Scarica Biologia cellulare: batteri e virus - Prof. Frangipani e più Appunti in PDF di Microbiologia solo su Docsity!
La microbiologia è la branca della biologia che studia la struttura e le funzioni dei microrganismi viventi, unicellulari, pluricellulari o acellulari, che non si vedono ad occhio nudo. Essa comprende lo studio dei batteri , di piccoli eucarioti come funghi, lieviti, alghe, protozoi e dei virus , parassiti intracellulari obbligati che hanno un’organizzazione non cellulare e non possiedono metabolismo proprio (possono avere genomi non a DNA). Il range nel quale l’occhio umano riesce a distinguere due corpi è da 1mm in su ; sotto questo livello si inizia a parlare di microscopia. Il microscopio ottico ha una risoluzione che permette di vedere da 1mm a 1 micrometro (Escherichia coli/staphilococco); quello elettronico permette di vedere fino ad 1 nanometro (virus dell’epatite B, poliovirus o AIDS virus) mentre per microrganismi fino a 0.1 nm (1 Armstrong) occorrono microscopi speciali. BATTERI I batteri sono presenti ovunque ( ubiquitari ); sulla terra sono 5x10^30 contro i 7x10^9 esseri umani. Essendo in numero così ampio, hanno colonizzato praticamente qualsiasi ecosistema sulla Terra. Il corpo umano è composto da mille miliardi di cellule umane e 10mila miliardi di cellule batteriche , che contribuiscono a tantissimi aspetti fondamentali della nostra fisiologia. Dal punto di vista del DNA, un uomo possiede 25mila geni umani e 2milioni e 500mila geni batterici. I microrganismi possono essere studiati :
- Osservandoli (grazie a tecnologie come il microscopio) e cercando di formulare ipotesi avvalorandole/confutandole tramite esperimenti che seguono il metodo scientifico.
- Coltivandoli in laboratorio (in terreno liquido o solido, anche se solo una piccolissima percentuale sulla totalità dei batteri presenti in natura è coltivabile in laboratorio; questo per due motivi: il primo è che l’habitat/nutrienti sono difficilmente ricreabili e così anche i loro parametri chimico/fisici, il secondo è il fatto che spesso si formano comunità batteriche complesse e quindi alcuni batteri vivono solo se sono presenti altri. Alcuni hanno anche delle dinamiche di crescita lentissime).
- Decodificando le informazioni contenute nel loro genoma (spesso il batterio non riesce ad essere coltivato in lab, ma attraverso delle tecniche si può estrarre solamente il loro acido nucleico). I batteri più noti sono i patogeni, ossia quelli che causano patologie nell’uomo o negli animali, come quelli che causano la meningite, le carie, le cistiti, la polmonite, le ulcere, le sepsi…. La cellula si divide in procariotica ed eucariotica ; le differenze risiedono nelle dimensioni, nella struttura e nella composizione. Negli E il materiale genetico è contenuto in un nucleo delimitato da membrane , nei P esso è addensato a formare il nucleoide. Il fatto che i P non hanno membrane nucleari porta ad una serie di adattamenti; se negli E l’espressione genica è compartimentata , nei P la trascrizione e la traduzione avvengono quasi simultaneamente ; la traduzione spesso inizia senza che la trascrizione sia finita. Nelle cellule dei P no n ci sono i mitocondri , poiché la respirazione avviene sulla membrana citoplasmatica e non ci sono i cloroplasti. Alcuni batteri però, hanno avuto bisogno di una compartimentalizzazione, ma sono delle eccezioni e questi
comparti sono diversi da quelli degli E. All’interno della famiglia dei batteri, vi è il domino degli Archea, la quale membrana non è un doppio strato fosfolipidico, ma è formata da polimeri di isoprene. Le cellule batteriche svolgono tutte le funzioni che svolgono quelle eucariotiche, ma in modo diverso; tutte sono capaci di:
- Assumere nutrienti dall’esterno, modificarli ed espellere rifiuti ( metabolismo , che può essere genetico, replicazione, trascrizione e traduzione, o catalitico, energia e biosintesi).
- Crescere , ossia di aumentare di numero o di biomassa tramite la traduzione dei nutrienti assunti dall’ambiente.
- Evolversi , ossia acquisire nel tempo nuove proprietà. Alcune cellule possiedono delle proprietà, come:
- La capacità di differenziarsi , formando nuove strutture cellulari come endospore, altamente resistenti a condizioni ambientali avverse, grazie alle quali il materiale genetico viene protetto affinchè l’ambiente non ritorna vivibile.
- La capacità di comunicare , ossia la capacità di comunicare tra loro producendo molecole liberate nell’ambiente , che vengono captate da altre cellule; ciò permette di coordinare delle funzioni, come l’espressione genica.
- La capacità di scambiarsi materiale genetico tramite il trasferimento di plasmidi, di virus batterici o DNA nudo.
- La capacità di spostarsi autonomamente , grazie ad appendici come flagelli, ciglia e pili , di cui sono dotate. CLASSIFICAZIONE DEGLI ORGANISMI VIVENTI 1 mar. 22 FORMA DELLE CELLULE BATTERICHE Ogni genere di cellula batterica ha una forma peculiare; queste vengono raggruppate in:
- Batteri coccoidi : forma rotondeggiante, possono vivere come cellule singole, come diplococchi (due cocchi), micrococchi (es. streptococco).
- Batteri a bastoncello : forma allungata a bacillo, come i micobatteri della tubercolosi; alcuni hanno
altro che provò questa teoria fu Spallanzani (metà ‘700), conducendo i suoi esperimenti sul brodo , esponendolo all’aria. Si osservava la formazione di qualcosa, anche mettendoci il tappo. Poi uccise con il calore quelle forme di vita e chiuse subito il tappo e la crescita non si verificò, mentre non chiudendolo essa si verificava. Il padre della microbiologia moderna fu Louis Pasteur (1822-95), che studiava le anomalie della fermentazione della birra, del vino e dell’aceto. Mise a punto la pastorizzazione, cioè un metodo di riduzione della carica microbica di alimenti che non possono essere sottoposti a fonti di calore troppo alte, scaldando questi alimenti a temperature basse per poco tempo. Studiò la rabbia, mettendo a punto un vaccino antirabbica e fu il padre della sieroterapia. Anche Pasteur per dimostrare la presenza dei microrganismi conduce esperimenti: versa un liquido in una fiasca non sterile, stirandone il collo con una fiamma; il liquido viene sterilizzato e il vapore viene espulso dall’estremità aperta. Da qui, il liquido viene fatto raffreddare lentamente e polvere e microrganismi restano intrappolati nel tratto curvo della fiasca facendo restare il liquido sterile. Nel lungo periodo, la fiasca viene inclinata così da portare la polvere all’interno del liquido, dove si svilupperanno microrganismi e di conseguenza non sarà sterile. Il padre della microbiologia clinica fu Koch e fu il primo che associò una sintomatologia clinica alla presenza di un microrganismo ; egli mise anche a punto la tecnica di coltivazione su piastra (a terreno solido), mentre l’utilizzo dell’agar-agar venne in mente a Fannie Hesse, moglie di un allievo di Koch. Koch studiava il colera, il carbonchio e la tubercolosi andando a studiare le componenti del sangue degli animali malati. Egli notò che vi erano delle cellule piccole sempre nell’animale malato e mai in quello sano, quindi, ipotizzò che fosse questa la ragione della patologia. Strisciando il sangue su una piastra di agar, si svilupparono delle colonie e le inoculò in un animale sano, per vedere se l’animale sviluppava la malattia; questo la sviluppava e moriva. Poi prelevò del sangue o un campione di tessuto a questo animale morto sviluppano una coltura per confrontare se ciò che ne derivava era uguale a ciò che aveva isolato in precedenza. Ancora oggi, affinché una malattia possa essere considerata infettiva, deve seguire i 4 postulati di Koch (con piccole modifiche, poiché ora sono stati “scoperti” i portatori sani). I padri della microbiologia ambientale furono Beijerinck e Vinogradskij, che si occuparono delle tecniche delle “culture di arricchimento” e del virus del mosaico del tabacco (Beijerinck padre della virologia) e Vinogradskij di autotrofia (capacità di un organismo di sintetizzare le proprie molecole organiche a partire da sostanze inorganiche) e di chemiolitotrofia (capacità metabolica di ricavare energia ossidando composti organici e non). Dalla metà del ‘900 inizia l’era della microbiologia molecolare e nasce la genetica dei microrganismi, che spiega processi legati alla trasmissibilità del DNA, i processi ecc. Nel 1953 si è scoperta la struttura del DNA e nel ’66 il dogma della biologia (DNA-RNA-proteine). Nel 1977 per la prima volta si è sequenziato il DNA e vennero scoperti gli Archea. La PCR , grazie a dei microrganismi che riuscivano a polimerizzare il DNA a temperature elevatissime, è stata scoperta nel 1985 e da lì iniziò l’era della microbiologia molecolare, genomica e proteomica , che hanno permesso si sequenziare i genomi, compreso quello umano.
TAPPE FONDAMENTALI
STRUTTURA DELLA CELLULA
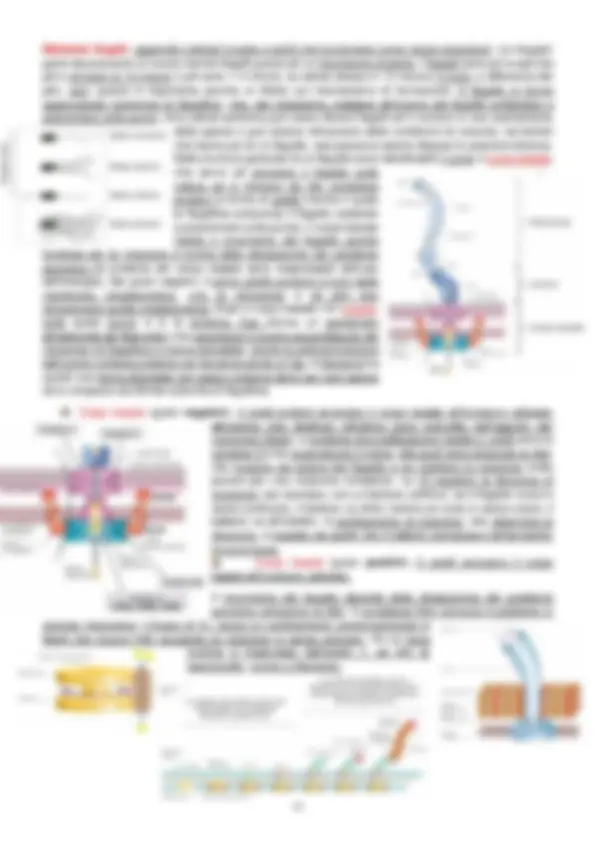
Nella cellula procariotica non c’è una membrana a delimitare l’acido nucleico, non ci sono organuli come mitocondri né strutture membranose come il RE o l’apparato del Golgi. Trascrizione e traduzione sono quasi fenomeni simultanei poiché non vi è la compartimentazione in organelli/posizioni specifiche. Le cellule procariotiche sono talmente piccole che si riescono a vedere solo con il microscopio ottico. Per vederle, le cellule devono essere colorate ; alcune sono colorate naturalmente, per altre bisogna ricorrere ad un certo tipo di colorazioni, come quelle a fluorescenza o quella di Gram. La colorazione di Gram (1884, medico danese Hans Gram) viene utilizzata tutt’oggi ed è differenziale , ossia colora in maniera differente due tipologie di batteri che a seguito della colorazione vengono suddivisi in gram positivi (blu) e gram negativi (rosso). Questa colorazione è possibile perche le cellule batteriche possono essere di due tipologie: la parete che le riveste nei gram negativi è costituita da un una
Gli acidi grassi sono legati al glicerolo tramite un legame estere. La composizione lipidica della membrana varia a sceonda della temperatura, in modo da mantenere più o meno costante la fluidità di membrana. Se la catena contiene acidi grassi saturi, la fluidità è minore e la temperatura di fluidificazione è più alta; se la catena contiene acidi grassi insaturi, la fluidità aumenta e la temperatura di fluidificazione è minore. Nella membrana procariotica no n c’è il colesterolo ma ci sono molecole che svolgono una funzione simile; il colesterolo ha una struttura planare che si intercala negli acidi grassi rendendo la membrana meno flessibile. Nei batteri a svolgere questa funzione ci pensano gli opanoidi, simili agli steroli sia strutturalmente che funzionalmente (Archea no opanoidi). La membrana degli Archea sono presenti catene isopreniche ; tra il glicerolo e l’acidi grassi vi è un legame etere poiché manca un carbonile. Il glicerolo negli Archea è L, mentre nei batteri è D. Anche negli Archea è possibile individuare una parte polare e una apolare ma non tutti hanno la membrana formata da un doppio strato. Alcuni hanno la membrana a monostrato, in cui la catena isoprenica è più lunga e possiede due teste polari; ciò influisce sulla resistenza. Il monostrato è più resistente del doppio strato poiché è più rigido e non si interrompe a metà. Negli Archea no n ci sono nemmeno gli opanoidi ma vi è la necessità di più stabilità; le catene isopreniche hanno la capacità di ciclizzare e formare parti planari che vanno a svolgere la stessa funzione del colesterolo e degli opanoidi. Queste strutture a monostrato le si trovano spesso in Archea ipertermofili che hanno bisogno di più resistenza. Nella membrana citoplasmatica vi sono proteine periferiche, associate alla membrana tramite legami deboli, o integrali, che possono essere canali, trasportatori o sensori. Le proteine che attraversano la membrana hanno natura anfipatica ; le regioni idrofobiche affondano nel doppio strato lipidico mentre le parti idrofile sporgono sui due lati della membrana plasmatica. Le funzioni di quest’ultima sono quella di barriera selettivamente permeabile , conservazione (riesce a conservare un gradiente protonico perché la concentrazione di ioni H+ è più elevata all’esterno che all’esterno) e produzione di energia (avviene la respirazione cellulare mentre negli eucarioti avviene nei mitocondri) e fa da sito di ancoraggi per proteine essenziali. Ci sono dei problemi però; la membrana è molto poco permeabile alla maggior parte delle molecole, come gli ioni (la bassissima permeabilità permette al batterio di generare un gradiente), di cui la cellula ha molto bisogno quindi necessariamente esisteranno sistemi che internalizzano queste molecole contro gradiente, ossia meccanismi di trasporto transmembrana, che utilizzano energia. 7 mar. ’ La membrana è quindi una barriera selettivamente permeabile , cioè è permeabile a particolari tipi di soluti, come acqua, gas, piccole molecole liposolubili prive di carica, mentre non lo è per ioni, composti polari e grosse molecole. La cellula, però, ha bisogno anche di questi ultimi quindi attiva dei sistemi in grado di trasportarli dall’esterno (dove sono maggiormente concentrati) all’interno della cellula. Questi sistemi prendono il nome di sistemi di trasporto e si dividono in trasporto passivo (diffusione semplice e
facilitata, avvengono senza dispendio di energia e secondo gradiente) e attivo (diretto o indiretto, avviene con dispendio di energia e contro gradiente). La diffusione semplice avviene secondo due modalità:
- Gradiente di concentrazione , quando il soluto è non carico e si muove da dove è più concentrato a dove lo è meno per raggiungere l’equilibrio. Nel caso la membrana non fosse permeabile al soluto, si osserva il fenomeno dell’ osmosi , che consiste nel trasferimento di molecole d’acqua da dove il soluto è meno concentrato a dove lo è di più per diluire la soluzione ( pressione osmotica invece è la pressione da applicare per impedire il passaggio di acqua)
- Gradiente elettrochimico , quando il soluto è carico , ed è la forza che si origina dal potenziale elettrico , ossia la differenza di cariche ai lati della membrana. 2.La diffusione facilitata avviene tramite l’ausilio di proteine canale o di trasporto. La proteina di trasporto ha un sito di legame specifico per il soluto da trasportare e alterna stati conformazionali : come prima cosa il soluto si lega alla proteina, che di conseguenza subisce una modificazione; rilascia il soluto e ritorna alla sua forma originaria pronta a legare altro soluto. Più il soluto è concentrato, più la diffusione è veloce, fino a quando tutti i trasportatori non sono saturati (la velocità diventa costante). Ci sono diversi tipi di trasporto: Uniporto , trasporto di una sola sostanza secondo gradiente. Simporto , trasporto che usa il flusso di un soluto secondo gradiente per muovere, nella stessa direzione, una molecola contro gradiente. Antiporto , trasporto che usa il flusso di un soluto secondo gradiente per muovere, in direzione opposta, una molecola contro gradiente. La proteina canale permette il passaggio di ioni , che non passerebbero perché sono carichi e hanno un ingombro notevole. Le proteine canale hanno eliche idrofobe che aprono il canale e parti idrofile a contatto con l’ambiente acquoso ; il movimento avviene sia tramite gradiente di concentrazione sia elettrochimico. L’apertura del canale e controllata dal voltaggio (variazione di carica) dal ligando (che si lega al sito bersaglio della proteina) e da uno stimolo meccanico. Altri trasportatori sono le porine , meno selettive che aprono un canale più ampio interagendo con il doppio strato lipidico. I trasportatori possono essere più o meno specifici (singolo substrato o una classe di substrati) e possono essere regolati nella loro espressione o nella loro attività; per esempio, se un batterio si trova in un ambiente povero di nutrienti, userà più trasportatori per essere sicuro di accumularne il massimo possibile. Un esempio di trasportatori sono quelli di tipo ABC (ATP binding cassette), solitamente utilizzati per il trasporto di amminoacidi rari come l’istidina. Questi sono presenti nel periplasma, che però non è energizzato, e affinché la proteina periplasmatica di legame possa intercettarli e legarli, e poi farli passare attraverso il canale che li porta nel citoplasma, vi è bisogno di energia, che è fornita dall’idrolisi dell’ATP presente nel citoplasma. 3.Il trasporto attivo è un movimento endoergonico di ioni e altri soluti anche contro gradiente ed è accoppiato ad un processo esoergonico ; è unidirezionale , poiché i trasportatori funzionano per determinati versi e contribuisce a mantenere l’ambiente interno costante e distante dall’equilibrio. Può essere:
- Diretto , come la pompa sodio-potassio che è associata direttamente all’idrolisi dell’ATP e mantiene elevati i livelli intracellulari di potassio e bassi quelli di sodio, tramite il loro spostamento contro gradiente.
- Indiretto : trasporto di zuccheri, amminoacidi e altre molecole accoppiato con il passaggio di ioni sodio o protoni secondo i loro gradienti elettrochimici.
- Traslocazione di gruppo : coinvolge più proteine e durante il processo il substrato da trasportare viene modificato chimicamente; per esempio, il glucosio viene trasformato in glucosio- 6 - fosfato a causa della fosforilazione a opera della fosfotransferasi.
Invece di un tetrapeptide, abbiamo un pentapeptide poiché dopo il DAP viene aggiunto un dimero di D-alanina, non un monomero (UDP-NAM + pentapeptide). Il pentapeptide si associa al NAG e attraversa la membrana plasmatica.
- II tappa: trasporto del glican-pentapeptide attraverso la membrana. Il glican-pentapeptide riesce ad attraversare la membrana grazie al bactoprenolo trifosfato , una molecola lipidica fortemente idrofoba. Il NAM- pentapeptide viene caricato sul bactoprenolo; al complesso si aggancia anche il NAG e viene formato il monomero (associato al bactoprenolo tri-fosfato). Quest’ultimo riesce ad attraversa re la membrana ed esporre all’esterno (periplasma) il monomero (si flippa). Il monomero adesso deve essere inserito nel peptidoglicano già presente per accrescere la parete (durante l’accrescimento della cellula prima della divisione).
- III tappa: polimerizzazione dei monomeri. Dopo che il monomero si inserisce, il primo legame che si forma è quello di tipo glicosidico (tra NAM e NAG) grazie alle transglicosilasi , poi avverrano i legami di transpeptidazione che coinvolgono la parte amminoacidica, catalizzati dalla transpeptidasi (PBP, penicillin-binding-proteins; la penicillina vi si lega bloccandone l’attività e bloccando quindi la sintesi della parete). Una volta che il monomero si trova nel periplasma, tutte le reazioni che avvengono lì hanno bisogno di energia (che non sia derivata dall’idrolisi ATP, che si trova nel citoplasma) e quindi usano l’energia insita nel pentapeptide , idrolizzando e scindendo il legame (a opera delle transpeptidasi) che tiene unito il dimero di D-alanina per formare il legame crociato DAP/D-alanina. La sintesi del peptidoglicano deve essere accoppiata alla presenza di enzimi con funzioni litiche , che vanno a idrolizzare legami tra i monomeri del peptidoglicano gia presente al fine di poterne inserire di nuovi. L’attività litica deve essere regolata e coordinata con l’accrescimento della parete in siti specifici, degradando il filamento di glicano e sostituendolo con 3 filamenti nuovi. Gli enzimi con funzioni litiche sono le autolisine, e ce ne sono di diversi tipi: le muramidasi e le transglicosilasi litiche (scindono il legame tra NAM e NAG), le amidasi (NAM e L-alanina), le endopeptidasi (aa-aa) e le carbossipeptidasi (X e D- alanina). GRAM POSITIVI vs GRAM NEGATIVI Nei gram positivi il peptidoglicano costituisce il 40% del peso secco della cellula, ma non solo. Sono presenti proteine associate alla parete, acido teicoico e acido lipoteicoico inseriti nel peptidoglicano. L’acido lipoteicoico è più grande e ha affinità con la parte lipidica andando a inserirsi nella membrana citoplasmatica. Questi acidi sono polimeri anionici costituiti da unità ripetute di 1,3-glicerol-fosfato o di 1,5 D-ribitol-fosfato legati tra loro da legami fosfodiesterici, che si legano al NAM del peptidoglicano covalentemente. Gli acidi teicoici determinano le caratterische chimico-fisiche della parete (porosità, elasticità, resistenza…), sono responsabili della carica negativa della superficie cellulare e controllano l’assimilazione dei cationi metallici, sono riserva di fosfato e sono importanti ai fini dell’attaccamento alla mucosa e allo smalto dentale.
Nei gram negativi sono presenti due membrane, esterna e interna (citoplasmatica); tra le due vi è il periplasma, che contiene il peptidoglicano. La membrana esterna è asimmetrica; la faccia rivolta verso il periplasma (fosfolipidi) è fatta in modo diverso di quella rivolta verso l’esterno (LPS=lipopolisaccaride). Troviamo sempre le porine, che attraversano la membrana esterna (la loro discriminazione avviene in base alla dimensione, non alla natura del substrato). L’LPS è costituito da lipide A , il core (esterno e interno) e l’antigene O. Il lipide A prende contatto coi fosfolipidi della faccia periplasmatica, la parte core è costante in tutti gli LPS mentre la parte variabile consiste negli zuccheri che vanno a formare l’antigene O. Il lipide A è formato da un numero variabile di acidi grassi legati ad un dimero di NAG esterificato con due gruppi fosfato (in modo da avere una parte polare). Questa parte poi lega il core polisaccaridico, che è una regione relativamente conservata formata da varie molecole di zuccheri (esosi, eptosi…) che poi legano l’antigene O, regione polisaccaridica molto variabile costituita da unità ripetute, fortemente immunogenica. Sulla membrana esterna possono inoltre esserci proteine come recettori specifici e lipoproteine (strutturali, di trasporto, di trasduzione del segnale). Un esempio è la proteina di Braun , che ancora membrana esterna al peptidoglicano. PERIPLASMA Il periplasma è lo spazio racchiuso tra membrana esterna e interna ed è costituito da una matrice simile ad un gel che contiene peptidoglicano e proteine, come le proteine trasportatori di aa come ABC. Esso è in continuità con lo spazio extracellulare tramite le porine e non contiene ATP o altre forme di energia equivalenti; infatti, i processi endoergonici devono accoppiarsi a sistemi della membrana citoplasmatica in grado di portare energia fino al periplasma. Le porine sono degli omotrimeri e ogni monomero forma un canale attraverso cui passano le molecole; alcune sono aspecifiche, e la specificità spesso è data dalle dimensioni del substrato, che passa secondo gradiente. COLORAZIONE DI GRAM Questa colorazione sfrutta le caratteristiche strutturali tipiche dei gram positivi (violetto) e negativi (rosso).
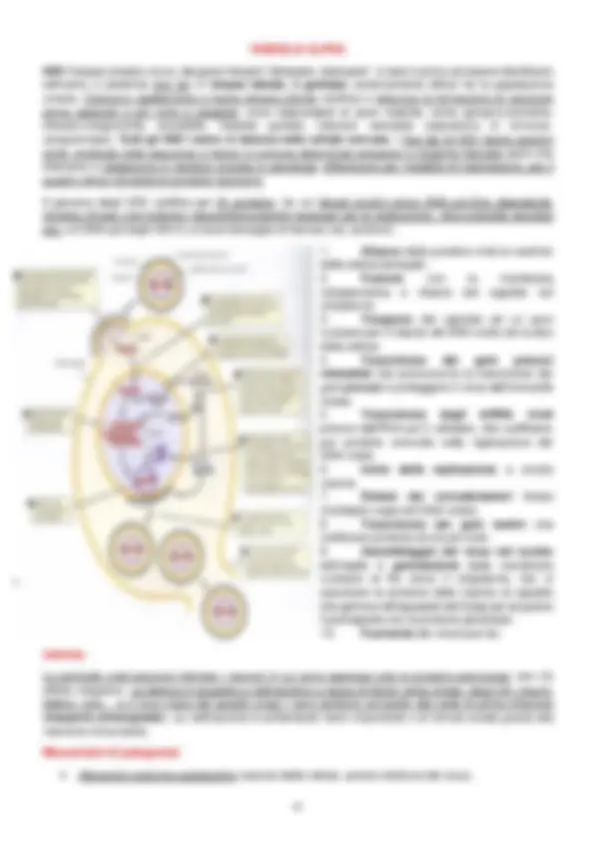
Glicogeno e amido sono polimeri del glucosio che costituiscono una fonte di riserva di carbonio e di energia e vengono prodotti quanod i batteri si trovano in periodi di limitata disponibilità di azoto con carbonio organico in eccesso. I poli-β-idrossialcanoati (PHA) sono presenti sia nei batteri che negli Archaea; i più diffusi sono i granuli di poli-beta-idrossibutirrato, costituito da monomeri di acido idrossibutirrico legati da un legame estere. Il gruppo R è molto variabile e servono da riserva di carbonio e di energia. La cianoficina è presente unicamente nei cianobatteri, capaci di fissare la CO 2 atmosferica trasformando carbonio organico in inorganico (fotosintesi). I granuli di cianoficina sono formati da un copolimero di acido aspartico e arginina e vengono utilizzati come riserva sia di energia, sia di azoto. Alcuni cianobatteri producono il benstonite, un minerale contenente bario, stronzio, magnesio che hanno la probabile funzione di appesantire la cellula per farle vivere nei loro habitat naturali, che possono essere le profondità di un mare ecc. I polifosfati sono polimeri di ortofosfati (centinaia) e servono da riserva di fosfato e energia. Altre riserve fondamentali sono quelle di zolfo; alcuni batteri ricavano energia dall’ossidazione dello zolfo, i granuli del quale vengono accumulati nel periplasma. Molti batteri possiedono “ORGANELLI” intracellulari , ossia strutture che permettono lo svolgimento di particolari funzioni. Questi possono essere:
- Magnetosomi : cristalli di magnetite o grigite circondati da un monostrato fosfolipidico. I magnetosomi si formano dall’invaginazione di un singolo strato della membrana citoplasmatica che ingloba i cristalli; questi permettono di orientare la cellula batterica rispetto ai campi magnetici terrestri e servono per la navigazione geomagnetica negli habitat acquatici.
- Clorosomi.
- Vescicole gassose : proteine che vanno a costituire una vescicola permeabile ai gas ma impermeabile all’acqua e permette, alle cellule che vivono negli ambienti acquatici, di galleggiare. I gas entrano nella vescicola, diminuendo la densità e facendole galleggiare; quando i gas vengono rilasciati, la densità aumenta e la vescicola “sprofonda”. La vescicola ha una struttura fusiforme e rivestimento proteico, dato da due proteine: GvpA (piccola e idrofobica, beta foglietti e costituisce la superficie dell’involucro) e GvpC (lega fra di loro i foglietti beta assumento una struttura ad alfa- elica).
- Microcompartimenti cellulari : strutture subcellulari con involucro di natura proteica, che contengono enzimi impegnati in vie metaboliche particolari. Durante la conversione di un substrato A nel suo prodotto B, è possibile che si formino degli intermedi volatili che si allontanerebbero dal sito in cui devono stare se non fossero trattenuti, oppure questi intermedi possono essere tossici e vi è la necessità di isolarli. SISTEMI DI SECREZIONE I sistemi di secrezione sono dei sistemi che permettono alla cellula di esportare proteine (e altre molecole) che vanno a costituire gli involucri cellulari, canali, la membrana citoplasmatica o quelle necessarie nel mezzo extracellulare. Le molecole sono sintetizzate nel citoplasma e affinchè possano raggiungere la loro posizione finale è necessario che siano presenti sistemi di trasporto e di secrezione specifici. La cellula riesce a riconoscere le proteine grazie ad un peptide segnale presente alla fine della sequenza amminoacidica. Le proteine secrete attraverso il sistema Sec (secretion) possiedono una sequenza segnale all’estremità dell’N-terminale (in media 20/25 amminoacidi che hanno una struttura conservata); queste andranno poi verso un apparato che fa da “porta d’uscita” dalla membrana. Non tutte devono uscire dalla cellula; alcune possiedono dei segnali che chiariscono se queste debbano o no uscire dalla membrana. La sequenza segnale , a seconda della catena laterale degli amminoacidi che la compongono, può avere una natura polare, apolare, acida, basica : i primi amminoacidi che la costituiscono avranno una natura idrofila basica, quelli che costituiscono la regione centrale avranno natura idrofobica mentre avranno una regione carbossiterminale moderatamente polare. Il peptide segnale al carbossiterminale possiede un sito di taglio riconosciuto dalle peptidasi (signal peptidase), che tagliano la sequenza segnale
una volta che la proteina esce dalla membrana. Le proteine possono avere delle strutture secondarie: ad alfa elica e a foglietto beta. L’alfa elica è una struttura elicoidale e rappresenta la più semplice disposizione che una catena polipeptidica può assumere; in questa struttura l’amminico di ogni legame peptidico è legato al carbonile di un altro localizzato a 4 aa di distanza nella stessa catena (per stabilizzare la struttura). Il foglietto beta è una struttura planare molto compatta a pieghe dove più sequenze si dispongono una accanto all’altra e possono avere andamento parallelo o non parallele; tra amminoacidi adiacenti si formato legami a idrogeno che stabilizzano la struttura.
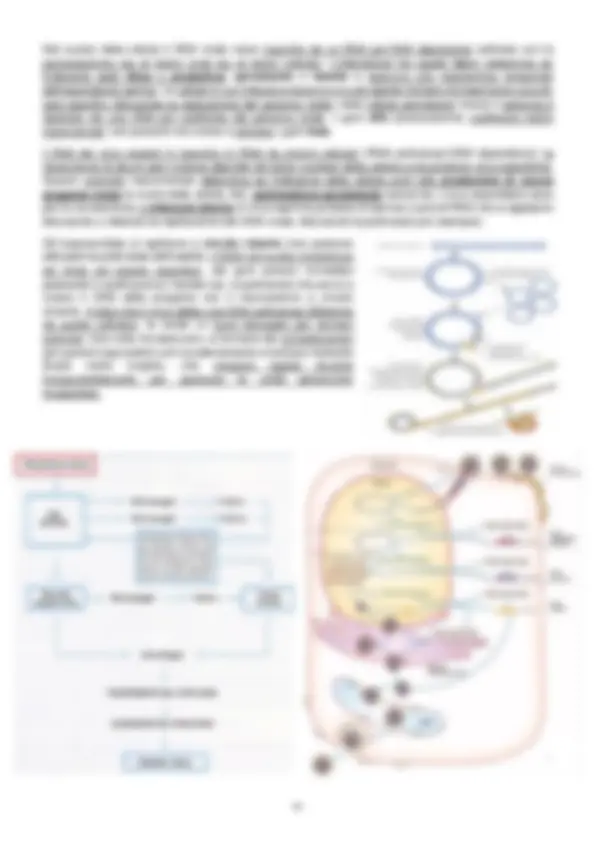
- Come vengono indirizzate le proteine attraverso la membrana? Attraverso il sistema Sec. Nella figura troviamo un mRNA con il ribosoma che sta svolgendo la traduzione, creando il polipeptide. La parte gialla è l’inizio (N-terminale, peptide segnale) mentre quella fucsia è la fine con l’estremità carbossiterminale. La presenza del segnale fa sì che le proteine secrete attraverso il Sec vengano legate attraverso il citoplasma dallo chaperone citoplasmatico SecB, che accompagna la proteina nel suo “non ripiegamento” poiché devono attraversare la membrana e non lo farebbero se fossero ripiegate (potrebbero nascondere il peptide segnale non permettendo al traslocone di legarsi). Per attraversare la membrana, la proteina viene indirizzata verso il SecA e passa attraverso un sistema proteico detto traslocone (SecYEG). Questo è un processo endoergonico, quindi serve energia che deriva dall’idrolisi dell’ATP; siccome l’energia metabolica è nel citoplasma, è presente l’ATPasi ( SecA ) che indirizza l’energia verso la membrana. Dopo che la proteina è uscita dal canale, la peptidasi taglia la sequenza segnale. Questo tipo di trasporto è post-traduzionale (la proteina viene prodotta interamente e poi trasportata) mentre ce ne sono altri in cui il trasporto è co-traduzionale (mentre la proteina viene tradotta, viene trasportata verso la sua destinazione finale). Chaperone molecolari : sono proteine che sono coivolte in varie funzioni fondamentali, come il ripiegamento delle proteine (corretto stato conformazionale delle proteine appena sintetizzate), l’assemblaggio di complessi multiproteici, la rinaturazione e/o degradazione di proteine danneggiate (alcune proteine si aggregano in corpi di diffusione, ossia complessi insolubili, e per renderle solubili si usano chaperon) o il mantenimento di uno stato parziale di denaturazione delle proteine (chaperon SecB, citoplasmatico). Nei batteri come i gram negativi questi chaperon sono presenti sia nel citoplasma che nel periplasma. Dal periplasma, le proteine possono rimanerci, essere trasportate all’esterno o essere inserite nella membrana citoplasmatica. Ma come avviene l’inserzione delle proteine nella membrana? Tramite il sistema SRP (signal recognition particle) costituito dalla ribonucleoproteina SRP (composta da proteina Ffh e un piccolo RNA 4.5S), il recettore di membrana del complesso SRP ( FtsY ), il ribosoma, il recettore per il ribosoma e il traslocone sec, che si associa ad una proteina della membrana YidC , il quale permette il corretto ripiegamento. L’SRP riconosce le proteine che vanno inserite nella membrana prima che queste vengano completamente tradotte grazie alla sequenza di riconoscimento, ossia una regione idrofobica all’amminoterminale (generalmente corrisponde alla prima elica transmembrana) che viene esposta all’esterno del ribosoma. Una volta che la ribonucleoproteina lega la sequenza di riconoscimento, la guida verso la membrana, sulla quale c’è un recettore ( FtsY ) che la accoglie e ne media l’inserimento; a questo punto il complesso si dissocia. Il recettore FtsY interagisce con un complesso multiproteico che aiuta l’integrazione della proteina nella membrana e che è formato dal traslocone Sec e la YidC. Questo tipo di trasporto è co-traduzionale.
Sistema TAT: il sistema TAT (twin-arginine translocation) riconosce come proteina da esportare delle proteine che hanno all’amminoterminale due arginine vicine. In Escherichia coli, il sistema TAT è composto da 3 proteine: la TatA , che forma una struttura multimerica che costituisce il canale, la TatB e la TatC coinvolte nel riconoscimento della sequenza segnale. Le proteine che vengono trasportate sono gia nella loro conformazione finale (struttura terziaria/quaternaria) e possono spesso coinvolgere cofattori, oppure TAT può trasportare interi complessi proteici. Questo tipo di trasporto è ovviamente post-traduzionale e richiede energia, che deriva dalla PMF (forza proton-motrice, gradiente si forma a cavallo della membrana citoplasmatica poiché all’interno vi è una concentrazione minore di H+ che entreranno secondo gradiente ). Solitamente le proteine trasportate con TAT rimangono nel citoplasma. Sistemi TAT indipendenti one-step: Sistema di secrezione di tipo I : nei gram negativi è costituito da tre componenti: outer membrane protein, membrane fusion protein e trasportatore ABC; queste tre proteine formano un canale unico che attraversa le due membrane e il trasportatore ABC e la membrane fusion proteine sono specifici per il substrato. L’energia che serve deriva dall’idrolisi di ATP. La proteina secreta ha la sequenza di riconoscimento al carbossiterminale e non viene tagliata. Sistema di secrezione di tipo III (iniettosoma): sistemi presenti in batteri patogeni che permettono l’iniezione di proteine effettrici (fattori di virulenza) direttamente nel citoplasma delle cellule eucariotiche bersaglio. Questo sistema è formato da numerose proteine che attraversano la membrana citoplasmatica e posseggono un’estremità detta ago , che a sua volta possiede un “ tappo ” il quale, una volta oltrepassata la membrana esterna, si apre e fa si che l’ago si infili nella cellula bersaglio; la secrezione è attivata dal contatto con la cellula bersaglio. Le proteine che costituiscono il sistema sono numerose e sono conservate mentre le altre (ago, effettrici…) sono specifiche. L’energia deriva dall’idrolisi dell’ATP e le proteine trasportate hanno segnale di riconoscimento che sembra trovarsi all’amminoterminale e sono presenti degli chaperon che mantengono le proteine nello stato non ripiegato finchè la proteina non arriva alla cellula bersaglio. Sistema di secrezione di tipo IV: assomiglia ad un pilo coniugativo e media la traslocazione di numerose molecole di natura differente ed è costituito da almeno 12 proteine e una struttura simile ad un pilo. BIOGENESI DELLA MEMBRANA ESTERNA (gram negativi): Tutti i precursori (proteine integrali della OM, lipoproteine, LPS…) sono sintetizzati nel citoplasma e devono essere trasportati verso il periplasma o la membrana esterna, che però non sono energizzati quindi l’energia deve trovare un modo per arrivare, dal citoplasma, alle altre destinazioni. Tutte le OMP studiate attraversano la membrana citoplasmatica mediante Sec o TAT per cui raggiungono il periplasma
unfolded e in questa condizione possono essere legate ad un chaperon permettendo il contatto di queste proteine con il SurA, che ne permette il ripiegamento, dopo che esse sono entrate in contatto con il sistema Bam; in E.coli , l’inserzione nella OM avviene attraverso un complesso della membrana esterna detto Bam (beta-barrel assembly machinery). Il Bam è formato da varie proteine, come la BamA (molto conservata) costituita da 12 foglietti beta e da una lunga regione amminoterminale solubile che protrude nello spazio periplasmatico. Questa regione riconosce sequenze specifiche al carbossiterminale di molte OMP. Il sistema Bam funziona grazie alla presenza di lipoproteine (proteine
- acidi grassi) che attraversano la IM tramite Sec e l’aggiunta della regione lipidica avviene dopo il loro trasporto attraverso la membrana citoplasmatica. Le lipoproteine vengono trasportate fino alla OM grazie ad un complesso proteico detto “ Lol ” che le riconosce e ne decide la localizzazione finale. Siccome i lipidi fanno fatica ad attraversare il periplasma (che è un mezzo acquoso), LolA li lega e porta la lipoproteina a LolB che la inserisce nella OM. Se la proteina ha l’acido aspartico in posizione +2 della lipoproteina allora questa resterà nella IM mentre se non lo possiede lì andrà nella OM. La sintesi di LPS avviene in due parti: prima l’antigene O e poi la porzione lipide A-core polisaccaridico, che poi vengono separatamente trasportati attraverso la IM. Entrambi sono sintetizzati nel citoplasma; i monomeri dell’antigene O attraversano la membrana tramite bactoprenolo (che era anche il trasportatore del peptidoglicano) e viene assemblato sul bactoprenolo sul lato periplasmatico della IM. La porzione lipide A-core polisaccaridico attraversa la membrana tramite un trasportatore ABC detto MsbA. L’LPS maturo viene generato nel periplasma grazie alla polimerasi Mzy e la ligasi WaaL. Una volta assemblato, il complesso multiproteico Lpt lo trasporta verso la OM. DIFFERENZIAMENTO CELLULARE Il differenziamento avviene quando la cellula batterica si differenzia dalla sua cellula madre ed il fenomeno è spesso correlato a fattori biologici e di sopravvivenza o a stimoli provenienti dall’ambiente. Il primo tipo di differenziamento cellulare è la capacità di formare endospore , che sono molto resistenti al calore e non possoo essere distrutte facilmente. Si formano durante un processo detto “sporulazione” e la sua funzione è quella di permettere al microrganismo di resistere anche in condizioni avverse (disidratazione, esaurimento di nutrienti); le spore possono germinare e dar vita a nuove cellule vegetative quando le condizioni ambientali diventano più favorevoli (i generi bacillus e clostridium tra i batteri - gram positivi- sporigeni). Alla fine del processo di sporulazione, da una cellula madre matura un’endospora matura che si libera grazie alla lisi della cellula madre (nel ciclo vegetativo, da una madre si originano due cellule figlie).
Quando e perché si formano le spore? Condizioni avverse agiscono come stimolo per innescare il processo di formazione della spora; per esempio, nel Bacillus subtilis, le condizioni avverse sono la carenza di nutrienti (carbonio, azoto, fosfati), elevata densità cellulare e presenza di sostanze tossiche. In vitro (beuta + terreno di crescita), si ha una crescità esponenziale fino a raggiungere la saturazione (finiscono i nutrienti, qua glucosio) e inizia la sporulazione. Le endospore resistono ad alte temperature (es. spore di clostridium botulinum resistono ai 100°C per 330 min), a radiazione UV, a disinfettanti e ad invecchiamento (possono rimanere quiescenti per anni). Studiare la sporulazione ha permesso di studiare la regolazione genica dei batteri e meccanismi che si possono trovare in batteri ma anche in altri organismi. A livello molecolare, tutto ciò che avviene durante la sporulazione avviene grazie all’espressione genica differenziale temporalmente coordinata tra la cellula madre e l’endospora. La sporulazione è il fenomeno meglio studiato di differenziamento dei microroganismi; la sua caratteristica principale è data da una organizzazione ed una coordinazione spaziale e temporale dell’espressione (trascizione e traduzione) dei geni coinvolti in ciascuna fase. Tutto il processo si basa sull’espressione differenziale dei fattori sigma, che diversificano il punto di attacco della RNA polimerasi. Quest’ultima esiste quindi in due forme: una senza e una con il fattore sigma. Quando il fattore c’è, la polimerasi si lega al gene e trascrive. I fattori sigma sono differenti a seconda del gene che devono trascrivere; i geni che servono in risposta ad elevate temperature, sono presenti solo quando le temperature sono alte, in modo da utilizzare l’energia metabolica quando serve oppure sono presenti e inattivi perché coperti dall’antisigma. La polimerasi si lega con un fattore sigma (formazione oloenzima), riconosce il promotore da cui iniziare la trascrizione e, dopo una serie di inizi abortivi, trova quello giusto e inizia a formare l’mRNA. Un batterio inizia a sporulare quando capta segnali ambientali, ma come li capta? Tramite la presenza di proteine-recettori sulla membrana plasmatica ( istidina chinasi ); quando queste proteine cambiano conformazione legandosi al segnale ambientale, si autofosforilano (meccanismo di trasduzione del segnale: il segnale viene raccolto e interpretato). Il gruppo fosfato può essere trasferito ad un’altra proteina detta “regolatrice della risposta”, che fa da interruttore (sistema a due componenti, Poshorelay) per l’espressione genica (geni utili in quel momento). Il ricevitore Spo0A del fosfato causa una cascata di fattori sigma espressi in specifici compartimenti cellulari, i quali permettono un’espressione differenziale tra la cellula madre e la spora portando alla formazione di tutte quelle strutture che nella spora sono presenti ma non nella cellula madre (nella cellula vegetativa c’è solo sigma A, mentre nella spora all’inizio solo sigma H, poi H e F, poi F e E, nella prespora E e G, poi A e K e poi c’è il rilascio della spora).
GERMINAZIONE
La germinazione è il processo che permette alla spora di dare nuovamente origine alla cellula vegetativa; è un processo più rapido (1h e 30 min). La germinazione avviene in presenza di appropriate condizioni ambientali; quando queste sono tornate favorevoli, la spora “espelle” una cellula vegetativa che da origine alle figlie dividendosi per normale scissione binaria. Un’endospora può rimanere quiescente per molti anni, prima di essere riconvertita passando per 3 fasi: Attivazione : un endospora non riesce a germinare se prima non viene attivata, anche se si trova in un ambiente favorevole. Questa attivazione è innescata da vari fattori tra i quali riscaldamento (per breve tempo) a temperatura subletale; il riscaldamento è ciclico; scaldando per la prima volta “sveglio” le spore, scaldando per la seconda volta “sveglio” le spore rimaste ma uccido le cellule vegetative, che iniziano la sporulazione ecc…. Germinazione : perdita di rifrangenza, resistenza, di dipicolinato di calcio e dei componenti della corteccia, aumento dell’attività metabolica e le SASP vengono degradate ed utilizzate come fonte di carbonio ed energia per la nuova cellula. Esocrescita : visibile rigonfiamento dovuto all’assorbimento di acqua e alla sintesi di RNA, proteine e DNA e il protoplasto sintetizza nuovi componenti emergendo dai resti del rivestimento della spora. 21 mar. ’ Altri esempi di differenziamento I batteri sono sempre stati ritenuti simmetrici ed in grado di dividersi attraverso la semplice scissione binaria con la conseguente creazione di cellule figlie identiche alla madre. Ogni batterio a forma di bastoncello è, tuttavia, asimmetrico con un nuovo polo creato durante la divisione cellulare e con un vecchio polo creato durante una precedente divisione. Per esempio, il Caulobacter crescentus sfrutta questa polarità per sostenere la sua motilità direzionale, per generare tassi di crescita variabili, per differenziare le dimensioni delle cellule e per produrre cellule figlie con destini diversi. Le cellule di C.crescentus sono dismorfe e possono essere peduncolate , non dotate di mobilità e provviste di un’appendice che permette di aderire a substrati solidi, e flagellate/sciamanti , dotate di un singolo flagello polare alla base del quale sono presenti pili. Questo dimorfismo è risultato di un processo di differenziamento che si attua ad ogni ciclo cellulare; solo le cellule peduncolate possono replicare il DNA e riprodursi per scissione binaria asimmetrica, al termine della quale si ottiene una cellula peduncolata e una flagellata. I mixobatteri sono batteri del suolo gram negativi, caratterizzati da una “vita sociale” in quanto si muovono in sciami su superfici solide per scivolamento. Questi sciami sono formati da migliaia di cellule che secernono enzimi idrolitici che lisano le altre cellule microbiche incontrate lungo il percorso. Durante i periodi di scarsità di nutrienti, la crescita si arresta ed inizia il processo di differenziamento con la formazione di corpi fruttiferi contenenti spore. La spora che germina da origine a delle cellule vegetative che si muovo in maniera coordinata per ottimizzare l’approvvigionamento di cibo mentre, in condizioni di carenza, alcune cellule all’interno dei corpi fruttiferi lisano e rilasciano sostanze utilizzate come nutrimento dalle cellule sopravvissute. Gli Streptomiceti sono batteri gram positivi filamentosi nei quali la replicazione del DNA non è accompagnata dalla formazione di cellule distinte, ma dalla formazione di setti separatori in un micelio; la