Scarica Riassunto e domande di Biochimica e più Appunti in PDF di Biochimica solo su Docsity!
Biochimica
CAPITOLO 1 – Biomolecole e cellule
La biochimica è lo studio della chimica della vita. I quattro elementi più abbondanti sono: idrogeno ,
ossigeno , carbonio e azoto (proprietà in comune, formano prontamente legami covalenti mettendo
in compartecipazione delle doppiette di elettroni). Inoltre, C, N e O, possono mettere in
compartecipazione uno o due doppiette di elettroni formando legami semplici o doppi. C, N e H
sono gli elementi più leggeri capaci di formare legami covalenti (la forza di un legame covalente è
inversamente proporzionale ai pesi atomici degli elementi che lo formano e quindi formano legami
covalenti molto forti). Gli atomi di C sono in grado di legarsi fra loro (4 legami disponibili) e possono
formare degli scheletri lineari o ramificati di un’immensa varietà di molecole organiche diverse.
Inoltre, nella struttura delle molecole organiche possono essere introdotte molte specie diverse di
gruppi funzionali. Le molecole possono avere configurazioni diverse a causa:
della presenza di doppi legami intorno ai quali non vi è libertà di rotazione:
o si hanno molecole diverse: isomero cis o trans i quali hanno funzioni diverse;
dalla presenza di eventuali centri chirali:
o la molecola chirale non è sovrapponibile alla sua immagine speculare.
Le biomolecole sono ordinate in una gerarchia di crescente complessità molecolare; tutte derivano da
precursori semplici , a basso peso molecolare. Questi vengono poi convertiti dalla materia vivente
nelle biomolecole base che vengono legate covalentemente l’una all’altra a formare macromolecole.
Così, gli amminoacidi sono le unità base delle proteine, i nucleotidi degli acidi nucleici, i
monosaccaridi dei polisaccaridi e gli acidi grassi sono le unità base dei lipidi. Al livello superiore le
macromolecole si associano fra loro formando i sistemi macromolecolari , come le lipoproteine
(lipidi + proteine) e i ribosomi (acidi. nucleici + proteine). Il modo in cui si associano non è covalente,
sono infatti legati da forze non covalenti deboli , come ponti idrogeno , interazioni idrofobiche e
interazioni di van der Waals (le numerose interazioni deboli che si portano a sistemi molto stabili).
Inoltre, l’associazione non covalente è molto specifica ed è il risultato di un preciso adattamento
geometrico reciproco fra i vari componenti ( complementarietà ).
Le proteine sono le macromolecole più rappresentate nella cellula (+50% del peso secco) e sono i
prodotti diretti e gli effettori dell’azione dei geni. Gli acidi nucleici servono a conservare e a
trasmettere l’informazione genetica (acidi nucleici e proteine sono macromolecole informazionali):
Ciascun acido nucleico contiene quattro o più nucleotidi disposti in una specifica sequenza e anche
ogni proteina contiene una sequenza specifica composta da circa 20 amminoacidi diversi. I
polisaccaridi hanno due funzioni essenziali, ovvero il deposito di materiale destinato a produrre
energia per l’attività cellulare e il fatto di essere elementi strutturali extracellulari. Infine i lipidi sono
i principali componenti strutturali delle membrane e hanno la funzione di deposito di energia.
CAPITOLO 2 – L’acqua
L’acqua rappresenta il 70-90% del peso della maggior parte delle forme di vita. È una molecola molto
reattiva. Quando due molecole d’acqua si avvicinano fra loro, si hanno delle attrazioni elettrostatiche
fra la carica parziale negativa di O di una molecola di acqua e la parziale carica positiva di H della
molecola adiacente. Questa attrazione provoca una ridistribuzione delle cariche in entrambe le
molecole che aumenta la loro interazione ( legame idrogeno). Ciascuna molecola d’acqua è in grado
di formare legami idrogeno con quattro molecole circostanti.
I legami idrogeno non avvengono soltanto nell’acqua, essi tendono a formarsi fra un piccolo atomo
altamente elettronegativo e un atomo di idrogeno legato covalentemente a un altro atomo
elettronegativo; si possono formare anche fra due parti della stessa molecola. Quando due molecole
sono unite da un solo legame idrogeno, il legame sarà molto debole in un sistema acquoso perché le
molecole d’acqua circostanti competono con le molecole di soluto per formare legami idrogeno.
Acqua come solvente : molti sali e altri composti ionici si dissolvono facilmente in acqua rispetto ad
altri solventi non polari. Il reticolo cristallino dei sali è unito da attrazioni elettrostatiche molto forti
ed è necessaria un grande quantità di energia per staccare gli ioni l’uno dall’altro: l’acqua dissolve
questi sali perché le forti attrazioni elettrostatiche fra i dipoli dell’acqua e gli ioni positivi e negativi
(del sale) superano la tendenza degli ioni ad attrarsi, quindi l’acqua tende ad opporsi all’attrazione
elettrostatica fra ioni positivi e negativi (questa tendenza è detta costante dielettrica D ).
Interazioni idrofobiche : l’acqua solubilizza sotto forma di micelle molti composti che contengono
sia gruppi fortemente apolari che gruppi fortemente polari. I gruppi polari (idrofili) sono esposti e
formano legami idrogeno con le molecole d’acqua, le catene idrocarburiche apolari insolubili
(idrofobe) sono nascoste all’interno. Queste micelle si formano spontaneamente. Dentro le micelle ci
sono forze addizionali fra le strutture idrocarburiche dovute a interazioni di van der Waals (non c’è
vero legame stechiometrico fra le code idrocarburiche, quindi si usa il termine interazione idrofobica).
CAPITOLO 3 – Le proteine
Le proteine sono le molecole organiche più abbondanti nelle cellule e costituiscono il 50%+ del peso
secco. Si trovano in ogni parte in quanto fondamentali per tutti gli aspetti della struttura e della
funzione delle cellule. Vi sono molti tipi, ciascuna specializzata per una diversa funzione biologica.
Composizione : tutte contengono carbonio , idrogeno , azoto e ossigeno (quasi tutte zolfo). Alcune
contengono elementi addizionali, quali fosforo , ferro , zinco e rame. I pesi molecolari sono molto
alti, ma in seguito a idrolisi acida danno luogo a un gruppo di composti organici semplici a basso
peso molecolare, gli α-amminoacidi. Queste contengono
almeno un gruppo α-carbossilico e un gruppo α-aminico , ma
differiscono fra loro nella struttura dei loro gruppi R ( catene
laterali ). Nelle proteine si trovano comunemente 20 diversi
amminoacidi. Nelle molecole proteiche gli amminoacidi sono
legati covalentemente a formare catene molto lunghe non
ramificate con legami chiamati legami peptidici formati per
eliminazione di H
2
O dal gruppo carbossilico di un amminoacido
e dal gruppo α-aminico del successivo ( condensazione ).
Legame peptidico : Planare e rigido (non è permessa una libera rotazione attorno al legame C-N, il
quale ha carattere di doppio legame); il legame C=O si comporta come legame singolo e inoltre il
gruppo imminico (-NH-) non ha tendenza a ionizzarsi o a protonarsi nell’ambito del pH. Per
quanto riguarda l’isomeria cis e trans del legame polipeptidico, è favorita dal punto di vista
termodinamico la forma trans del legame peptidico dovuto al fatto che nella posizione cis c’è un
enorme ingombro sterico che destabilizza la struttura e quindi non è termodinamicamente favorita
(solo la prolina si trova in conformazione cis). I legami peptidici sono molto resistenti e non si
rompono ( proteasi ) anche se esposti ad alta temperatura o alte concentrazioni di agenti denaturanti
come l’urea. Poiché i gruppi amminico e carbossilico degli amminoacidi sono impegnati nel legame
peptidico le proprietà delle proteine sono determinate dalle catene laterali.
Struttura delle proteine
Struttura primaria : La struttura primaria è la struttura covalente che corrisponde alla precisa
sequenza degli amminoacidi nella sua catena. Per convenzione i biochimici definiscono la sequenza
delle proteine con l’orientamento N→C (aminico→ carbossilico). La sequenza primaria di una
proteina determina il suo ripiegamento che ne determina la funzione.
Strutture secondarie : Sono le sue strutture tridimensionali locali, le conformazioni assunte dai vari
tratti della catena. Conformazione : indica la disposizione spaziale di gruppi sostituenti che sono
liberi di assumere molte posizioni diverse, senza rottura di legami, in seguito alla rotazione intorno
ai legami singoli presenti nella molecola. Tra le innumerevoli conformazioni teoricamente possibili
per ogni proteina, solo una (quella associata alla sua funzionalità) è definita conformazione nativa.
La conformazione di una proteina è stabilizzata da interazioni deboli (legami a idrogeno, legami
ionici e interazioni idrofobiche) e gli unici legami covalenti in grado di influenzare la
conformazione proteica sono i ponti disolfuro. La catena polipeptidica di una proteine assume
diverse conformazioni le quali conferiscono l’attività biologica, esse sono sostanzialmente di tre tipi:
α-elica : è destrorsa e gli amminoacidi si avvolgono attorno ad un asse centrale con i gruppi R
in catena laterale che sporgono radialmente. Ponendo in alto l’amminoacido N-terminale, i
legami peptidici ruotano in modo da rivolgere tutti i carbonili C=O in basso per formare
legami idrogeno con i gruppi N-H (tutti rivolti in alto) del quarto amminoacido che segue,
sull’ansa sottostante. Questa leggera sfasatura fa sì che in ogni spira vi siano un po’ meno di
quattro amminoacidi (3,6 per la precisione). La lunghezza di un singolo giro è di 5,4 Å.
Conformazione β : è la conformazione più frequente nelle proteine caratterizzata da un
andamento a zig-zag dello scheletro covalente peptidico. In questa struttura i legami peptidici
planari assumono la configurazione di un foglietto pieghettato e rivolgono gli ossigeni dei
carbonili C=O alternativamente da un lato e dall’altro e così pure fanno gli idrogeni dei gruppi
N-H. Inoltre, i gruppi R delle catene laterali sporgono alternativamente sotto e sopra il piano
del foglietto. Le catene polipeptidiche sono disposte una a fianco all’altra formando un
modulo definito foglietto β. Il foglietto β è stabilizzato da legami a idrogeno tra filamenti β
adiacenti. Le catene polipeptidiche adiacenti in un foglietto β possono essere parallele (stesso
orientamento, legami più deboli) o antiparallele (orientamento opposto).
Ripiegamento β : è un elemento di struttura secondaria, formato da 4 a.a. e stabilizzato da un
legame H, che consente la completa inversione di direzione della catena polipeptidica e quindi
un suo forte impaccamento. Spesso rappresenta la coniugazione tra due α-eliche o tra due
filamenti β antiparalleli. Sono tipici delle proteine globulari.
o In alcune proteine, l’unica struttura secondaria regolare è l’α-elica: Mioglobina.
o In alcune proteine, l’unica struttura secondaria regolare è il foglietto β: Avidina, proteina
presente nell’albume dell’uovo che lega la Biotina (vitamina H, vitamina B7).
o Nella maggior parte delle proteine, sono presenti α-eliche e foglietti β: in media, il 31% delle
proteine globulari è costituito da α-eliche, il 28% da foglietti β, il resto da random coil
(strutture secondarie non ripetitive quelle sequenze che adottano una conformazione stabile
ad anse e avvolgimenti) e loop.
Struttura terziaria : Descrive l’organizzazione nello spazio tridimensionale di tutti gli atomi della
catena polipeptidica organizzati in distinti elementi di struttura secondaria e in dipendenza della sua
specifica struttura primaria. In condizioni fisiologiche la struttura tridimensionale che si ottiene è
detta nativa e ad essa è associata una funzione biologica.
Le principali interazioni chimiche che possono contribuire al mantenimento della struttura terziaria
(così come della struttura secondaria) sono il legame ad idrogeno, le interazioni ioniche, le interazioni
dipolo-dipolo. In alcune proteine sono presenti anche dei ponti disolfuro determinati dalla reazione
di ossidazione a carico di due residui laterali di cisteina.
Motivi : sono combinazioni regolari di strutture secondarie (α-eliche o β-foglietti) che hanno
una topologia particolare e sono organizzati in una struttura tridimensionale caratteristica.
Molte proteine contengono uno o piò motivi;
Domini : unità funzionali e/o strutturali distinte di una proteina. Sono responsabili di una
particolare funzione o interazione, contribuendo al ruolo globale di una proteina.
Struttura quaternaria : La struttura quaternaria si riferisce solo alle proteine composte di più
catene o subunità ; definisce il modo in cui le varie subunità si incastrano una nell’altra (le interazioni
possono essere legami deboli come legami idrogeno e forze di Van der Waals, oppure forti ossia
ionico o covalente). Le proteine con subunità identiche vengono dette oligomeri e le subunità che le
compongono protomeri. Un protomero, a sua volta, può essere costituto da una catena polipeptidica
o da alcune catene polipeptidiche diverse; ad esempio, l’emoglobina è un tetramero formato da due
dimeri (oligomero formato da due protomeri) di protomeri αβ.
Altre strutture : In base alla loro struttura complessiva, alcune proteine vengono indicate come
proteine fibrose (in genere hanno un solo tipo di struttura secondaria) di forma allungata, altre sono
indicate come proteine globulari , cioè di forma sferica (possono contenere più tipi di struttura
secondaria) come ad esempio sia alfa eliche che foglietti beta. Esiste poi un altro gruppo di proteine
che sono dette proteine di membrana , che sono cioè legate alla membrana plasmatica.
Appartengono a questo gruppo molti recettori (es. recettore per l’insulina), i trasportatori di
membrana (es. i GLUT, la Na+/K+ ATPasi) e i canali ionici.
Denaturazione e folding
Folding : Processo attraverso il quale le proteine ottengono la loro struttura tridimensionale nativa
(biologicamente attiva). Avviene sia contemporaneamente alla sintesi proteica che alla fine di questa
(soltanto una volta terminato il folding le proteine possono assumere la loro funzione fisiologica).
Esistono fattori che ostacolano il ripiegamento corretto come la mancata sintesi contemporanea di
tutti i domini della proteina, la presenza di grandi quantità di macromolecole (possibili interazioni
scorrette), a causa dello stress dovuto ad esposizione a sostanze tossiche (radicale dell’ossigeno o
metalli pesanti) o presenza di mercaptano e urea ( esperimento di Anfisen ); Il folding è un processo
spontaneo in condizioni fisiologiche. Invece, la denaturazione ( unfolding ) è il processo inverso
ma non coinvolge legami peptidici dello scheletro polipeptidico (la proteina denaturata non è sempre
completamente srotolata, può anche avere conformazione parzialmente ripiegata). Può avvenire
rinaturazione ( refolding ) della proteina quando la denaturazione è reversibile. La denaturazione
comporta perdita dell’attività biologica (inattivazione degli enzimi), coagulazione , aggregazione
e l’incremento della sensibilità agli enzimi digestivi (aumento della digeribilità con la cottura).
Foldasi e chaperon molecolari : in vivo il ripiegamento delle proteine necessita di un
macchinario di folding mediato da foldasi e chaperon molecolari. Le foldasi aumentano la
velocità del processo di ripiegamento. I chaperon molecolari intervengono sia in condizioni
normali che di stress; assicurano il raggiungimento e il mantenimento del corretto stato
conformazionale delle catene polipeptidiche appena sintetizzate; stabilizzano le proteine
danneggiate formatosi a seguito di stress chimici o fisici, facilitandone la degradazione o la
loro rinaturazione; impediscono le aggregazioni dei siti intermedi del folding.
Capitolo 4 – Funzioni delle proteine
Funzione di sostegno strutturale
Le proteine che svolgono principalmente un ruolo di sostegno strutturale sono le proteine fibrose :
Hanno forma allungata e sono caratterizzate da un unico elemento della struttura secondaria ripetuto
(o tutte in conformazione alfa o tutte in conformazione beta). Esse sono insolubili in acqua a causa
dell’elevata percentuale di amminoacidi idrofobici all’interno e sulla superficie della proteina, sono
fisicamente resistenti ed elastiche , svolgendo un ruolo meccanico-strutturale. Proteine fibrose sono:
α-Cheratina (struttura alfa): Componente principale di capelli, lana, penne, unghie, ecc. Sono
ricche di residui di cisteina e contengono quindi molti ponti disolfuro che aumenta la
resistenza alla trazione e le rende insolubili;
Collageno (il quale ha una sua propria forma che non è presente in nessun’altra proteina):
Principale componente proteico del tessuto connettivo (presente in tendini, legamenti, tessuto
sottocutaneo, ossa, muscoli, ecc.) rappresenta la proteina più abbondante nell’organismo.
α-Cheratina : Composta da due α-eliche destrorse che si avvolgono l’una sull’altra con
superavvolgimento sinistrorso ( coiled coil ). Le regioni delle due eliche a contatto sono ricche di
amminoacidi idrofobici. L’aggregazione di più coiled coil originano protofilamenti e protofibrille.
Sono ricche di residui Cys e in base al contenuto possono essere Hard keratin (alto contenuto di
Cys, si trova nei capelli, corna e unghie) e Soft keratin (basso contenuto di Cys, si trova nella pelle).
L’alto contenuto di ponti disolfuro rende le cheratine indigeribili da parte della maggior parte degli
animali; anche il nostro organismo non riesce a ridurre i ponti S-S e quindi non potendo modificare
la struttura rigida, non possiamo utilizzarla, nonostante sia una proteina fonte di aa essenziali.
Collageno : Un altro tipo principale di proteina fibrosa presente negli animali superiori, il collageno
del tessuto connettivo, ricopre tutti i muscoli ed è inoltre il principale costituente dei tendini. È
costituito da lunghissime eliche sinistrorse contenenti più di 1000 residui che unite da ponti idrogeno
si avvolgono a formare un’elica destrorsa a 3 filamenti chiamata tropocollageno (vera e propria fune
molecolare, la cui associazione con altre molecole di tropocollageno genera la fibra di collageno ).
Questa struttura è resa possibile dalla sequenza ripetuta Gly - X (spesso Pro) - Y (spesso Hyp/Hyl).
La catena polipeptidica del collageno è costituita per il 33% da residui di glicina ( Gly ) e per il 15-
30% da residui di Prolina ( Pro ) o 4-idrossiprolina (Hyp). Questi a.a. formano legami H che
stabilizzano il protocollageno e legami covalenti tra le molecole di tropocollageno nel collageno. La
catena polipeptidica contiene a.a. modificati a livello post-traduzionale, quali idrossiprolina ( Hyp )
e idrossilisina ( Hyl ) che vengono sintetizzati a partire da prolina e lisina dagli enzimi
Prolilidrossilasi e lisilidrossilasi. Queste idrossilasi richiedono la presenza di acido ascorbico
( vitamina C ) per svolgere la loro funzione e in sua assenza può sopraggiungere lo scorbuto ; in queste
condizioni, il collageno sintetizzato non è in grado di formare fibre in maniera corretta, conducendo
a lesioni cutanee, fragilità dei vasi sanguini, scarsa capacità di rimarginare le ferite e infine il decesso.
Il valore nutrizionale del collageno è molto limitato, sia per la sovrabbondanza di Gly, Pro, Hyp e
Hyl sia per la limitata azione proteolitica delle proteasi del tubo digerente sulla molecola di collageno.
La gelatina alimentare è costituita da collageno denaturato, portato in soluzione e gelificato.
Le molecole di tropocollageno si allineano in modo sfalsato a formare fibre di collageno. Le eliche
all’interno di una molecola di tropocollageno e molecole di tropocollageno in una fibra sono unite da
legami covalenti (legami crociati ) che coinvolgono le catene laterali di residui di lisina, idrossilisina
e istidina. Il numero di legami crociati aumenta con l’età , rendendo il collageno più rigido e fragile.
Funzione di difesa
Tutti i vertebrati hanno un sistema immunitario in grado di distinguere le molecole proprie da quelle
estranee e di distruggere ciò che viene riconosciuto estraneo. Si distinguono per tanto:
Molecole self : quelle che fanno parte del nostro organismo;
Molecole non-self : sono le molecole che non fanno parte del nostro organismo; s
o Si attiva la risposta immunitaria che coinvolge sistemi di risposta in difesa di elementi
estranei, quali batteri o virus.
La difesa contro i microbi è mediata dalle reazioni precoci dell’ immunità innata (meccanismi di
difesa cellulari o biochimici preesistenti all’infezione e pronti a reagire con rapidità limitando
l’invasione degli elementi estranei) e da altre più tardive dell’ immunità adattativa (aumenta la
capacità difensiva ad ogni successiva esposizione a un particolare agente infettivo la quale si sviluppa
in risposta ad una specifica infezione e si adatta ad essa). Si divide a sua volta in immunità umorale
e in immunità cellulo-mediata. Le cellule che agiscono in questo tipo di risposta immunitaria sono
dette linfociti. Gli anticorpi sono le proteine per l’immunità adattativa umorale ( del sangue ).
Caratterizzata da specificità per molecole diverse e dalla capacità di memoria cioè quando un
individuo viene a contatto per la prima volta ( immunizzazione primaria ) con una molecole non-
self, la risposta immunitaria agisce dopo 7-14 giorni; se in seguito questo individuo viene a contatto
con la stessa molecola ( immunizzazione secondaria ), la risposta immunitaria sarà rapida e intensa.
Quando i linfociti B entrano a contatto con la sostanza estranea, producono grandi quantità di
immunoglobuline, di cui una parte di esse viene escreta fuori dalla cellula nel plasma, mentre l’altra
parte si lega sulla superficie di questi linfociti B produttori ( cellule della memoria ).
Antigene : qualsiasi molecola (ogni natura chimica) o agente patogeno in grado di indurre una
risposta immunitaria è detta antigene (molecole non-self). Nella definizione di antigene bisogna
distinguere due caratteristiche fondamentali: l'antigenicità (o specificità antigenica che rappresenta
la capacità dell'antigene di combinarsi specificamente con anticorpi e recettori per l'antigene) e
l'immunogenicità (o potere immunogeno che indica la capacità dell'antigene di indurre una risposta
immunitaria). La piccola parte di antigene che lega l'anticorpo specifico è detta epitopo o
determinante antigenico (la singola molecola di antigene può contenere diversi epitopi riconosciuti
da anticorpi differenti) e si divide a sua volta in epitopo o determinante lineare (caratterizzati da
una specifica sequenza lineare amminoacidica) o epitopo o determinante conformazionale
(riconosciuti dal sistema immunitario come complessi tridimensionali).
Anticorpi : sono proteine solubili prodotte dalle plasmacellule del sistema linfatico di una data specie
di vertebrati, coinvolte nella risposta immunitaria verso agenti antigeni e sostanze riconosciute come
estranee. Sono dette anche immunoglobuline (forma globulare). Sono quindi caratterizzate dalla
disposizione degli amminoacidi polari/carichi sulla superficie e quelli apolari all’interno, nel core
della proteina. Costituiscono il 20% delle proteine del sangue. La funzione degli anticorpi è quella di
riconoscere/legare una sostanza estranea all’organismo e indirizzarla alla degradazione.
Struttura degli anticorpi : quattro catene polipeptidiche ( struttura quaternaria ), sono 2
leggere e 2 pesanti ; disposte internamente le catene pesanti , legate insieme da ponti S-S; le
catene leggere (più piccole e corte), si trovano nelle braccia della Y e sono poste esternamente,
legate alle catene pesanti mediante ponti S-S. Si distinguono regioni variabili (cambiano da
anticorpo ad anticorpo, dipende dalla sostanza estranea e determinano la specificità) e regioni
costanti (uguali per tutte le Ig della stessa specie) in entrambe le catene. Le immunoglobuline G
(IgG) sono la classe principale di molecole anticorpali e sono le proteine più abbondanti nel
plasma. Ogni catena pesante è ripiegata in 4 domini mentre ogni catena leggera è ripiegata in
2 domini. Conformazione unicamente a foglietto-β , in particolare con filamenti antiparalleli.
Organizzazione del sarcomero : Il sarcomero è costituito principalmente da actina e miosina. Queste
due proteine si alternano all’interno del sarcomero. Le linee Z sono gli estremi del sarcomero. Al
centro è presente una banda M alla quale è legata la miosina che si estende dal centro verso destra
e verso sinistra, ma non arriva mai fino alle linee Z. L’actina, invece, parte dalla linea Z e va verso
il centro , senza raggiungere la banda M. La proteina titina mantiene i fasci di miosina al centro del
sarcomero e si estende per tutta la lunghezza del sarcomero (da una linea Z alla successiva); la titina
è responsabile dell’allungamento del muscolo rilassato ed è infatti particolarmente elastica.
Contrazione muscolare : Le modificazioni conformazionali della testa della miosina accoppiate alle
fasi del ciclo di idrolisi dell’ATP determinano la dissociazione della miosina da una subunità di actina
per associarsi subito dopo con un’altra subunità di actina lungo il filamento (i filamenti di actina e di
miosina scorrono gli uni rispetto agli altri). L’interazione tra actina e miosina è regolata in modo che
la contrazione possa avvenire solo in risposta a specifici segnali provenienti dal sistema nervoso. La
regolazione è mediata da tropomiosina e troponina (tropomiosina interagisce con l’actina
nascondendo i siti di interazione con le teste di miosina). Quando si ha la contrazione muscolare,
l’impulso nervoso induce il rilascio dello ione Ca
2+
dal reticolo sarcoplasmatico. Il Ca
2+
si lega alla
troponina inducendo una variazione conformazionale del complesso tropomiosina-troponina
esponendo i siti di legame dell’actina per le teste di miosina permettendo la contrazione.
Fonti energetiche della contrazione : L’ATP fornisce l’energia necessaria per la contrazione
muscolare e se ne rifornisce attraverso il processo della glicolisi aerobia (con ossigeno) procede fino
alla formazione di 38 molecole di ATP mentre con la glicolisi anaerobia ( fermentazione ) si forma
acido lattico e 2 molecole di ATP. Nel muscolo il glucosio è immagazzinato sotto forma di glicogeno
(forma di energia accumulata nel muscolo che gli permette di contrarsi anche se non disponibile il
glucosio ematico). Dopo la morte dell’animale, in seguito all’arresto della circolazione sanguigna, il
muscolo si trova in assenza di ossigeno ( anossia ) instaurando così una glicolisi anaerobia post-
mortem con produzione di acido lattico (pH da 7 a 5,5). La riduzione del pH è fondamentale per la
buona conservazione della carne in quanto limita la proliferazione microbica.
Trasformazione del muscolo in carne : La carne degli animali non è immediatamente edibile, è
necessario tempo affinché le carni diventino adatte al consumo, durante il quale la struttura e la
composizione del muscolo subiscono modificazioni fisiche e chimiche. Il sistema nervoso rimane
eccitabile per circa 1,5 h dopo la macellazione e il muscolo può contrarsi, successivamente il muscolo
perde la sua flessibilità e si instaura il rigor mortis : una volta esaurite le scorte di ATP, il muscolo
non ha più energia per favorire la contrazione e si crea un’interazione irreversibile tra actina e miosina
(si legano saldamente l’una all’altra, formando il complesso actomiosina ). Dopo il rigor mortis inizia
un processo di proteolisi da enzimi contenuti nei lisosomi delle cellule che prevede la degradazione
enzimatica dei filamenti di actina e miosina con conseguente rilascio della fibra contratta.
Funzione di trasporto
Alcune proteine ( di trasporto ) si legano a delle sostanze per consentirne un efficiente trasporto
attraverso il torrente circolatorio. L’ossigeno (O2) è un gas fondamentale per la vita perché consente
l’ossidazione dei nutrienti per ricavarne energia ( ATP ). L’ossigeno è pochissimo solubile in acqua e
quindi non può essere trasportato in modo efficace per semplice diffusione. La maggior parte dei
vertebrati produce Mioglobina ed Emoglobina che consentono l’efficiente solubilizzazione e
stoccaggio dell’ossigeno. I vertebrati dispongono di un sistema circolatorio ( circolazione doppia )
in cui il sangue viene pompato dal cuore all’interno dei vasi sanguigni ( circolazione sistemica )
mentre i vasi sanguigni a livello di polmoni e tessuti/organi periferici hanno una struttura capillare
(circolazione polmonare) che consente un efficiente scambio di gas con il sangue e si distingue
anch’esso in sangue venoso (povero di O2 e ricco di CO2) e sangue arterioso (ricco di O2).
Cuore dei mammiferi : costituito da 4 camere e 2 atri (superiori) comunicano tramite valvole con i
due ventricoli. Il cuore viene rifornito di ossigeno e di sostanze nutritive tramite le arterie
coronarie , che si ramificano dal tratto iniziale all’aorta.
Cuore destro ( atrio e ventricolo destro ): Il sangue deossigenato (ricco di CO2) del circuito
polmonare (piccola circolazione), viene pompato verso i polmoni; negli alveoli l’anidride
carbonica viene ceduta e il sangue si arricchisce di ossigeno, il quale viene messo a
disposizione della grande circolazione;
Cuore sinistro ( atrio e ventricolo sinistro ): Il sangue ossigenato (ricco di O2 e sostanze
nutritive) del circuito sistemico (grande circolazione), viene pompato verso tutti i tessuti, dove
cede ossigeno e sostanze nutritive raccogliendo invece quelle di rifiuto.
Sangue: Tessuto connettivo fluido , costituisce circa l’8% del peso corporeo (in un uomo adulto il
volume sanguineo è di circa 5-6 L). Nel sangue si distinguono due componenti diverse che possono
essere separate tramite centrifugazione; una matrice fluida ( plasma , 54 - 58% del volume) contenente
proteine plasmatiche e la parte corpuscolata ( elementi figuranti) , cioè cellule o frammenti di cellule.
La percentuale degli elementi figuranti si chiama ematocrito ed è composto da piastrine , leucociti
( globuli bianchi ) ed eritrociti ( globuli rossi ) i quali sono cellule particolari senza nucleo e molti
organelli cellulari, hanno forma biconcava (aumenta la superficie di scambio con il plasma e sono
prodotti dal midollo osseo (contengono un’elevatissima concentrazione di emoglobina ).
Proteine plasmatiche : Fibrinogeno (prodotto dal fegato, coinvolto nella coagulazione del sangue),
le α-globuline (comprendono ormoni proteici deputati al trasporto di ormoni insolubili nel sangue
come le HDL che trasportano i grassi e il colesterolo tra i vari tessuti) le β-globuline (lipoproteine
deputate al trasporto di grassi e di colesterolo, vitamine liposolubili o ioni poco solubili in acqua), le
Immunoglobuline (costituiscono gli anticorpi, sono proteine prodotte dai linfociti B per la difesa
dell’organismo) e l’ Albumina , (svolge il duplice ruolo di trasportare gli acidi grassi liberi mobilizzati
dal tessuto adiposo e di dare un importante contributo alla pressione osmotica del sangue).
Globine : Sono una famiglia di proteine con struttura primaria, secondaria e terziaria simili. Presenti
in alcuni batteri e in tutti gli eucarioti, la maggior parte lega e trasporta l’ossigeno; sono costituite da
una parte proteica (la globina ) e una non proteica, il gruppo eme (gruppo prostetico, molecola
organica legata covalentemente alla catena polipeptidica della proteina, la quale svolge delle
importanti funzioni all’interno della proteina). Nei mammiferi la mioglobina (monomerica, 1 gruppo
eme), presente nel tessuto muscolare, favorisce la diffusione dell’ossigeno, mentre l’ emoglobina
(tetramerica, due catene α + 2 catene β, 4 gruppi eme) è responsabile del suo trasporto nel sangue.
All’interno del gruppo eme (deriva da una molecola chiamata protoporfirina ) è presente il ferro
nella forma Fe^2+ ed è in grado di legare l’ossigeno in modo reversibile. La forma ossidata Fe^3+
non è in grado di legare l’ossigeno.
Emoglobina : I globuli rossi trasportano la maggior parte dell’ossigeno presente nel sangue.
L’emoglobina contiene 4 catene (2 alfa + 2 beta) e 4 gruppi eme con cui lega 4 molecole di ossigeno
ed esiste in due diverse conformazioni: T (teso) e R (rilassato). L’ossigeno lega sia lo stato T che lo
stato R ma ha maggiore affinità per lo stato R. Il legame dell’ossigeno all’emoglobina è di tipo
cooperativo : inizialmente una molecola di ossigeno si lega ad una subunità nella conformazione T,
questo legame determina una modificazione conformazionale dallo stato T a quello R che viene
comunicato alle subunità adiacenti aumentando la loro affinità per l’ossigeno. L’emoglobina è un
tetramero , il legame dell’ossigeno essendo cooperativo , aumenta l’affinità tanto più è alto il numero
di molecole di ossigeno legate (massimo 4). La mioglobina invece è un monomero , il legame
dell’ossigeno non è cooperativo.
Temperatura e pH : La velocità di tutte le reazioni chimiche aumenta con l’aumentare della
temperatura. Anche gli enzimi obbediscono a questa legge, tuttavia quest’ultimi sono proteine e
quindi sensibili alla denaturazione termica. Come tutte le proteine, gli enzimi hanno una struttura
terziaria che è sensibile al pH e denaturano a pH molto bassi o molto alti. In generale, un enzima è
attivo in intervallo di 4-5 unità di pH attorno al suo massimo (attività diminuisce se ci si allontana).
Inibizione enzimatica : Molte sostanze alterano l’attività degli enzimi, legandosi ad esso in modo da
influenzare il legame del substrato e/o il numero di turnover dell’enzima stesso. I composti che
riducono l’attività degli enzimi sono noti come inibitori. E sono suddivisi in due classi:
Inibitori reversibili : interazione tra proteine e molecole inibitrici mediata da interazioni deboli,
da legami quindi reversibili. Si distinguono in:
o Inibizione competitiva : Competono con il substrato per il sito attivo. Sono in genere
analoghi strutturali del substrato. Rimuovono l’enzima dal ciclo catalitico naturale.
L’effetto può essere annullato impiegando concentrazioni molto elevate di substrato;
o Inibizione non competitiva : Si legano all’enzima solo dopo che si è già formato il
complesso ES, ma non all’enzima libero. Il legame dell’inibitore non competitivo, che
non necessariamente deve essere simile al substrato, può causare una distorsione
strutturale del sito attivo, rendendo l’enzima cataliticamente inattivo;
o Inibizione mista : Sia enzima libero, sia il complesso enzima-substrato legano l’inibitore.
Inibitori irreversibili : si combinano in modo irreversibile o distruggono un gruppo funzionale
dell’enzima che è essenziale per la sua attività catalitica. Molti sono dei veleni.
o reagenti gruppo-specifici : DIFP; disattiva irreversibilmente l’enzima acetilcolinesterasi,
sistema fondamentale per il funzionamento del sistema nervoso;
o analoghi del substrato : Si legano covalentemente ai residui amminoacidici del sito attivo
inibendone l’attività (chimotripsina + TPCK);
o inibitori suicidi : Sono in grado di portare avanti le prime tappe della reazione enzimatica,
ma invece di essere trasformati nel prodotto finale della reazione, vengono convertiti in
composti molto reattivi che si combinano in modo irreversibile con l’enzima.
Enzimi allosterici : Presentano oltre al sito di legame per il substrato, un sito di legame per una
molecola definita modulatore allosterico positivo o negativo. Un sito allosterico di un enzima
è un sito lontano, distinto dal sito attivo a cui si lega l’effettore allosterico. Il legame dell’effettore
allosterico provoca una modificazione conformazionale che fa cambiare l’attività dell’enzima
(quindi modifica il sito attivo) e può essere:
o Inibitore allosterico = Modulatore allosterico negativo;
o Attivatore allosterico = Modulatore allosterico positivo.
Enzimi digestivi : L’idrolisi spontanea del legame peptidico ( proteolisi ) è una reazione spontanea ma
lentissima in condizioni fisiologiche; la velocità aumenta se catalizzata dalle proteasi (o peptidasi )
e questo processo riveste importantissimi ruoli fisiologici. Diverse proteasi sono prodotte nei diversi
tratti dell’apparato digerente; nello stomaco, l’ingresso di una proteina stimola la secrezione
dell’ormone gastrina che stimola la secrezione di acido cloridrico (denatura le proteine) e la sintesi
di pepsinogeno (attivato a pepsina con meccanismo autocatalitico). Nel duodeno invece è stimolata
la sintesi dell’ormone colecistochinina che stimola la secrezione di diverse proteasi pancreatiche :
chimotripsina , tripsina , elastasi e carbossipeptidasi ; la loro specificità è determinata dalla natura
chimica della catena laterale dell’aminoacido che partecipa, col suo gruppo carbossilico, alla
formazione del legame peptidico. In particolare, nel sito attivo delle tre proteasi si deve posizionare
con corretta orientazione il legame che deve subire rottura idrolitica. Nel sito attivo è anche presente
una cavità di legame, con diverse caratteristiche per ognuna, in grado di legare preferenzialmente una
catena laterale amminoacidica grande e apolare, come nel caso della chimotripsina, o carica
positivamente, come nel caso della tripsina, o di piccole dimensioni, come nel caso della elastasi.
Attivazione proteolitica : Molti enzimi vengono attivati irreversibilmente mediante idrolisi di un
legame peptidico presente nei precursori inattivi detti zimogeni o proenzimi. Un tipico esempio di
attivazione proteolitica è quella degli zimogeni pancreatici. Quando sono liberati nel duodeno, il
tripsinogeno viene attivato da un’enteropeptidasi in tripsina che a sua volta attiva mediante taglio
proteolitico la chimotripsina, la carbossipeptidasi e l’elastasi. Nel pancreas le proteasi sono
ulteriormente inattivate mediante il legame di specifici inibitori proteici.
Intolleranza al lattosio : Stato patologico associato alla incapacità di digerire il lattosio causata da
una carenza di lattasi , una glicosidasi (beta-galattosidasi) che catalizza la rottura idrolitica del legame
glicosidico presente nel lattosio, con liberazione dei monosaccaridi glucosio e galattosio assorbiti poi
nel tratto intestinale. L’attività della lattasi nei villi intestinali dell’intestino tenue inizia a diminuire
nella maggior parte delle persone circa a 2 anni di vita con una riduzione geneticamente progressiva.
Nei soggetti intolleranti al lattosio si osserva una forte riduzione o totale perdita della attività lattasica.
La scarsa o assente attività lattasica nell’intestino tenue permette al lattosio (relativamente insolubile
e non assorbibile come tale da parte della mucosa intestinale) di passare indigerito nel colon. Qui, lo
zucchero viene fermentato dalla flora batterica intestinale fino a produrre gas ed acidi organici,
responsabili di reazione infiammatoria con conseguente sintomatologia a livello intestinale.
Capitolo 5 - Il Metabolismo
Il metabolismo è l’insieme dei processi biochimici ed energetici che si svolgono negli organismi
viventi. Le reazioni enzimatiche sono organizzate in vie metaboliche e il metabolismo consta di due
processi opposti che si svolgono in contemporanea nella cellula:
Catabolismo : ottengono molecole dall'ambiente (complesse) e le degradano per ottenere
molecole semplici ed energia.
Anabolismo : molecole semplici + ATP per formare (processi di costruzione) molecole
complesse utili all'organismo e alla cellula.
Gli obbiettivi del metabolismo sono ottenere energia chimica dall’ambiente, convertire molecole dei
nutrienti in molecole utili all’organismo e polimerizzare precursori monomerici per ottenere
macromolecole quali proteine, acidi nucleici e polisaccaridi. Negli organismi avvengono sempre
processi esoergonici che forniscono l’energia necessaria per permettere i processi endoergonici.
L’energia della cellula è presente sotto forma di ATP (adenosina tri-fosfato) presentante legami ad
alta energia; dopo la sua formazione viene immediatamente degradata e mai conservata (non esistono
riserve di ATP) in quanto ha troppe funzioni ed un eccesso potrebbe causare problemi all’interno
della cellula. Quando si rompono i legami viene liberata energia chimica che permette la formazione
o la rottura di altri legami (ATP + Glucosio → Glucosio- 6 - P + ADP). Una molecola di ATP viene
consumata nell’arco di 1 minuto dalla sua produzione (catabolismo e anabolismo). Le vie possono
avere composti di partenza e arrivo uguali, ma devono avere almeno un enzima specifico per una
delle due vie, queste devono inoltre essere irreversibili e ciò è permesso dal fatto che sono
termodinamicamente favorevoli in una sola direzione (tappa regolata).
Catabolismo : Insieme dei processi metabolici in grado di abbattere le grandi molecole. Questi
includono la scomposizione e l’ ossidazione delle molecole assunte con l’alimentazione. Il
catabolismo completo dei nutrienti porta alla formazione di CO 2
, H
2
O e NH 3
e alla produzione di
ATP. Il NAD
+
e il FAD sono le molecole che trasportano gli elettroni ceduti dai nutrienti che si
ossidano. I processi di ossidazione devono essere sempre accompagnati da processi di riduzione
ovvero la riduzione dei trasportatori di elettroni NAD
+
(forma ossidata) → NADH (forma ridotta).
Monosaccaridi (zuccheri semplici): Possono contenere un
numero variabile di atomi di carbonio (più comuni hanno 5 o 6
atomi di carbonio). Gli enantiomeri (isomeri che sono l’immagine
speculare l’uno dell’altro) sono indicati come D- e L- (vedi anche
gli amminoacidi). I glucidi naturali sono enatiomeri D. Il
glucosio è l’esoso più comune insieme al fruttosio e rappresenta
la principale fonte energetica della maggior parte degli organismi.
È il prodotto principale della fotosintesi. I monosaccaridi con 5 o
più atomi di carbonio possono assumere una struttura ciclica.
Dalla ciclizzazione si determina la formazione di due possibili
anomeri ( α e β ) che si distinguono per l’ orientamento del
gruppo OH legato al carbonio 1. Le due forme si convertono
l’una nell’altra. L’anomero α del glucosio è il 36% in soluzione.
Tutte le cellule contengono glucosio che sfruttano come fonte di
energia. Il glucosio esiste in soluzione in forma lineare ( 1% ) e
nella forma più stabile ad anello ( 99% ).
Aldosi e chetosi : Carboidrati = Idrati di carbonio. Sono poliidrossi-aldeidi e poliidrossi-chetoni.
I carboidrati aldeidici vengono chiamati aldosi mentre quelli chetonici sono detti chetosi.
Disaccaridi : Consistono di due monosaccaridi uniti da legame glicosidico tramite condensazione
( ossigeno fa da ponte tra i carboni dei due zuccheri coinvolti). In questo modo almeno uno dei due
carboni anomerici è «bloccato». Il legame glicosidico può essere α o β a seconda che la molecola
che contribuisce con il proprio carbonio 1 al legame sia α o β glucosio. I più comuni disaccaridi sono
il maltosio , il lattosio e il saccarosio.
Polisaccaridi : L’ amido è il più abbondante nelle piante e il glicogeno negli animali, sono in genere
depositati sotto forma di grossi granuli nel citoplasma delle cellule. Quando vi è sovrabbondanza di
glucosio , le unità di esoso sono immagazzinate per mezzo dell’unione enzimatica a estremità di
catene di amido o di glicogeno ; quando sono necessarie per il metabolismo, le unità vengono
rilasciate enzimaticamente per essere usate come fonte di energia.
Amido, Glicogeno e Cellulosa : L’ amido è formato da amilosio e amilopectina ; il primo è costituito
da lunghe catene prive di ramicazioni e le unità monomeriche sono unite in legame α(1→4).
L’amilopectina è altamente ramificata, presenta legami α(1→4) nello scheletro, ma i punti di
ramificazione sono costituiti da legami α(1→6). Il glicogeno invece è il polisaccaride principale di
deposito delle cellule animali (particolarmente abbondante nel fegato). Nelle cellule epatiche il
glicogeno è presente in grossi granuli, che sono a loro volta ammassi di granuli più piccoli composti
di singole molecole altamente ramificate. Come l’amilopectina, è un polisaccaride di D-glucosio in
legame α(1→4) ma è più ramificata e compatta; infatti, le ramificazioni α(1→6) si trovano ogni 8- 12
residui di glucosio. Cellulosa : Zucchero strutturale delle piante, è un polimero lineare.
Digestione dei carboidrati : Le cellule intestinali possono assorbire solo monosaccaridi, quindi i
disaccaridi, oligosaccaridi e polisaccaridi devono essere prima idrolizzati nelle componenti
monosaccaridiche. La digestione dell’amido inizia nella cavità orale, continua nel duodeno e si
completa a livello degli enterociti.
- Nella cavità orale troviamo l’ amilasi salivare che non riesce a digerire completamente le
molecole in quanto attacca solo i legami α(1→4)-glicosidici e non quelli α(1→6) liberando
maltosio ed isomaltosio (disaccaridi costituiti dall'associazione di due unità di glucosio,
rispettivamente unite da legami α-1,4 e α-1,6), maltotriosio (tre molecole di glucosio) e
destrine (7-9 unità di glucosio, con presenza di una ramificazione). L'attività delle α-amilasi
salivari si arresta nello stomaco a causa dell'acidità che caratterizza l'ambiente gastrico;
- La digestione dei carboidrati riprende e si completa nell'intestino tenue, grazie all'azione
combinata dei succhi pancreatici ed intestinali. Nei primi, è presente un enzima α-amilasi
pancreatica , che come tale trasforma la miscela di frammenti lineari e ramificati proveniente
dalla bocca in ulteriori maltosio e destrine le quali non possono essere idrolizzate dalle
amilasi pancreatiche e subiscono l'azione di appositi enzimi deramificanti o disaccaridasi
intestinali (α-1,6 glicosidasi, α-destrinasi o isomaltasi) presenti nelle cellule epiteliali
dell'intestino tenue le quali completano la digestione dei di- e tri-saccaridi producendo
monosaccaridi (glucosio, galattosio e fruttosio) che possono essere assorbiti da sistemi di
trasporto specifici siti nelle cellule epiteliali dell’intestino tenue;
- I monosaccaridi prodotti dalla digestione vengono trasportati all’interno delle cellule grazie
alla presenza di specifici trasportatori di membrana.
La Glicolisi
È la via metabolica che degrada il glucosio per ottenere piruvato ed è ubiquitaria negli organismi
viventi; avviene nel citosol (fase solubile nel citoplasma delle cellule). Permette la produzione netta
di 2 molecole di ATP per ogni molecola di glucosio degradata a piruvato e da quest’ultimo può
continuare sia in condizioni aerobie ( ciclo dell’acido citrico ) che anaerobie ( trasformato in lattato ).
La reazione inversa (anche se non ha esattamente gli stessi passaggi) della glicolisi è la
gluconeogenesi che permette la sintesi di glucosio a partire da precursori non saccaridici. La glicolisi
comprende due fasi: fase degli esosi (o fase dell’apporto di energia o fase preparatoria) che coinvolge
molecole a 6 atomi di C le quali sono convertite in un prodotto comune ( gliceraldeide 3-fosfato ) e
la fase dei triosi (o fase del recupero di energia o fase della generazione di energia) che coinvolge
invece molecole a 3 atomi di C ed in essa avvengono le reazioni di ossido-riduzione e i meccanismi
che permettono la conservazione dell’energia attraverso la fosforilazione dell’ADP in ATP. In questa
fase si formano 4 ATP e quindi sottraendo le 2 ATP della fase preparatoria, la resa netta è di 2 ATP
+ 2 piruvato per molecola di glucosio degradata
Fase degli esosi
Reazione 1 : Fosforilazione del D-glucosio da parte dell’ATP; Utilizzando ATP, la molecola neutra
di glucosio viene preparata per le successive tappe enzimatiche mediante la sua fosforilazione e
trasformazione in una molecola carica negativamente. La fosforilazione del glucosio in posizione 6
da parte dell’ATP a glucosio 6-fosfato è catalizzata dall’ esochinasi utilizzando 1 molecola di ATP.
Reazione 6 : Ossidazione della gliceraldeide 3-fosfato a 1,3-bifosfoglicerato; Questa è una delle
reazioni più importanti in quanto essa conserva l’energia derivante dalla ossidazione del gruppo
aldeidico della gliceraldeide 3-fosfato nel suo prodotto di ossidazione 1,3-bifosfoglicerato. La
reazione è catalizzata dalla gliceraldeide 3-fosfato deidrogenasi. In questa reazione il gruppo
aldeidico della gliceraldeide 3-fosfato viene ossidato a gruppo carbossilico.
L’energia sviluppata nella reazione redox viene conservata transitoriamente sotto forma del
composto ad alta energia 1,3-bifosfoglicerato , la cui degradazione nella reazione 7 successiva
permetterà di produrre ATP.
Reazione 7 : Trasferimento di gruppo fosfato da 1,3-bifosfoglicerato a ADP ; L’ 1,3-
bifosfoglicerato formato nella reazione precedente reagisce poi enzimaticamente con ADP con
trasferimento del gruppo acil fosfato all’ADP (si definisce fosforilazione a livello del substrato la
fosforilazione di ADP in ATP all’interno di una via metabolica) e formazione di 3 - fosfoglicerato. La
reazione è catalizzata dalla fosfoglicerato chinasi. N.B.: A questo punto vengono prodotte 2 ATP e
2 NADH per ogni molecola di glucosio (una per ogni GAP prodotta precedentemente).
Importante! Questa modalità di fosforilazione (o sintesi di ATP ) a livello del substrato si
contrappone alla fosforilazione ossidativa (ha una produzione molto più elevata di ATP rispetto alla
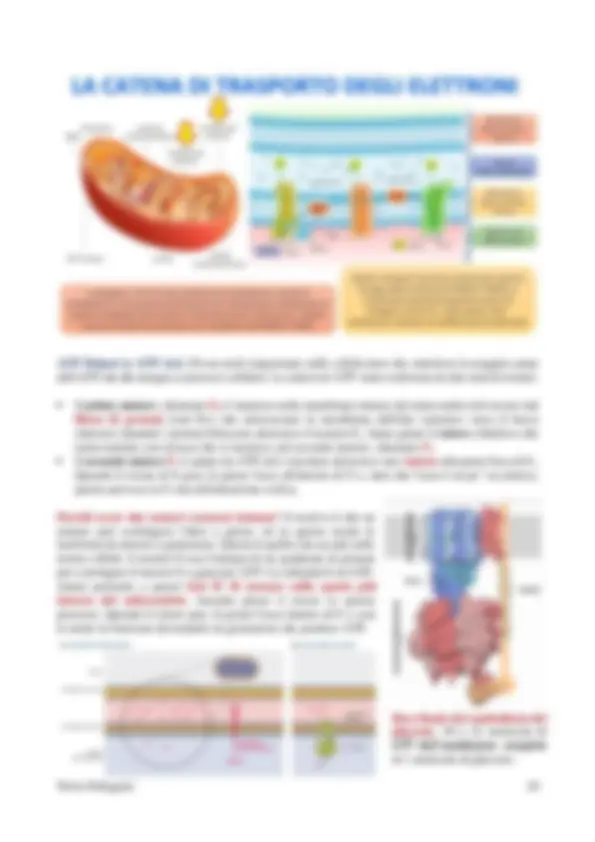
fosforilazione a livello del substrato) che è associata invece alla catena respiratoria.
Reazione 8 : Conversione del 3-fosfoglicerato in 2-fosfoglicerato ; Reazione catalizzata dall’enzima
fosfoglicerato mutasi. Un gruppo fosfato legato all’enzima viene ceduto al C2 di 3PG generando il
2,3 bifosfoglicerato che cede poi il fosfato in C3 all’enzima originando il 2 - fosfoglicerato.
Reazione 9 : Deidratazione di 2-fosfoglicerato a fosfoenolpiruvato ; Si forma un composto
fosforilato ad alto livello energetico (PEP). Essa è catalizzata dall’enzima enolasi.
Reazione 10 : Trasferimento del fosfato da fosfoenolpiruvato a ADP ; Il trasferimento del gruppo
fosforico del fosfoenolpiruvato ad ADP, che porta alla formazione del piruvato libero , è catalizzato
dall’enzima piruvato chinasi. Si tratta di nuovo di una fosforilazione a livello del substrato che
conclude la glicolisi con un bilancio al netto dell’investimento energetico (1 glucosio, 2 NAD+ e 2
ATP) di 2 Piruvato, 2 NADH e 2 ATP (4 totali – 2 investiti).
La regolazione delle vie metaboliche : La PFK1 ( fosfofruttochinasi ) catalizza la terza reazione
della glicolisi, che determina in modo irreversibile l’ingresso del glucosio nella glicolisi (il glucosio
6 - fosfato può prendere altre vie diverse dalla glicolisi). L’ATP, il citrato e i protoni (H
) segnalano
che nella cellula c’è ampia disponibilità di fonti energetiche e infatti inibiscono l’enzima. Al contrario
l’AMP attiva la fosfofruttochinasi in quanto segnala una carenza energetica. In pratica è il rapporto
[ATP]/[AMP] che regola la velocità con cui il glucosio viene inviato alla degradazione.
Insulina controlla ingresso del glucosio nelle cellule : Nell’uomo la concentrazione di glucosio nel
sangue (la glicemia ) è elevata ed è mantenuta a valori costanti in modo tale che lo zucchero possa
prontamente pervenire ai tessuti. L’ingresso del glucosio nelle cellule è dettata dall’interazione
dell’ ormone insulina (prodotta dal pancreas ) con uno specifico recettore della membrana
plasmatica. Nel muscolo scheletrico, nel muscolo cardiaco e nel tessuto adiposo il trasportatore del
glucosio è sequestrato in piccole vescicole e si trasferisce sulla membrana plasmatica in presenza di
insulina (insulino dipendenti). In altri tessuti (cellule del fegato e neuroni) l’assorbimento del glucosio
è insulino indipendente. In caso di diabete la quantità di insulina prodotta non è sufficiente, il
glucosio si accumula nel sangue originando la condizione definita di iperglicemia.
La fermentazione
Termine generale con il quale si indica la degradazione anerobica (cioè in assenza di ossigeno) del
glucosio e di altri nutrienti organici per ottenere energia (sotto forma di ATP). Per questo motivo, la
glicolisi può essere indicata anche con il termine di “demolizione anaerobica del glucosio”. La
mancanza di ossigeno impedisce alla cellula di "sfruttare" a fondo il potenziale energetico del
glucosio e non ottiene l’ATP che è in grado di produrre con ciclo di Krebs e ATP sintasi. La cellula
necessita di cofattori ossidati (NAD
) in grado di ossidare il substrato e continuare la glicolisi. Per
questo motivo gli elettroni del NADH vengono ceduti al piruvato, che ossida il NADH, riducendosi.
Fermentazione alcolica : Non è svolta dai mammiferi ma dai lieviti (es. Saccharomyces cerevisiae).
Le reazione consiste di due passaggi , catalizzati da altrettanti enzimi. L'enzima chiamato piruvato
decarbossilasi (PDC) libera una molecola di anidride carbonica (CO 2
) dal piruvato (in un processo
chiamato decarbossilazione ), ottenendo acetaldeide. Quest'ultima viene ridotta a etanolo
dall'enzima alcol deidrogenasi (ADH), il quale utilizza come cofattore il NADH(H+) ossidandolo
nuovamente a NAD+ e rendendolo nuovamente disponibile per la glicolisi.