Scarica sistema del complemento e più Dispense in PDF di Immunologia solo su Docsity!
Lez. 07-
IL SISTEMA DEL COMPLEMENTO
Nel sangue e nei fluidi extracellulari sono disperse molecole in grado di riconoscere i microrganismi e di promuovere la risposta immunitaria innata, assicurando una difesa precoce contro i patogeni che penetrano nel circolo sanguigno o che hanno localizzazione extracellulare. Tali molecole effettrici funzionano principalmente in due modi:
- Fungono da opsonine legandosi ai microrganismi e favorendone la fagocitosi mediata da macrofagi e neutrofili. Difatti, i fagociti esprimono recettori di membrana per le opsonine che permettono l’internalizzazione del complesso microrganismo-opsonina e la successiva eliminazione dei patogeni ingeriti;
- Dopo aver legato il microrganismo, queste molecole promuovono le risposte infiammatorie richiamando nuovi fagociti nella sede di infezione;
- A volte, tali molecole riescono a provocare la morte diretta del patogeno. Le molecole effettrici solubili rappresentano la controparte umorale dell’immunità innata, esattamente come gli anticorpi lo sono di quella adattativa. I principali componenti di tale immunità innata umorale sono: anticorpi naturali, collectine, pentrassine, ficoline e sistema del complemento. In questa lezione andremo ad approfondire proprio quest’ultima componente. Il SISTEMA DEL COMPLEMENTO comprende numerose proteine plasmatiche che collaborano fra loro con tre obiettivi: opsonizzare i microrganismi; promuovere il reclutamento dei fagociti; uccidere direttamente i microrganismi. L’attivazione di tale sistema consiste in una cascata proteolitica in cui un precursore enzimatico inattivo, definito zimogeno , viene modificato per poter diventare una proteasi attiva che taglia la proteina successiva della cascata inducendone l’attività proteolitica. Questa cascata enzimatica determina l’amplificazione della quantità dei prodotti proteolitici che si generano ad ogni tappa e che svolgono le funzioni effettrici del sistema del complemento. CENNI STORICI : il nome complemento deriva dagli esperimenti svolti da Jules Bordet poco dopo la scoperta degli anticorpi. L’immunologo ha scoperto che se si fosse incubata una miscela di batteri con un siero contenente anticorpi ad una temperatura di circa 37°, i batteri stessi sarebbero stati lisati. Tuttavia, se il siero, prima di essere aggiunto ai batteri, veniva riscaldato a temperature più alte come 56°, perdeva la sua attività litica. Tale perdita non era dovuta alla denaturazione della struttura degli anticorpi, essendo essi termostabili (= nonostante il calore mantengono le loro funzioni), perciò Bordet comprese che nel siero doveva trovarsi un’altra componente termolabile che aiutava (→”complementava”) la lisi batterica da parte degli anticorpi. Il complemento è presente sia nei sieri sanguigni immuni sia in quelli non immuni. Questa osservazione hanno condotto alla conoscenza di molte reazioni immunologiche, alcune studiate dallo stesso Bordet e ancora valide. Le tecniche da lui ideate hanno notevolmente contribuito allo sviluppo dell'immunologia e alla preparazione dei sieri e dei vaccini, che oggi sono la salvezza di milioni di vite umane.
VIE DI ATTIVAZIONE DEL SISTEMA DEL COMPLEMENTO
La prima fase nell’attivazione del sistema del complemento è il riconoscimento di molecole sulle superfici microbiche, le quali sono, però, assenti sulle cellule dell’ospite. Ciò avviene secondo tre modalità identificate come vie del complemento :
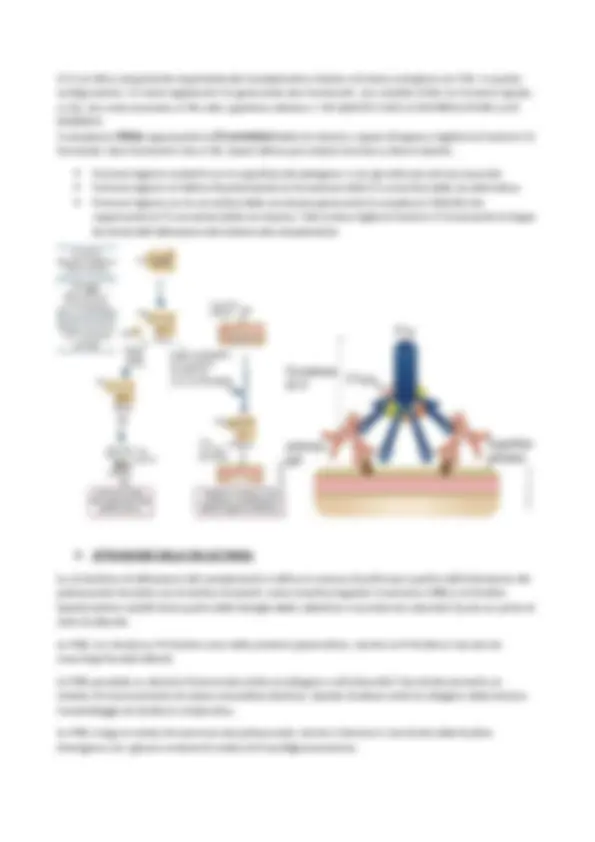
- VIA CLASSICA , così chiamata perché scoperta per prima. Essa utilizza la proteina plasmatica C1q e per identificare gli anticorpi legati alla superficie microbica o ad altre strutture. Quando C1q lega la porzione Fc degli anticorpi, due serinproteasi, C1r e C1s , si attivano e iniziano la cascata proteolitica. Altre proteine solubili dell’immunità innata, le pentrassine, possono legare C1q e dare inizio alla via classica;
- VIA ALTERNATIVA venne scoperta successivamente alla via classica, ma è filogeneticamente più antica di essa. Il processo inizia quando una proteina del complemento chiamata C3 riconosce determinate strutture microbiche superficiali, quali, per esempio, l’LPS batterico. Inoltre, C3 può attivarsi costitutivamente anche a bassi livelli, ma la deposizione sulle cellule self viene inibita da proteine inibitorie. Al contrario, i microrganismi non esprimono tali proteine inibitrici, per cui sulla loro membrana può verificarsi l’attivazione del sistema del complemento. è chiaro, dunque, che questa via è in grado di distinguere le cellule self dai microrganismi;
- VIA LECTINICA inizia con una proteina chiamata lectina legante il mannosio (Mannose-Binding Lectin, MBL ) che riconosce i residui terminali di molecole di mannosio presenti su glicoproteine e glicolipidi microbici. MBL appartiene alla famiglia delle collectine ed è caratterizzata da una struttura esamerica simile a quella di C1q. In seguito al legame di MBL ai microrganismi, due zimogeni, MASP1 e MASP2, si associano ad essa (MBL) e iniziano la cascata proteolitica identica a quella della via classica. Il riconoscimento dei microrganismi da parte di una delle qualsiasi tre vie del complemento porta al reclutamento sequenziale e all’assemblaggio di ulteriori proteine del complemento per formare complessi ad attività proteasica. Ne è un esempio il complesso C3 convertasi, la cui attività comprende la scissione della proteina centrale del complemento C3 in due frammenti, C3a e C3b. Il frammento C3b è il più lungo dei due, e si lega covalentemente alla superficie cellulare del microbo che ha attivato la via del complemento. In questo modo, esso funge da opsonina legandosi covalentemente ai microrganismi e promuovendone la fagocitosi. Invece, il frammento più piccolo c3a viene rilasciato in circolo, dove stimola l’infiammazione agendo come fattore chemotattico per i neutrofili, inducendo la degranulazione dei mastociti e aumentano la permeabilità vascolare, in modo da permettere la fuoriuscita di plasma e proteine plasmatiche nei tessuti sede d’infezione. Il frammento C3b può anche legare altre proteine del complemento, formando con esse un complesso proteasico che viene definito C5 convertasi che scinde la proteina C5 In C5a (piccolo) e C5b (grande). Allo stesso modo di C3b, C5b resta legato alla membrana plasmatica del microrganismo, mentre C5a svolge le stesse funzioni proinfiammatorie di C3a. Inoltre, il frammento C5b innesca la formazione di un altro complesso proteico, costituito da C6, C7, C8 e C9, definito COMPLESSO DI ATTACCO ALLA MEMBRANA (MAC). Quest’ultimo forma pori sulla membrana plasmatica dei microbi che hanno innescato l’attivazione del sistema del complemento, lisandoli.
- L’attivazione del complemento avviene attraverso la proteolisi sequenziale delle sue diverse componenti, che porta all’assemblaggio di complessi enzimatici dotati a loro volta di attività proteolitica. Le proteine inizialmente inattive che poi danno il via alla cascata proteolitica di eventi che porta all’attivazione del complemento prendono il nome di zimogeni. Tale meccanismo permette una rapida amplificazione del processo, poiché ogni singola molecola dell’enzima che viene attivata in ciascuna tappa attiva altre molecole in tappe successive.
- Molti dei prodotti di attivazione del complemento diventano biologicamente attivi in seguito al taglio e si legano covalentemente agli antigeni sulla superficie o agli anticorpi che hanno riconosciuto il microrganismo, nonché ai corpi apoptotici. In fase fluida, le proteine del complemento sono presenti in forma inattiva oppure vengono attivate transitoriamente per frazioni di secondo, lasso di tempo insufficiente a far sì che esse funzionino correttamente. In seguito al legame con microrganismi, con anticorpi associati a microrganismi o con cellule in apoptosi, esse vengono stabilmente attivate. Di conseguenza, è chiaro che le funzioni del sistema del complemento di espletano limitatamente alla superficie del microrganismo o alla sede in cui gli anticorpi hanno legato gli antigeni, evitando che il fenomeno si espanda ad altri distretti.
- I prodotti biologicamente attivi della cascata del complemento stimolano i processi infiammatori. Il reclutamento di neutrofili e monociti genera un ambiente proinfiammatorio che favorisce l’eliminazione dei patogeni.
- L’attivazione del complemento è inibita da proteine espresse sulla membrana delle cellule dell’ospite, ma non sulla superficie dei microrganismi. Queste proteine prevengono o almeno limitano gli effetti lesivi dannosi derivanti dall’attivazione del complemento. Visto che i microrganismi non contengono tali proteine inibitrici, il sistema del complemento distingue cellule self da cellule estranee, e completa la sua attività solo su queste ultime.
VIE DI ATTIVAZIONE DEL COMPLEMENTO
Precedentemente sono state introdotte le tre vie di attivazione del sistema del complemento, le quali differiscono per le modalità con cui vengono avviate, ma portano tutte alla generazione di complessi enzimatici in grado di tagliare C3, la componente centrale e più abbondante del complemento. La via alternativa e quella lectinica sono considerate risposte immunitarie innate, mentre quella classica è un meccanismo effettore dell’immunità umorale adattativa. L’evento principale dell’attivazione del complemento, a prescindere dalla via, è la proteolisi del frammento C3 con la formazione dei prodotti biologicamente attivi, C3b e C3a, grazie al complesso proteolitico della C convertasi. Per convenzione, il frammento più lungo è sempre denominato con la lettera “b”, mentre quello più corto con la lettera “a”. Una volta formato, C3b si lega covalentemente alla superficie del microrganismo o dell’anticorpo ad esso associato. Tutte le funzioni biologiche dipendono dal taglio proteolitico del frammento C3 → es: il sistema del complemento promuove la fagocitosi perché C3b lega covalentemente i microbi, e i fagociti, cioè neutrofili e macrofagi, esprimono i recettori per C3b. Inoltre, i prodotti biologicamente attivi di C3 stimolano l’infiammazione. Inoltre, in tutte e tre le vie, dopo la scissione di C3, si assembla un secondo complesso enzimatico chiamato C5 convertasi, che scinde il frammento C5 in C5a e C5b, contribuendo al processo infiammatorio e alla formazione di pori sulla membrana dei microrganismi.
Dunque, le vie di attivazione differiscono per il modo in cui viene prodotto C3b, ma in seguito alla scissione di C5 seguono tappe comuni.
L’assenza di queste proteine sui microorganismi consente l’ulteriore della C3 convertasi della via alternativa e la progressione dell’attivazione del sistema del complemento. Nella via alternativa, inoltre, interviene una proteina specifica chiamata properdina, la quale viene secreta dai neutrofili attivati, dai macrofagi e da alcuni linfociti T. Essa è capace di legarsi al frammento C3bBb col fine di stabilizzarlo. L’azione della properdina avviene preferenzialmente sulla superficie dei microrganismi e non su quella delle cellule dell’ospite. Tale proteina è l’unico regolatore positivo del sistema del complemento finora conosciuto. Il frammento C3b genarato dalla C3 convertasi della via alternativa può legarsi alla C3 convertasi stessa, portando alla formazione di un complesso contenente un frammento Bb e due molecole C3b. Tale complesso rappresenta la C5 convertasi della via alternativa, tagliando il frammento C5 e innescando le fasi terminali dell’attivazione del complemento.
2. ATTIVAZIONE DELLA VIA CLASSICA
La via classica è avviata dal legame della frazione C1 ai domini CH2 delle IgG o ai domini CH3 delle IgM complessate all’antigene. Le classi di anticorpi IgG1 e IgG3 nell’uomo sono le più efficienti nell’attivare il sistema del complemento. La frazione C1 è un complesso proteico multimerico composto dalle subunità C1q, C1r e C1s. La subunità C1q è deputata al legame con l’anticorpo, mentre C1r e C1s hanno attività proteasica. C1q ha una struttura simile ad un ombrello: presenta catene disposte radialmente, ognuna delle quali ha una testa globulare. Le subunità, poi, sono tutte connesse a uno stelo centrale con struttura simile a quella del collageno. Per portare a termine la sua funzione, C1q lega le regioni Fc delle catene μ e alcune catene pesanti γ. Solo più anticorpi legati ad un antigene multivalente (e NON i singoli anticorpi liberi circolanti) possono innescare l’attivazione della via classica del complemento. Ma perché? Perché ogni regione Fc ha un singolo sito di legame per C1q, mentre tale subunità necessita dell’interazione con almeno due regioni Fc per attivarsi. Dunque, affinché la via classica si attivi, ogni frammento C deve aver accesso ad almeno due o più regioni Fc. Dato che ogni molecola IgG possiede solo una regione Fc, è necessario che esse siano aggregate tra loro prima di legare C1q. Il legame tra più IgG si forma quando esse legano epitopi identici di un antigene multivalente. Le IgM libere e quelle circolanti nel plasma non legano C1q perché in questa configurazione le regioni Fc sono inaccessibili a C1q, di conseguenza è necessaria una modificazione conformazionale per esporre Fc a C1q e farle legare. Inoltre, è bene specificare che le IgM sono più efficaci nel legare C1q, dato che una singola molecola è in grado di legare due C1q. Le subunità C1r e C1s sono serin-proteasi che nel complesso proteico C1 sono organizzate a formare un tetramero contenente due molecole di ciascuna subunità (C1r 2 -C1s 2 ). La subunità C1r viene attivata dal legame di due o più teste della subunità C1q alle regioni Fc delle IgG o delle IgM. A sua volta, C1r attiva C1s mediante taglio proteolitico. C1s agisce sulla componente successiva della cascata del complemento, cioè C4, generando, in seguito a scissione proteolitica, C4a e C4b. C4a viene rilasciata in circolo dove svolge diverse attività biologiche. Il frammento più grande C4b è strutturalmente omologo a C3b, in quando possiede un gruppo tioesterico reattivo capace di formare legami esterici o amidici con la superficie dell’antigene o con l’anticorpo a esso complessato. Il legame covalente di C4b garantisce che l’attivazione della via classica prosegua a stretto contatto con la superficie dell’antigene o dell’immunocomplesso.
La MBL e le ficoline si legano a serin-proteasi associate a MBL, la cui sigla è MASP. Esistono vari tipi di MASP: MASP- MASP-2 → rappresenta la proteasi che taglia le frazioni C4 e C MASP- In generale, le MASP hanno una struttura omologa a quella delle proteasi C1r e C1s e svolgono funzioni simili, ovvero di taglio delle frazioni C4 e C2 per attivare la via del complemento. Gli oligomeri di MBL si associano MASP-1 e MASP-2 o MASP-2 e MASP-3. Gli eventi seguenti sono identici a quelli della via classica.
SEQUENZA TERMINALE DI ATTIVAZIONE DEL COMPLEMENTO
Le C5 convertasi generate durante le tre vie di attivazione del complemento innescano l’attivazione delle componenti terminali, le quali, a loro volta, inducono l’assemblaggio del MAC (Membrane Attack Complex) dotato di attività citolitica. Infatti, le C5 convertasi tagliano il frammento C5 in due parti: C5a e C5b. La prima ha potenti effetti biologici su diversi tipi cellulari, svolgendo la stessa attività proinfiammatoria di C3a; la seconda è costituita da due catene, una α e una β. Una volta liberato C5b, esso va a legare C6 a livello della sua porzione plasmatica. In questo modo, C6 subisce un cambiamento conformazionale e il complesso C5b-C6 si lega alla membrana cellulare mediante interazioni ioniche ed idrofobiche. La frazione plasmatica C7 si lega alla catena α di C5b, formando un complesso più grande denominato C5b- C6-C7. Il C7 complessato subisce una transizione anfifilica e penetra nella membrana, dove concorre al rilascio di alcune micelle fosfolipidiche, senza però formare in essa veri e propri pori. La proteina C8 è un trimero costituito da tre catene distinte, una delle quali lega la frazione C5b del complesso precedentemente formato, e genera un eterodimero, legando in modo covalente la seconda catena; la terza catena si inserisce nel doppio strato fosfolipidico della membrana plasmatica. Qui, il nuovo complesso C5b-C6-C7-C8 forma dei pori instabili → solo un numero molto elevato di questo tipo di complesso è in grado di causare lisi cellulare. C9 è la componente finale della cascata del complemento. Si tratta di una proteina sierica che polimerizza sulla membrana del complesso C5b-8 per formare pori costituiti da complessi C5b-9. Ovviamente, C9 va a legare il complesso C5b-8, generando un MAC completamente attivo. Attraverso i pori possono passare acqua e ioni, e la dimensione del canale che formano varia in base al numero di molecole C9 coinvolte. Inoltre, diverse molecole di C9 possono anche assemblarsi a formare complessi tubulari. L’ingresso di acqua provoca il rigonfiamento osmotico e la lisi della cellula, sulla cui superficie era presente MAC.
RECETTORI PER LE PROTEINE DEL COMPLEMENTO
Molti degli effetti biologici del sistema del complemento sono dovuti al legame dei suoi componenti e/o frammenti biologici con i recettori espressi su vari tipi cellulari.
6. RECETTORI PER I FRAMMENTI C3a, C4a, C5a I recettori per i frammenti C3a, C4a e C5a stimolano l’infiammazione attraverso il reclutamento di macrofagi e mastociti. Il recettore meglio caratterizzato è quello per C5a, localizzato su neutrofili, eosinofili, basofili, monociti, macrofagi, mastociti, cellule endoteliali, cellule muscolari lisce, cellule epiteliali e astrociti. Appartiene alla famiglia di recettori accoppiati a proteine G, esattamente come il recettore per il frammento C3a.
REGOLAZIONE DELL’ATTIVAZIONE DEL COMPLEMENTO
L’attivazione della cascata del complemento e la stabilità dei suoi prodotti bioattivi sono strettamente regolate con due obiettivi: Impedire che l’attivazione avvenga su cellule normali dell’ospite; Limitarne la durata. La regolazione è garantita da diverse proteine circolanti e di membrana, molte delle quali appartengono alla famiglia delle RCA (Regulators of Complement Activity), codificate da geni omologhi adiacenti tra, che si trovano sul cromosoma 1. Le RCA comprendono varie proteine di membrana: DAF/CD55 (Decay Accelerating Factor) MCP/CD46 (Membrane Cofactor Protein) CR1/CD35 (Recettore per il complemento di tipo 1) CR2/CD21 (Recettore per il complemento di tipo 2) CD Le RCA comprendono anche proteine circolanti nel plasma: C1 INH (C1 Inhibitor) Proteina /Fattore 1 Proteina/Fattore H C4BP (C4 -Binding Protein) Ma come mai l’attivazione del sistema del complemento e la sua attività devono essere finemente controllate? Abbiamo detto che il complemento viene costitutivamente attivato in condizioni fisiologiche, per cui la regolazione è necessaria per impedire la sua azione sulle cellule normali dell’ospite e per far sì che, anche quando agisce sui patogeni, non si propaghi sulle cellule circostanti. Ci sono diversi meccanismi che impediscono la formazione della C3 convertasi nelle fasi precoci di attivazione o che la inattivano, così come ne esistono altrettanti per la C5 convertasi e per il complesso MAC.
→ C1 INHIBITOR, C1 INH (inibitore di C1r, C1s e MASP-2)
C1 INH è una proteina della famiglia delle serpine e funge da inibitore delle serin-proteasi. La sua azione si esplica mimando il normale substrato costituito da C1r e C1s, diventando il loro principale bersaglio quando C1q inizia l’attivazione del complemento legandosi a un anticorpo in fase fluida.
C1 INH viene tagliato e si associa covalentemente al tetramero C1r 2 -C1s 2 che si dissocia dalla C1q, bloccando l’attivazione del complemento. In questo modo, l’inibitore previene l’accumulo nel plasma di molecole del complesso C1r 2 -C1s 2 enzimaticamente attive, limitandone la disponibilità per l’attivazione delle tappe successive del complemento. Inoltre, C1 INH ha il compito di inattivare la proteina MASP-2, inibendo la via lectinica del complemento. Un deficit di C1 INH causa la malattia ereditaria autosomico-dominante chiamata angioedema ereditario , caratterizzata da episodi di edema della cute e delle mucose. Alcuni pazienti affetti da tale patologia mostrano livelli plasmatici di C1 INH ridotti del 20/30% rispetto alla norma, valore più che sufficiente affinché l’attivazione di C1 cessi di essere controllata, generando tagli sregolati di C2 e C4.
→ PROTEINE INIBITORIE DELLA FAMIGLIA RCA (inibizione delle componenti di C3 e C
convertasi)
L’assemblaggio delle componenti della C3 convertasi e della C5 convertasi può essere inibito dal legame delle proteine inibitorie della famiglia RCA ai frammenti C3b e C4b associati alla superficie cellulare. Quando il frammento C3b si associa alla superficie di cellule normali dell’ospite, può essere legato da: MCP (CD46); CR1; DAF; fattore H, una proteina plasmatica. Analogamente, C4b può essere legato da: o DAF; o CR1; o MCP; o C4BP (C4 binding protein). Queste proteine impediscono il legame a C3b o C4b degli altri componenti della C3 convertasi, come Bb della via alternativa e C2a della via classica, bloccando la prosecuzione della cascata del complemento. È bene specificare che le proteine MCP, DAF e CR1 sono espresse esclusivamente sulla superficie di cellule di mammifero (cellule ospite), ma non sulla parete dei batteri, per cui impediscono selettivamente l’attivazione del complemento sulle cellule dell’ospite ma non sui patogeni. Un altro meccanismo che impedisce l’azione del complemento sulle cellule dell’ospite è rappresentato dalla presenza di elevate quantità di acido salicilico sulle superfici cellulari dei mammiferi, che favorisce il legame di C3b al fattore H, e non al fattore B.
Tramite molte ricerche effettuate in vitro, è stata delineata una sorta di gerarchia delle proteine che inibiscono l’attivazione del complemento, secondo la quale le proteine più importanti sarebbero CD59, DAF e MCP. È probabile che la loro importanza rifletta le relative concentrazioni delle proteine sulla superficie delle cellule. Un aumento esponenziale dell’attivazione del complemento rende inefficace l’azione delle proteine regolatore. Difatti, nonostante l’efficienza dei meccanismi regolatori dell’attivazione del complemento, la fagocitosi mediata da tale sistema alle cellule ospite contribuiscono alla patogenesi di malattie su base immunologica. Ciò avviene perché elevate quantità di anticorpi si depositano sulle cellule ospite, generando un numero così grande di frammenti del complemento da annullare l’efficienza dei meccanismi di regolazione.
FUNZIONI E DEFICIT DEL COMPLEMENTO
FUNZIONI DEL COMPLEMENTO
Le principali funzioni svolte dal sistema del complemento sono: Promuovere la fagocitosi; Promuovere la lisi dei microrganismi; Stimolare l’infiammazione. Inoltre, i prodotti di attivazione del complemento forniscono il segnale accessorio per l’attivazione dei linfociti B e la produzione di anticorpi. La fagocitosi, l’infiammazione e l’attivazione dell’immunità umorale (produzione anticorpi) sono tutte funzioni regolate dal legame dei frammenti del complemento a diversi recettori di superficie, mentre la lisi cellulare è dovuta al MAC.
- OPSONIZZAZIONE E FAGOCITOSI I microrganismi che attivano il complemento vengono opsonizzati dai frammenti C3b, iC3b e C4b e vengono, poi, fagocitati tramite l’interazione di queste proteine con recettori specifici espressi dai macrofagi e dai neutrofili.
La via alternativa e quella classica portano alla generazione dei frammenti C3b e iC3b e al loro legame alla superficie cellulare. Qui, questi frammenti agiscono da opsonine che vengono riconosciute dai recettori presenti sulla superficie dei neutrofili e macrofagi. Il frammento C4b è generato solo dalla via classica, mentre C3b sia dalla via classica che da quella alternativa. Insieme, tali frammenti legano CR1, mentre iC3b lega CR3 (Mac-1) e CR4. Il solo legame di C3b e C4b a CR1 non è sufficiente a promuovere la fagocitosi dei microrganismi: al contrario, ciò avviene solo se questi vengono anche opsonizzati dalle IgG che si legano ai recettori Fcγ espressi dai fagociti. Anche l’attivazione dei macrofagi mediata da IFN-γ potenzia la fagocitosi di CR1. La fagocitosi dei microrganismi da parte di C3b e iC3b è uno dei principali meccanismi di difesa contro le infezioni sia nell’immunità innata che specifica. L’importanza del complemento come difesa dell’ospite è dimostrata dalle risposte contro batteri dotati di una capsula ricca di polisaccaridi che avvengono essenzialmente con meccanismi umorali, Infatti, le IgM dirette contro i polisaccaridi della capsula si legano ai batteri, attivando la via classica e causando la loro eliminazione mediante fagocitosi a livello splenico.
2. ATTIVAZIONE DELLA RISPOSTA INFIAMMATORIA I frammenti C3a, C4a e C5a attivano l’infiammazione acuta agendo su mastociti, neutrofili e cellule endoteliali. Mastociti → i frammenti che vi si legano, ne inducono la degranulazione con conseguente liberazione di mediatori vasoattivi, come l’istamina. Queste proteine sono dette anafilotossine perché la reazione che scatenano a livello dei mastociti sono simili al processo di anafilassi. Neutrofili → C5a stimola la motilità e l’adesione stabile all’endotelio, oltre che il burst ossidativo che provoca la produzione di ROS. Cellule endoteliali → il C5a aumenta la permeabilità vascolare e l’espressione di P-selectina, che favorisce l’adesione dei neutrofili alle cellule endoteliali stesse.
Inoltre, gli immunocomplessi associati al frammento C3b si legano a CR1, favorendone l’eliminazione mediante il trasporto ai fagociti epatici. La proteina C3d, che deriva dal taglio della frazione C3, si lega a CR2 espresso dai linfociti B, attivandoli e fornendo loro un segnale per l’avvio della risposta umorale. Il frammento C3d si genera quado il complemento è attivato direttamente da un antigene o quando l’anticorpo riconosce l’antigene. L’attivazione del complemento porta al legame covalente del frammento C3b e del suo prodotto di degradazione C3d all’antigene I linfociti B , così , possono legarsi all’ antigene sia attraverso le loro Ig di membrana, sia attraverso le loro Ig, sia con CR2 che funge da corecettore rendendo più efficente la trasduzione del segnale. Inoltre, nei centri germinativi degli organi linfoidi secondari, gli antigeni opsonizzati possono essere catturati dalle cellule dendritiche follicolari, che espongono tali antigeni ai linfociti B (è importante per la selezione dei linfociti B ad elevata affinità all’antigene). Il ruolo fondamentale del complemento nelle risposte immunitarie umorali è evidenziato dalle gravi alterazioni dello sviluppo dei centri germinativi e della produzione di anticorpi osservati nei topi knockout per geni che codificano per le frazioni C3 e C4 e per il recettore CR2.
DEFICIT DEL COMPLEMENTO
Difetti genetici nella sintesi delle proteine del complemento e delle proteine regolatorie sono la causa di varie malattie. Tra i deficit a carico delle componenti della via classica (C1q, C1r, C4,C2 e C3), quello più riscontrato è a carico di C2. Inoltre, più del 50% dei pazienti sviluppa una sindrome autoimmune simile al lupus eritematoso sistemico. Le ragioni che associano questi deficit a patologie autoimmuni da immunocomplessi sono sconosciute, ma si suppone che questi deficit possono comportare un’insufficiente eliminazione degli immunocomplessi circolanti. Infatti, se non adeguatamente rimossi, gli immunocomplessi tendono a depositarsi sulle pareti dei vasi e nei tessuti, dove, legandosi agli FcR, attivano i leucociti innescando il processo infiammatorio. Il sistema del complemento concorre anche all’eliminazione dei corpi apoptotici contenenti il DNA frammentato. Questo materiale di degradazione cellulare è una delle probabili fonti di antigeni nucleari che scatenano la risposta anticorporale autoreattiva nel lupus. Bisogna anche ricordare che alcune proteine del complemento regolano la trasduzione del segnale del recettore per l’antigene nei linfociti B: in assenza di questa regolazione può verificarsi un difetto di tolleranza dei linfociti B, con conseguente innesco di una risposta autoimmunitaria. Pazienti con deficit delle frazioni C2 o C4 reagiscono in due modi differenti: in alcuni casi mostrano un’aumentata suscettibilità alle infezioni, mentre in altri sono del tutto asintomatici. Il Deficit della C3 causa infezioni gravi e ricorrenti da batteri piogeni spesso fatali, evidenziando l’importante ruolo che questa componente svolge nei fenomeni di opsonizzazione, fagocitosi ed eliminazione di questi batteri.
DEFICIT DELLE COMPONENTI DELLA VIA ALTERNATIVA
Tali deficit causano un’aumentata suscettibilità a infezioni meningococciche. Mancanze a carico dei fattori B e D sono rare, al contrario di quelle legate alla properdina. Queste ultime si verificano su un gene presente sul cromosoma X trasmesso con carattere recessivo. Mutazioni dei geni codificanti per la lectina MBL e per la MASP-2 contribuiscono ad alcune immunodeficienze.
DEFICIT DELLE COMPONENTI DELLA SEQUENZA TERMINALE
Questi deficit sono stati descritti a carico delle frazioni C5, C6, C7, C8 e C9, e causano una tendenza a sviluppare infezioni da Neisseria. Ciò dimostra come la lisi mediata dal complemento sia cruciale per la difesa contro i microrganismi patogeni che possiedono una parete molto sottile.
DEFICIT DELLE PROTEINE REGOLATRICI DEL COMPLEMENTO
Questi deficit causano un’attivazione incontrollata del complemento, con conseguenti alterazioni cliniche. Deficit della proteina DAF (vedi sopra); Deficit dell’i nibitore di C1 (vedi sopra); Deficit del fattore I determina un consumo aberrante della frazione C3 con formazione incontrollata della C3 convertasi in fase fluida. La sua conseguenza clinica consiste nell’aumento della suscettibilità alle infezioni da batteri piogeni. Deficit del fattore H sono estremamente rari e causano un eccesso di attivazione della via alterativa e quindi un eccessivo consumo di C3. Le manifestazioni cliniche consistono in glomerulonefriti generate dall’insufficiente eliminazione degli immunocomplessi che vanno a depositarsi a livello renale. È stata descritta una forma della sindrome uremica-emolitica associata a deficit delle proteine regolatorie, dovuta per lo più alla mutazione del gene del Fattore H e del gene MCP. In questa patologia, i bambini presentano anemia emolitica microangiopatica, trombocitopenia, insufficienza renale acuta, tutte manifestazioni dovute a danno endoteliale causato dall'attivazione della via alternativa del complemento. Inoltre, specifiche mutazioni a carico del Fattore H o MCP portano a un legame meno efficientemente di C3b e C4b sulle superfici endoteliali, generando un'incontrollata attivazione del complemento che determina la formazione di microtrombi e danno vascolare. Le manifestazioni cliniche dei deficit del Fattore I o del Fattore H sono simili a quelle causate da un autoanticorpo definito fattore nefritogeno (C3NeF) diretto contro la C3 convertasi della via alternativa (C3bBb). Il C3NeF stabilizza il complesso C3bBb, proteggendolo dalla dissociazione favorita dal Fattore H, causando un eccessivo consumo della frazione C3. I pazienti che presentano in circolo tali anticorpi sviluppano spesso glomerulonefriti, causate dall'inadeguata eliminazione degli immunocomplessi circolanti e depositati a livello renale. Specifiche varianti alleliche del Fattore H sono associate alla degenerazione maculare correlata all'età: l'eccessiva infiammazione in assenza di regolazione del complemento contribuisce alla distruzione dei fotorecettori della regione maculare con conseguente cecità.