¡Descarga Todo sobre Macrófagos y más Monografías, Ensayos en PDF de Inmunología solo en Docsity!
Macrófagos
Resumen Los macrófagos son células de el sistema inmune innato y representan un componente importante de la primera linea de defensa contra patogenos y celulas tumorales. Aquí se describen sus diversas funciones en la inflamación y la defensa tumoral, y se presentan los mecanismos, herramientas, vías de activación y estados aplicados. El enfoque principal está en el papel y el origen de las especies reactivas de oxígeno (ROS), las importantes vías de señalización TLR/NF-κB y la polarización M1/M2 de los macrófagos.
Introducción
Este texto de lección ha sido escrito para presentar la naturaleza y función de los macrófagos a los estudiantes de bioquímica, farmacia, química y materias similares. Los macrófagos son células importantes del sistema inmunitario innato o inespecífico presentes en todos los vertebrados. Como todas las células inmunes, los macrófagos se derivan de una célula madre hematopoyética pluripotente en la médula ósea. A partir de esto, por un lado, se desarrollan linfocitos y células asesinas naturales (NK) y ciertas células dendríticas a través de una célula precursora linfática. Por otro lado, los eritrocitos, plaquetas y precursores de granulocitos y macrófagos se desarrollan a través de una célula progenitora mieloide; los monocitos, neutrófilos, basófilos o eosinófilos surgen de los precursores antes mencionados. Los monocitos que migran de la circulación al tejido maduran y se convierten en macrófagos (Figs. 1, 2). En la circulación, el rango normal de monocitos está entre el 2% y el 8% del recuento de glóbulos blancos, lo que corresponde a 200-800 monocitos por microlitro de sangre. Los monocitos son células redondas con un núcleo en forma de riñón y tienen un tamaño de 25 a 30 μm. Al madurar en macrófagos, asumen diferentes formas según su funcionalidad (ver “Polarización de macrófagos M1/M2”) y, por lo general, se vuelven aún más grandes. Los macrófagos se pueden caracterizar por marcadores de superficie o marcadores celulares. Estos están bien establecidos, por ejemplo, para distinguir los diferentes fenotipos resultantes de la polarización de macrófagos (Tabla 3). Sin embargo, debido a su ascendencia común, estos marcadores se comparten en gran medida con las subpoblaciones de células dendríticas o microglía correspondientes [7, 36, 63, 68].
Fig. 1
Hematopoyesis. Las células precursoras mieloides y linfáticas se desarrollan a partir de células madre hematopoyéticas pluripotentes y autorrenovables. A partir de estas células progenitoras, se desarrollan linfocitos y varias células mieloides, que representan el arsenal celular del sistema inmunitario (con permiso [75]) Tabla 1
Tejido Macrófago Hígado Células Kupffer
Pulmón Macrófagos alveolares, neumocitos tipo II Piel células de langerhans Cavidades serosas macrófagos serosos Tejido conectivo Histiocitos Articulaciones/cartílago Células sinoviales (tipo A) Hueso Osteoclastos Riñón macrófagos de mesangio Cerebro Microglía
Desde la mitad de la gestación a lo largo de la vida, los macrófagos están presentes en todos los tejidos. Dependiendo del tejido en el que residen, los macrófagos poseen diferentes formas, por ejemplo, células de Kupffer en el hígado o células de Langerhans en la piel (Tabla 1). En ratones, los macrófagos presentes en muchos tejidos, incluidos la piel, el hígado, los riñones y el cerebro, se originan en el saco vitelino o en el hígado fetal. En la edad adulta, en ausencia de factores estimulantes, los monocitos circulantes no contribuyen sustancialmente al contenido tisular de macrófagos. Otros tejidos, como el corazón y el tracto gastrointestinal, reclutan precursores monocíticos en mayor medida donde los monocitos inmigrantes se diferencian en macrófagos residentes. En la vida adulta, los macrófagos residentes en muchos tejidos representan una mezcla de células que se originan durante el desarrollo y de precursores monocíticos circulantes. Sin embargo, en humanos adultos, debido a la vida prolongada, la mayoría de los macrófagos tisulares parecen ser reclutados de la circulación [4, 24, 46]. Los macrófagos funcionan en la defensa
contra patógenos y en la eliminación de células viejas, senescentes o muertas, pero también cumplen funciones importantes en la homeostasis y reparación de tejidos, por ejemplo, cicatrización de heridas y regeneración muscular [25]. Existen varias poblaciones de macrófagos que desempeñan funciones distintas y no redundantes en la fibrosis, la reparación y la regeneración de tejidos [26, 69]. Aún queda por establecer, sin embargo, si los macrófagos residentes o reclutados representan subconjuntos funcionalmente diferentes o son capaces de asumir todos los estados posibles dependiendo del microambiente celular/tejido.
Los macrófagos son una parte importante de la primera línea de defensa. Durante la infección, inflamación o lesión tisular, pueden seguir señales quimiotácticas para migrar al tejido dañado/sitio inflamatorio, donde ingieren patógenos y desechos celulares a través de un proceso llamado fagocitosis y los digieren en fagolisosomas, que resulta de la fusión de fagosomas con lisosomas. Los lisosomas son orgánulos encerrados en una membrana que contienen un conjunto de hidrolasas capaces de descomponer las diversas clases de macromoléculas. Esta degradación no solo contribuye a la eliminación de patógenos y la limpieza de un sitio inflamatorio, por ejemplo, sino que también facilita la presentación de los péptidos resultantes a las células del sistema inmunitario específico, como los linfocitos T o B. Esto sucede a través de la presentación en moléculas del complejo mayor de histocompatibilidad II (MHC II). De este modo, se puede iniciar una respuesta inmunitaria específica de antígeno. Además, las enzimas asociadas al fagolisosoma facilitan la producción de grandes cantidades de especies reactivas de oxígeno (ROS) que funcionan para matar de manera efectiva los patógenos ingeridos. A través de la acción de factores estimulantes, por ejemplo, en el caso de lesión tisular, se reclutan grandes cantidades de monocitos inflamatorios, precursores de macrófagos, de la médula ósea a través de gradientes de quimiocinas y diversas moléculas de adhesión. En tales situaciones fisiopatológicas, los macrófagos migratorios (reclutados) superan con creces a los macrófagos residentes en los tejidos [9, 22]. En consecuencia, los factores de crecimiento y las citocinas liberadas en el microambiente tisular local obligan a los macrófagos residentes y reclutados a proliferar y sufrir cambios fenotípicos y funcionales sustanciales [34]. Dependiendo de los estímulos microambientales y la naturaleza de la estructura del patógeno/antígeno que encuentra un macrófago, podría favorecerse la diferenciación en macrófagos M1 o M2. El perfil de diferenciación predominante determina la
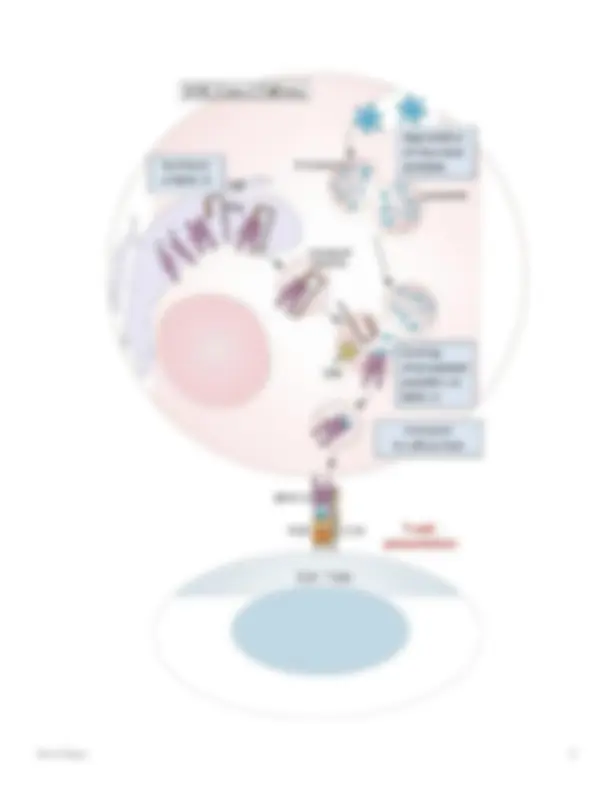
macrófagos, ya sea reclutados en respuesta a un estímulo o residentes en tejidos, atraen y activan otras células inmunitarias es la producción y liberación de factores solubles que incluyen citocinas proinflamatorias como la interleucina-1β (IL-1β) y la necrosis tumoral. factor alfa (TNFα), IL-6, IL-12, así como quimiocinas como el ligando 8 de quimiocina con motivos C-X-C (CXCL8, IL-8). Esto sucede tan pronto como los macrófagos encuentran patógenos/daño. Mientras que la IL-1β y el TNFα provocan, entre otras cosas, la activación endotelial, el aumento de la permeabilidad vascular y la activación de los linfocitos, la IL-12 conduce a la activación de las células asesinas naturales (NK), un tipo de linfocito citotóxico fundamental para el sistema inmunitario innato.. Las células NK funcionan para eliminar las células infectadas por virus y las células infectadas por otros patógenos intracelulares y las células tumorales. Los macrófagos tienen un equipo receptor típico para el reconocimiento efectivo de patógenos. Estos llamados receptores de reconocimiento de patrones (PRR), cuyos representantes más conocidos pertenecen a los receptores tipo Toll (TLR), reconocen estructuras conservadas que son características de diferentes clases de patógenos. Estas estructuras típicamente patógenas y, por lo tanto, "extrañas" también se denominan patrones moleculares asociados a patógenos (PAMPS) e incluyen, entre otros, ARN de doble cadena, ADN CpG no metilado, lipoproteínas y lipopolisacáridos (LPS) (Fig. 3).
Fig. 3
Receptores tipo Toll (TLR) y su especificidad de ligando. Diferentes TLR y su especificidad de ligando. Los TLR reconocen una variedad de estructuras asociadas a patógenos (PAMP). El reconocimiento de LPS por TLR-4 requiere las proteínas accesorias CD14 y MD-2. TLR-2 reconoce ligandos muy diferentes e interactúa con TLR-6 y TLR-1. TLR-3 está involucrado en el reconocimiento de ARN viral de doble cadena (dsRNA), TLR-5 se une a flagelina, una proteína de los flagelos bacterianos, y TLR-9 reconoce el ADN bacteriano sobre la base de sus motivos CpG no metilados ([76] con amable permiso)
Generación de ROS/RNS
La producción de especies reactivas antimicrobianas de oxígeno y nitrógeno (ROS/RNS) es un importante mecanismo microbicida de las células fagocíticas. Los defectos enzimáticos congénitos asociados con la falta o la reducción de la actividad de las enzimas formadoras de ROS/RNS se manifiestan en la destrucción incompleta de patógenos o células tumorales y los pacientes afectados sufren infecciones recurrentes o cáncer, respectivamente. Las ROS se generan intracelularmente como subproductos durante varias reacciones de transferencia de electrones, especialmente en la cadena respiratoria mitocondrial. Sin embargo, la estimulación mediada por receptores de la producción de ROS es la clave para importantes funciones efectoras de los macrófagos. Varios receptores, incluidos los receptores del complemento, los TLR y los receptores de péptidos bacterianos (p. ej., el receptor fMLF) inducen la producción de ROS/RNS en los fagolisosomas. Aquí, el ensamblaje de la NADPH oxidasa juega un papel dominante. Sus subunidades p22 y gp91, que se encuentran en la membrana celular y por lo tanto en el fagolisosoma, se ensamblan con las subunidades inicialmente ubicadas en el citosol (p40, p47 y p67) que proporcionan la NADPH oxidasa activa. La formación de la NADPH oxidasa activa conduce a un aumento en el consumo de oxígeno, lo que se denomina "explosión oxidativa". Esto crea un radical anión superóxido (O⋅−2) en la luz del fagolisosoma, que se convierte en H2O2 por la superóxido dismutasa (SOD) [59]. Otras reacciones conducen a la formación de ROS tóxicos, incluidos OH⋅ y OCl− a partir de H2O2. El repertorio antimicrobiano de los macrófagos también incluye óxidos de nitrógeno tóxicos, especialmente NO⋅. En las células que producen tanto O⋅−2 como NO⋅, como los macrófagos, la reacción casi limitada por difusión entre NO⋅ y O⋅−2 es probable que ocurra y produce
Los miembros del grupo heterólogo de citocinas se unen a diferentes tipos de receptores específicos, entre ellos receptores cinasas (p. ej., receptor de TGF-β1) y receptores ligados a tirosina cinasa que carecen de actividad cinasa intrínseca. El último grupo de receptores también se conoce como receptores de citocinas (p. ej., receptor de IL-1β y TNFα). La unión del ligando a estos receptores por una amplia variedad de citoquinas o factores de crecimiento generalmente provoca la dimerización o trimerización de las subunidades monoméricas, lo que a su vez induce la activación rápida de las quinasas asociadas con el receptor, típicamente miembros de la familia de tirosina quinasas JAK (Janus quinasa).. La fosforilación del propio receptor permite el reclutamiento y la unión a través de los dominios SH2 (homología src 2) de los miembros de la familia de factores de transcripción STAT (transductores de señales y activadores de la transcripción) al receptor. Esto permite que las JAK fosforilen STAT, que posteriormente forman dímeros que son capaces de ingresar al núcleo e iniciar específicamente la transcripción de genes objetivo, que incluyen citocinas, SOCS1–3 y factores de transcripción como GATA3, C-Maf, c-myc, NFAT1 /NFAT [30, 33].
Señalización TLR/NF-κB
La familia de factores de transcripción NF-κB juega un papel crucial en las respuestas inflamatorias y apoptóticas. Los miembros de la familia incluyen NF-κB1 p50, NF-κB2 p52, RELA (también llamado p65), RELB y c-REL1 que, como homodímeros o heterodímeros, median la transcripción de los genes diana de NF-κB [66]. Se retienen en el citoplasma por interacción con la molécula inhibidora IκB. Se pueden discriminar dos cascadas de activación de NF-κB. En la vía de activación clásica (canónica), en respuesta a varias señales, IκB se fosforila por serina proteína quinasas, principalmente IKK pero también IKK1. A continuación, IκB se degrada en el proteasoma, lo que facilita la translocación al núcleo de p50/p65 NFκB. Este heterodímero es el
complejo más abundante, a menudo denominado "NF-κB". Inicia y mejora la expresión de genes diana, incluidos iNOS, la citoquina quimioatrayente, IL-8, el TNFα inflamatorio, IL-1β e IL-6, y moléculas de adhesión (ICAM1) y quimioquinas (IP-10, MCP-1, MIP-1; Fig. 5, para más detalles ver [45]). En la cascada de activación alternativa, IKK1 media el procesamiento de p100 a p52, este último formando heterodímeros con relB que al transferirse al núcleo inician la transcripción de los genes diana de NF-κB. C-Rel es de especial importancia para, por ejemplo, la inducción transcripcional de IL- p40 en macrófagos [38].
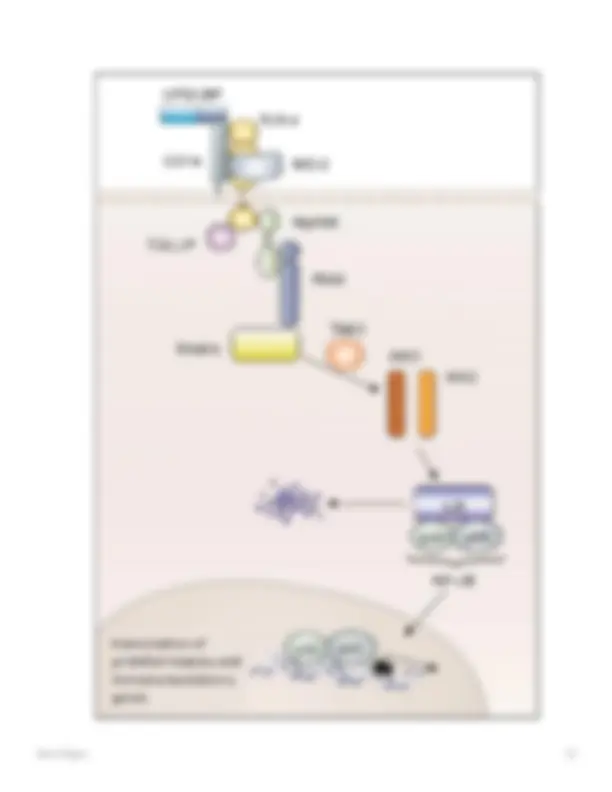
Fig. 5
Vías de transducción de señales de los receptores tipo Toll. La unión del ligando (LPS/LBP/CD14; MD-2) a TLR conduce a la asociación de moléculas adaptadoras como MyD88, Toll-interacting Protein (TOLLIP), la proteína quinasa IRAK y TRAF6 (factor asociado al receptor de TNF). 6). TRAF6 activa las quinasas 1 y 2 de IκB (IKK-1/2) a través de la quinasa TAK1 (quinasa activada por TGF-β). Estas quinasas fosforilan IκB, lo que conduce a la degradación de la proteína inhibidora y libera NF-κB como dímero. NF-κB migra al núcleo de la célula e induce una activación transcripcional de genes proinflamatorios e inmunomoduladores ([76] con permiso)
Algunos estímulos activan preferentemente la vía alternativa; otros, como la linfotoxina- β (LTβ) y el LPS, activan ambas cascadas [10, 11]. Se supone que las dos cascadas de NF-κB se activan de manera secuencial: la respuesta temprana mediada por dímeros que contienen p65, la última luego continúa una respuesta con complejos que contienen relB para mantener la activación de NF-κB [54].
El estrés oxidativo, las proteínas virales, los factores de crecimiento (angiotensina II), los mitógenos, las citoquinas inflamatorias IL-1β y TNFα a través de los receptores IL1/IL-18 o los receptores TNF, respectivamente, los receptores tipo NOD y los ligandos TLR activan IKKβ [31]. La activación de todos estos receptores también alimenta otras vías de señalización que interfieren, directa o indirectamente, con la vía de señalización de NF-κB [14]. Tras la activación por LPS, TLR4 a través de Myd88 activa NF-κB, pero además activa la proteína quinasa activada por mitógeno (MAPK), las vías IRF5 y, a través de la señalización endosomal de TLR4, se induce IRF3 y la producción de interferones antivirales potentes [14]. Los efectos autocrinos de cualquier citocina liberada por los macrófagos activados aumentan aún más la complejidad de las respuestas resultantes.
En humanos, existen al menos diez TLR diferentes que difieren en su especificidad hacia estructuras patogénicas distintas y características, como LPS, ARN de doble cadena o ADN CpG no metilado, los llamados patrones moleculares asociados a patógenos. Por esta razón, los TLR también se denominan PRR (receptores de reconocimiento de patrones). El primer TLR que se identifica es el TLR4, que es activado por LPS (bacterias gramnegativas). LPS se une a la superficie del macrófago a través de LBP y se une a TLR4 en este complejo (Fig. 4).
Señalización redox
La evidencia acumulada respalda un papel de señalización para las ROS que generan los macrófagos durante el estallido oxidativo.
En los últimos años se ha endurecido la opinión de que las ROS, al menos cuando están presentes en ciertas concentraciones, pueden verse como un segundo mensajero. En pleno acuerdo con esta función, las ROS muestran ciertas características, como la inducción transitoria dependiente del estímulo (p. ej., a través de la activación de la NADPH oxidasa), función específica (que interfiere específicamente con componentes de señalización como, p. ej., quinasas) y la rápida terminación de la señal por enzimas antioxidantes/antioxidantes (catalasa, superóxido dismutasa, glutatión peroxidasas) [18]. La modulación de la concentración intracelular de calcio [Ca2+]i es un mecanismo general involucrado aquí porque muchas vías de señalización intracelular realmente dependen de él.
Se ha demostrado que las ROS inducen la activación de NF-κB, aunque los mecanismos subyacentes aún deben dilucidarse por completo [32]. La glutatión S reversible se ha establecido como un mecanismo subyacente a la regulación de, por ejemplo, la proteína activadora del factor de transcripción 1 (AP1) [19].
Está bien establecido que las ROS producidas por los macrófagos (O⋅−2 y las especies formadas posteriormente a partir de ellas) contribuyen a la oxidación de las lipoproteínas de baja densidad (LDL). Los macrófagos absorben rápidamente el LDL oxidado, lo que lleva a su transformación en "células espumosas", que se asocian con un mayor riesgo de aterosclerosis y complicaciones relacionadas con esto. En particular, se ha demostrado que la propia LDL oxidada interfiere con muchas vías de señalización en los macrófagos [41].