de diferentes cargas (positivos→ cationes; negativos → aniones ) las cuales están repartidas de manera no uniforme, creando así una distribución asimétrica de los iones. Hay moléculas que no pueden atravesar la membrana de ninguna manera (carecen de cualquier tipo de canales). Son un ejemplo los aniones orgánicos que produce la misma célula, para los cuales la membrana es completamente impermeable. Así pues, los elementos que sí pueden traspasarla han de intentar compensar esta diferencia de cargas. En esta diferencia de cargas tienen lugar dos fuerzas opuestas entre sí: la fuerza química y la fuerza eléctrica. Fuerza química: El movimiento que realizan las moléculas al desplazarse de zonas de mayor concentración a zonas de menor concentración. Fuerza eléctrica: El movimiento que realizan las moléculas en función de la atracción o repulsión que surge de la diferencia o igualdad de sus cargas eléctricas. Algo muy importante también a tener en cuenta es que los movimientos iónicos no solo dependen de estas dos fuerzas, sino de la permeabilidad. Cuando unos iones estén separados por una membrana completamente permeable, estos la atravesarán libremente hasta alcanzar el equilibrio. Por el contrario, si están separados por una membrana semipermeable, los iones que puedan cruzarla se dispondrán de forma asimétrica para compensar los iones que no pueden hacerlo, dando como resultado una diferencia de potencial tanto eléctrico como químico. El potencial de membrana es entonces, esta diferencia de potencial eléctrico que se origina por esta distribución no equitativa de los iones. Según en qué situación se encuentre la neurona, a este potencial de membrana se le llamará de diferentes maneras. -Si la neurona no recibe ni envía ningún estímulo, se le llamará potencial de reposo. -Si la neurona recibe un pequeño estímulo, pero no es suficiente para que llegue al umbral de descarga, se le llamará potencial local. (Pierde intensidad) -Si la neurona recibe un pequeño estímulo suficiente para llegar al umbral de descarga, se le llamará potencial de acción. (No pierde intensidad)
Canales
iónicos
Pasivos Activos
Dependientes
de voltaje
Dependientes
de ligandos
Conceptos importantes: Distribución asimétrica (¿Por qué existe?) Fuerza química y eléctrica (Qué son y cómo actúan) Permeabilidad (¿Qué es y qué importancia tiene?) Potencial de membrana (¿Qué es, como se crea y de que maneras puede variar?) 1.2.1 Potencial de reposo Cuando la neurona se encuentra inactiva, es decir, ni recibe ni envía información, tienen una diferencia de potencial a través de su membrana de -70mV (el interior de la célula es considerado negativo ya que contiene un mayor número de cargas negativas). Por lo tanto, el potencial de reposo es de -70mV. Los iones que encontramos a ambos lados de la membrana neuronal son: Iones de cloro: Cl- Iones de sodio: Na+ Iones de potasio: K+ Luego están los aniones orgánicos (A-): principalmente proteínas de carga negativa que se encuentran en el interior. Debido a que la membrana es semipermeable la distribución de estos iones a ambos lados de la membrana no es simétrica. Los aniones orgánicos solo se encuentran en el fluido intracelular. El resto se encuentran tanto en el compartimento intracelular como en el extracelular, pero recordamos que su distribución es desigual. -El K+^ se encuentra principalmente en el medio intracelular. -El Na+^ i el Cl-^ se encuentran principalmente en el medio extracelular. Antes hemos dicho que existen canales pasivos y activos en la membrana que ayudan a que los iones puedan traspasar la membrana. En el potencial de reposo cobran más importancia los canales pasivos ya que son los que están siempre abiertos independientemente de si les llegan estímulos o no. Dado que hay muchos canales pasivos para el K+, la membrana será mucho más permeable al potasio que al sodio en estado de reposo. Así pues, el K+^ puede atravesar la membrana a favor de su gradiente electroquímico (según las fuerzas químicas y eléctricas) y distribuirse de manera asimétrica contrarrestando los iones que no la pueden atravesar (en especial los aniones proteicos). El potasio trata de llegar al equilibrio, que es ese momento en que las fuerzas eléctricas (que lo empujan a quedarse) y las químicas (que lo empujan a salir) se igualan. El potencial de equilibrio del K+^ es de -1100 mV. Sin embargo, el valor del potencial de reposo es de -70 mV, por lo que el potasio no se encuentra en equilibrio. ¿Por qué? Pues porque en reposo la membrana no es totalmente impermeable al Na+, de manera que la pequeña entrada de sodio que se produce contrarresta la salida de K+. El potencial de membrana de la neurona depende, en gran medida, del potencial de equilibrio del elemento cuya permeabilidad con la membrana, en ese momento, sea mayor. Por ejemplo, en el potencial de reposo, el ion con más permeabilidad es el potasio, por eso este potencial se parece tanto al de equilibrio de dicho elemento. Conceptos importantes: Potencial de reposo (¿Qué es / en qué situación se da?) Potencial de equilibrio del K+^ (¿Cómo influye en el potencial de reposo y por qué?) Potencial de membrana ¿Estímulo? Sí ¿Llega al umbral de descarga? Sí Potencial de acción No Potencial local No Potencial de reposo
CADV (¿Cuáles son los que participan?) Fases del potencial de acción (¿Cuáles son y que sucede en cada una de ellas?) Fundamentos del potencial de acción
¿Por qué se produce la despolarización durante el potencial de acción?
Para que se desencadene un potencial de acción es necesaria una estimulación intensa (que llegue al umbral) para que produzca una despolarización de la membrana. En este momento se abren algunos canales de Na+^ dependientes de voltaje: el sodio se precipita hacia el interior, impulsado tanto por fuerzas químicas como eléctricas, y se produce una rápida despolarización (lo cual hace que aun se abran más canales de sodio, que producen más despolarización). El inicio del potencial de acción se corresponde a un aumento de permeabilidad del sodio debido a la apertura de sus canales. Los canales de sodio se abren con mucha rapidez en comparación con los de potasio. El valor del potencial de equilibrio Na+^ es de +55 mV, sin embargo el potencial de acción tiene un pico (punto de mayor despolarización) de +40 mV, con lo que el sodio nunca llega a alcanzar el equilibrio.
¿Por qué el potencial de membrana no llega al valor del potencial de equilibrio del Na+?
Hemos dicho antes que los canales de sodio se abren más rápido que los de potasio. Esto significa que en cuanto llega el estímulo, los primeros que se abren son los de sodio, provocando más diferencia de potencial, que a su vez abre más canales. Este aumento de potencial que produce la apertura de canales de Na+^ hace que se abran los canales de potasio antes de que el sodio llegue a su potencial de equilibrio. El sodio se encontraba en mayor cantidad en el exterior de la célula, y el potasio en mayor cantidad en el interior. Con la apertura de CADV de sodio, este entra dentro de la célula despolarizándola y haciéndola más positiva. A continuación se abren los CADV de potasio, que sale de la neurona haciendo su interior negativo.
¿Por qué se produce la repolarización?
Los CADV de sodio tienen rapidez tanto para abrirse como para cerrarse. Esto conlleva que cuando el sodio está a punto de alcanzar su potencial de equilibrio, se abren los canales de potasio, y a su vez, se cierran los de sodio porque “se les ha acabado el estímulo”. La repolarización consistía en que el potencial de membrana recuperaba su potencial de reposo, es decir, recupera la negatividad que había ganado con la entrada de sodio. Los iones de K+^ salen impulsados tanto por fuerzas eléctricas como químicas. -Fuerzas químicas: La concentración de potasio es mayor en el interior, por lo que esas fuerzas lo impulsan a salir. -Fuerzas eléctricas: La entrada masiva de Na+^ ha hecho que el interior se vuelva positivo, lo que crea repulsión al ión positivo del K+, con lo cual, esta fuerza también lo impulsa a salir para perder positividad y recuperar el potencial de reposo. El potencial de equilibrio del potasio es de -110 mV. Esto provoca que cuando la neurona recupera su potencia de reposo (-70 mV), esta siga ganando negatividad para intentar llegar a su equilibrio. (Coloquialmente se podría decir que el potasio ha cogido carrerilla, y aunque ha llegado al potencial de reposo, no puede parar de golpe, hecho que produce una hiperpolarización). ¿Por qué se produce la hiperolarización? Tal y como acabamos de ver, el flujo de potasio aún sigue activo ya que sus canales aún no se han cerrado. Este hecho hace que se produzca la hiperpolarización: el interior se vuelve más negativo con respecto el potencial de reposo. Pero poco después se cierran los canales de potasio haciendo que los valores se vayan recuperando lentamente hacia el valor del potencial de reposo.
Conceptos importantes: (Básicamente, saber responder a las preguntas de los puntos anteriores) Canales de Na+^ y canales de K+ Es importante remarcar y destacar las diferencias entre los canales de sodio dependientes de voltaje y los canales dependientes de voltaje de potasio que participan en el potencial de acción. Los canales de Na+^ se abren y cierran con rapidez , mientras que los de K+^ se abren y cierran con más lentitud. Antes hemos hablado de la bomba de Na+-K+^ cuya función es expulsar tres iones de sodio hacia el exterior de la neurona e introducir dos de potasio. Cuando ha acabado el potencial de acción, esta bomba cobra especial importancia ya que aumenta su actividad para restablecer las concentraciones iniciales y que han sido alteradas. 1.4 Codificación de la información: El potencial de acción es la unidad básica del lenguaje, la forma en que el SN codifica y transmite información; es la unidad básica de transmisión de información en el SN. Para que se produzca este potencial de acción, hemos visto que es necesario que el estímulo llegue al umbral de descarga para que así se abran los canales de Na+^ dependientes de voltaje. El potencial de acción sigue la ley del “todo o nada” , que postula que un potencial de acción se da o no se da: si se desencadena se transmitirá a lo largo de todo el axón conservando siempre el mismo tamaño o intensidad. (Imaginemos que el potencial de acción es como el gatillo de una pistola. Puedes apretarlo un poco sin llegar a disparar, y este volverá a su sitio sin haberse producido el tiro. Pero si lo presionas hasta cierto punto, la pistola se dispara). Así pues, el potencial de acción se da con toda su amplitud llegando a los +40 mV como máximo, aproximadamente. 1.5 Periodos refractarios Cuando en un punto de la membrana se ha producido un potencial de acción, no se puede volver a producirse en otro en el mismo lugar inmediatamente , sino que tiene que transcurrir un determinado periodo de tiempo. -Periodo refractario: Es el tiempo que tiene que transcurrir tras un potencial de acción para que un estímulo umbral sea capaz de producir un nuevo potencial de acción en el mismo punto. -Periodo refractario absoluto: Es la parte del periodo refractario en la que por muy alta que sea la intensidad del estímulo no se produce un potencial de acción. -Periodo refractario relativo: Es la parte del periodo refractario en la que, si la estimulación es lo suficientemente intensa (por encima del estímulo umbral), se consigue producir un nuevo potencial de acción; por lo tanto, es un periodo en el que el umbral de descarga es más elevado.
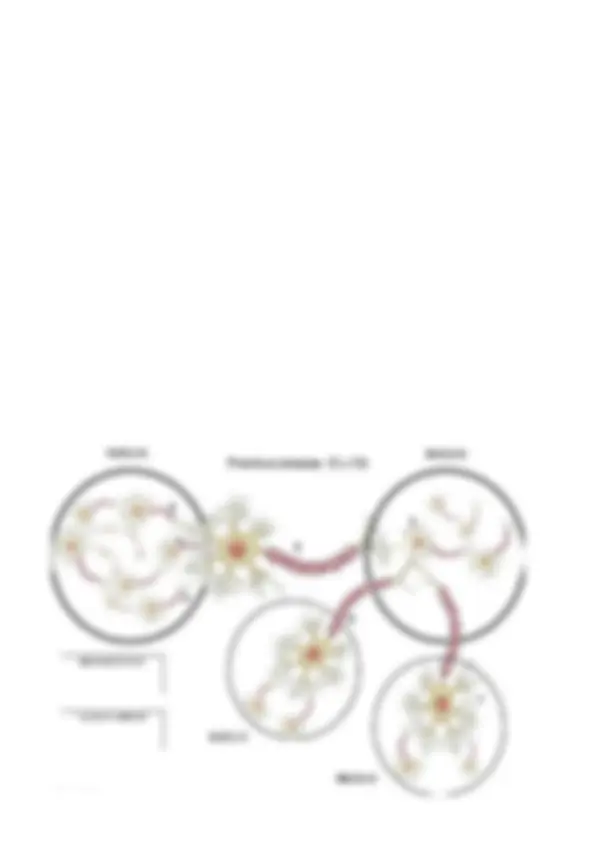
Sinapsis química : La información se transmite de un lugar a otro mediante una sustancia química que se fabrica en el interior de la neurona denominada neurotransmisor. La membrana presináptica tiene unas vesículas sinápticas donde se encuentran los neurotransmisores. La membrana postináptica tiene una región densa/gruesa llamada densidad postsináptica. Dentro de ella hay proteínas que actúan como receptores postsinápticos y también hay canales iónicos controlados por ligando (normalmente cerrados que se abren al unirse un ligando al receptor). Cuando llega el PA al botón terminal provoca que se liberen moléculas de neurotransmisores en el espacio sináptico. Estas moléculas de neurotransmisor llegan a la neurona postpsináptica y se unen a los receptores postsinápticos. Esta unión hace que se abran algunos canales controlados por ligando. Al abrirse estos canales, cambia la permeabilidad de la membrana para determinados iones (ahora podrán entrar mejor). Esto provocará un cambio en el potencial de la membrana postsináptica: o se da una despolarización o una hiperpolarización. Este cambio será la respuesta postsináptica (como consecuencia de la transmisión de información). Si la respuesta postsináptica es una despolarización se denomina sinapsis excitadora, si es una hiperpolarización se denomina sinapsis inhibidora. Otro tipo de transmisión sináptica es la sinapsis de paso : En el cual, todo el axón presenta varicosidades que funcionan de forma parecida al botón terminal, tienen vesículas sinápticas y toda la maquinaria para liberar neurotransmisores. Cuando llegan PA a las varicosidades provoca la liberación de neurotransmisores. La diferencia es que no se liberan únicamente NT desde el final del axón a un punto concreto de la membrana postsináptica, sino que se liberan desde diferentes puntos y llegan a puntos diferentes también.
2. Mecanismos básicos de transmisión sináptica química 2.1 Liberación del neurotransmisor (NT) El botón terminal de la neurona presináptica libera NT, lo hace desde la zona activa, que es una zona especializada de la membrana presináptica desde donde las vesículas sinápticas liberan el NT al espacio sináptico. La llegada del PA supone una despolarización provocando que se abran los canales de Ca+ controlados por voltaje. Al abrirse estos canales, el Ca2+ (de ion +) tiende a entrar hacia dentro del botón terminal. Estos iones de Ca2+ son los responsables de que se libere el NT al espacio sináptico ya que el Ca2+ se
une a las vesículas sinápticas y hace que estas se muevan hacia la zona activa y allí liberen el NT. Hay dos maneras de liberación: Exocitosis : En este proceso la membrana de la vesícula se funde con la membrana plasmática. Se abre la membrana y lo que hay en la célula puede salir libremente al exterior, al espacio sináptico. Endocitosis : La membrana de la vesícula vuelve a ser absorbida por el interior del botón terminal (reciclaje de vesículas). Esta podrá servir de nuevo como vesícula sináptica. La cantidad de NT que se libera depende del PA que llegue al botón terminal:
- Pocos PA: pocos canales de Ca+ → entran pocos iones de Ca+ → pocas vesículas liberan NT.
- Muchos PA: muchos canales de Ca+ → entran muchos iones de Ca+ → muchas vesículas liberan NT. 2.2 Receptores postsinápticos El NT liberado al espacio sináptico se difunde por él y llega así a la membrana postsináptica En esta membrana pueden las moléculas de NT unirse a los receptores postsinápticos. Esta unión de los NT a los receptores postsinápticos provoca la apertura de los canales controlados por ligando. Al abrirse estos canales cambia la permeabilidad de la membrana para determinados iones. Se produce un intercambio de iones y el hecho de que entren o salgan iones +/- provoca un cambio en el potencial de membrana y con ello una respuesta/efecto postsináptico. Los receptores postsinápticos tienen moléculas proteicas. Características: Alta especificidad frente a un tipo concreto de NT. El mismo NT es reconocido por diferentes tipos de receptores. Es decir, un mismo NT puede activar diferentes receptores. El mismo NT puede tener diferentes efectos cuando se une a diferentes receptores. Esto es debido a que los diferentes receptores dan lugar a la apertura de diferentes canales y con ello da lugar a producirse diferentes efectos (hiperpolarización, despolarización…) 2.2.1 Tipos de receptores en función del mecanismo por el cual se abren Ionotrópicos : Son complejos receptor-canal (el mismo complejo proteico contiene el canal iónico y el receptor del NT). Se une al NT al receptor y el canal se abre directamente. Metabotrópicos : La unión del receptor con el NT desencadena una cadena de procesos metabólicos que culminan con la obertura del canal iónico. Esta cadena puede ser corta o larga: Corta : Sólo interviene una proteína (proteína G). Cuando se une el NT con el receptor se activa la proteína G y ésta hace que se abra el canal iónico provocando así un cambio de permeabilidad en la membrana y un efecto postsináptico. Larga : Es lo más común. Interviene un 2º mensajero (mensajero intracelular), siendo el 1r mensajero el NT. Hay diferentes tipos de 2º mensajeros:
- AMPc (ademosín monofosfato cíclcico): Se sintetiza a través del ATP.
- Diacilglicero (DG) e IP3.
- Ácido araquidónico COMPLETAR: no sé cómo desarrollarlo, en los
Si la suma de PEPs y PIPs no llega al umbral de descarga se producirá un potencial local (el cual se va debilitando) y no llegará a trasmitir el mensaje a la siguiente neurona. ¿Cómo se suman las sinapsis?
- Sumación temporal : Es la suma de los efectos de 2 o más PA que llegan a la misma sinapsis en momentos cercanos. La suma da lugar a una despolarización más grande y más duradera. Es más probable que se pueda producir un PA ya que puede llegar al umbral de descarga.
- Sumación espacial : Es la suma de los efectos de 2 o más PA que llegan al mismo tiempo pero en diferentes lugares de la neurona postsináptica. La suma definitiva es menor. a. Si hay dos sinapsis excitadoras: Despolarización b. Si hay 1 sinapsis inhibidora: Hiperpolarización 2.4 Mecanismos de inactivación del neurotrasmisor La inactivación es un proceso por el cual se elimina el NT del espacio sináptico. Hay dos mecanismos de inactivación:
- Degradación enzimática: Hay una enzima en el espacio sináptico capaz de degradar el NT, la puede convertir en otra molécula y de esta manera ya no es capaz de activar el receptor.
- Recaptación: El NT es inactivado mediante un mecanismo de recaptación (mecanismo de transporte activo de alta afinidad) que, recapta el NT y lo transporta al interior del botón terminal (desaparece del espacio sináptico) pero sí seguirá siendo útil. Al ser un trasporte activo conlleva un gasto de energía. Y es de alta afinidad porque reconoce rápidamente el NT. Las sinapsis están rodeadas/envueltas de células gliales. Así, el NT también puede ser llevado a las membranas de estas células gliales. Podemos decir, que la inactivación de los NT puede deberse a: inactivación enzimática, recaptación, o ambas simultáneamente. 3. Modulación sináptica 3.1 Receptores presinápticos También hay receptores presinápticos que modulan la cantidad de NT que se libera cuando llega un PA (podrán hacer que aumente o disminuya la cantidad de NT liberada). Normalmente es un efecto de modulación inhibidora, es decir, que disminuya la cantidad de NT. Estos receptores presinápticos puede ser: autoreceptores o heteroreceptores. Autoreceptores : Son receptores presinápticos que se activan cuando se les une el NT que se libera de la propia neurona, es decir, afectan directamente sobre la liberación del NT o actúan sobre su síntesis. El NT se une y después vuelve al espacio sináptico. Cuando se une una molécula de NT al autoreceptor, este hará que se libere más o menos NT, y los PA se transformarán: hará que los
siguientes PA se inhiban o aumenten al sacar NT (normalmente es un proceso de inhibición. Los NT puede, de alguna manera, autorregularse. Heteroreceptores : reconocen otro NT diferente al que libera su propia neurona. A menudo reconoce el NT que libera una 3ª neurona que establece sinapsis axoaxónica sobre esta neurona principal. El efecto será que se modulará la liberación del NT disminuyéndolo. 3.2 Inhibición y facilitación presináptica 3.2.1 Inhibición presináptica: Si un PA llega al botón terminal, se abrirán canales de Ca+ con lo que se liberará una cantidad de NT al espacio sináptico y esto hará que se produzca un PA. Llega un PA en una 3ª neurona, desde el botón terminal libera NT, actúan los heterorecetores presinápticos. La activación de los receptores presinápticos tendrá un efecto inhibidor sobre los canales de Ca+ (se abrirán menos canales de Ca+ controlados por voltaje). Mientras dura este efecto inhibidor sobre los canales de Ca+ al interior del botón terminal (ya que están inhibidos), entra menos Ca+, se libera menos NT de lo normal al espacio sináptico. Por tanto, el PEP que se produce en la membrana postsináptica es más pequeño de lo normal Esto hace que la cantidad de información que se trasmite por sinapsis excitadora, pero la neurona postsináptica no queda inhibida. 3.2.2 Facilitación presináptica: Cuando llega un PA a la sinapsis axoaxónica, el NT que se libera activará los heteroreceptores presinápticos, y ahora la activación de estos facilitará la apertura de los canales de Ca+ controlados por voltaje. Si mientras dura este efecto llega un PA a esta sinapsis principal, éste PA hará que entre más Ca+ a interior del botón terminal (más NT también). El PEP que se produce en la membrana postsináptica es más grande de lo normal. Se transmite más información. [Autoreceptor (activación del receptor) vs recaptación (transporte NT)] 4 .Mecanismos de plasticidad sináptica Es la capacidad de las sinapsis de modificarse, conllevan a una serie de cambios bioquímicos y/o estructurales en las conexiones entre las neuronas que proporcionan al organismo la capacidad de adaptación al entorno (cambios comportamentales), ésta no es ni buena ni mala, es una propiedad intrínseca: Cambios en el entorno → Cambios en el SN → Cambios en la conducta. Podemos decir, entonces, que tenemos plasticidad conducta porque tenemos plasticidad sináptica. Hay dos tipos de cambios: Cambios no genómicos: implican la modificación de proteínas ya existentes pero, no hay una nueva síntesis. Menor latencia de inicio. Poco estables y normalmente implican cambios a corto plazo.
Acetilcolina (Ach)
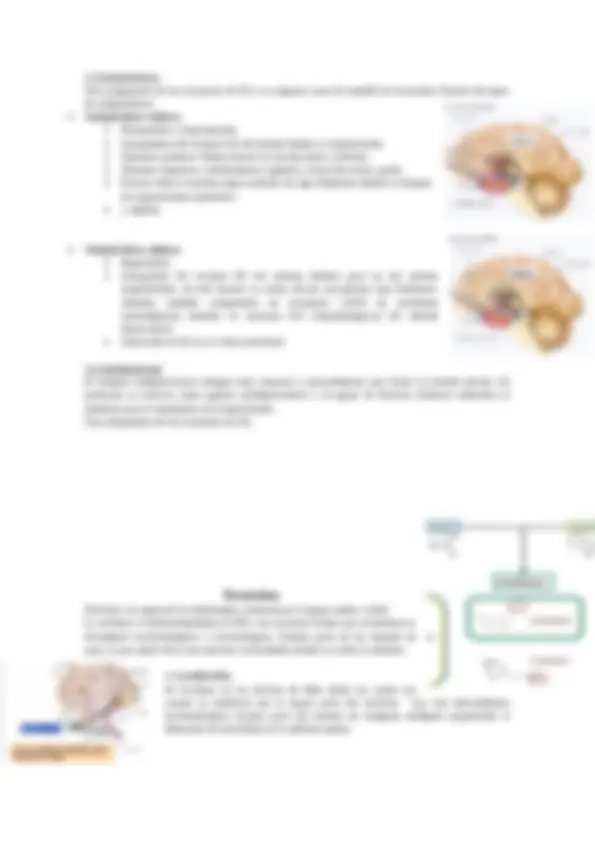
1. Localización y principales funciones: En el SNC: Los somas de las neuronas colinérgicas se encuentran en : o Área Septal Activación cortical, Atención, Aprendizaje y Memoria o Núcleo Basal de Meynert y o Tronco del encéfalo sonido paradoxal En el SNP: o Somático (SNS): Se encarga de obtener y transmitir la información detectada por los sentidos y de enviar instrucciones que permiten el movimiento voluntario de los músculos. Unión neuromuscular -> Contracción de los músculos esqueléticos o Autónomo o vegetativo (SNA): Rama parasimpática: Se encuentra activa cuando el cuerpo está en estado de reposo, estimula la peristalsis, relaja el mitocondrias, contrae los bronquios, etc. Rama simpática: Prepara al organismo para la actividad física, aumentando la frecuencia cardíaca, dilatando los bronquios, contrayendo el recto, relajando la vejiga… 2. Síntesis e inactivación La acetilcolina (ACh) está formada por acetato y colina, que se unen mediante la acción de la enzima acetilcolinatransferasa, que al mismo tiempo necesita Acetil-Coenzim A para poder transferir el acetato. Esta acción tiene lugar en el citoplasma neuronal de los terminales nerviosos. La formación de acetilcolina está limitada por la concentración intracelular de la colina, y esta viene determinada por la recaptura de colina dentro del terminal nervioso. Las neuronas no pueden sintetizar colina de nuevo y, por lo tanto, es suministrada desde el citoplasma. La mitad de colina empleada en la síntesis de ACh se cree que proviene directamente de Ach reciclada o liberada, hidrolizada a colina por la colinesterasa.
Otra fuente de colina es la ruptura de la fosfatilicolina, que puede aumentar en respuesta de la liberación de ACh. Una vez liberada la hendidura sináptica, la acetilcolina se une durante un tiempo muy corto a sus receptores sinápticos antes de ser degradada por la acetilcolinesterasa (AChE) que está concentrada en la hendidura se une específicamente a la acetilcolina y la rompe en dos moléculas, liberando los precursores de su síntesis, es decir, el acetato y la colina. Se inactiva por degradación enzimática.
3. Receptores: Receptores nicotínicos: Son receptores ionotrópicos. Cuando se fijan al ACh o a la nicotina se abre un poro que permite el paso a través de su bomba Na+-K+. Se caracterizan por una respuesta rápida que actúa mediante la despolarización directa de la membrana postsináptica en activar los canales de Na+. - Agonistas: Incrementan la actividad del receptor: Nicotina, N-acetilcolina, muscarina… - Antagonista: curare. - Efecto postsináptico: Despolarización rápida - Localización: Unión neuromuscular, sinapsis preganglionar del SNA y en algunas zonas del SNC (PEP). Receptores mucarínicos: Son receptores metabotrópicos que están involucrados en la memoria, el aprendizaje, la depresión o manía. Son de respuesta lenta. - Agonistas mucarínicos: Muscarina, Metalcolina, Carbacol… - Antagonistas mucarínicos: Atropina, Escopolamina… - Efecto postsináptico: Despolarización lenta y prolongada (algún subtipo puede producir (PIP). - Subtipos: Del M1 al M5. - Localización: SNC y sinapsis proganglionar (SNA parasimpático). 4. Farmacología Hemicolnium: Disminuye la síntesis de acetilcolina ya que no permite la entrada de colina en el botón terminal. Toxina botulínica: Disminuye la liberación de acetilcolina (extremadamente tóxica, parálisis muscular, se utiliza en estética (Botox)). Veneno de la viuda negra: Aumenta la liberación de acetilcolina provoca contracciones musculares, taquicardias, aumento de secreciones y dolor generalizado). Inhibidores del AChE (enzima que degrada la Ach): o Reversibles: neostigmina: antídotos, miastenia grave, Alzheimer. o Irreversibles: pesticidas, armas químicas. o Gas Sarín: Disminuya la inactivación de Ach. Ésta se acumula sobre la placa motora y se sobreactivan los receptores musculares. Esto provoca la contracción tetánica de la musculatura que puede causar la muerte por parálisis respiratoria. Implicaciones clínicas: Alzheimer: Acumulación de amiloide y “cabdells” neurofibrilares, formación de placas seniles. Se trata con anticolinesterásicos.
COMPT (Catecol-orto-metil transferasa): Enzima que degrada el neurotransmisor fuera, en el espacio sináptico.
3. Localización (DA): En el SNC: Vía negro-estriada : Va desde la sustancia negra hasta la estriada. Esta vía está relacionada con el control de los movimientos precisos. Está relacionada con el Parkinson, una degeneración de la sustancia negra está relacionada con la enfermedad del Parkinson. Vía mesocorticolímbica : Desde el área tegmental ventral al sistema límbico y corteza. 4. Funciones e implicaciones clínicas: Control de los movimientos preciosos y participación en el movimiento motor (sistema negroestriado). Circuitos neurales de la recompensa y el placer (sistema mesocorticolímbico). Los reforzadores naturales y artificiales activan este sistema. Además, las drogas adictivas acaban activando la liberación de dopamina en los núcleos colindantes (circuito mesolímbico). Esquizofrenia. Produce un aumento de DA en el sistema límbico y un decrecimiento de este en la corteza prefrontal (las vías ubicadas en estos lugares intervienen en importantes funciones emocionales, motivacionales y cognitivas). Parkinson: Alteración en las neuronas dopaminérgicas que intervienen en la motricidad. Esta patología se caracteriza por la rigidez, la dificultad en el movimiento y el temblor en reposo.
Noradrenalina
1. Localización: Junto a la Adr se encuentra en el SNC NA en el SNA: neuronas postganglionares de división simpática NA y ADR: hormonas suprarrenales: Glándula suprarrenal: Corteza suprarrenal y médula suprarrenal (donde hay más ADR que NA) 2. Funciones e implicaciones clínicas: Aprendizaje y memoria Procesos de vigilia y alerta Regulación de respuestas vegetativas (homeostasis) Regulación de los estados de ánimo: depresión. Respuesta del organismo ante el estrés (situación de emergencia):