¡Descarga beta oxidación, estudio general y más Apuntes en PDF de Bioquímica solo en Docsity!
UNIVERSIDAD DE SAN CARLOS DE GUATEMALA
FACULTAD DE ODONTOLOGÍA
BIOQUÍMICA
OXIDACIÓN DE ACIDOS GRASOS
Los ácidos grasos son utilizados en nuestro cuerpo a través de la vía metabólica denominada β oxidación, esta vía es de suma importancia debido a que la mayoría de tejidos de nuestro cuerpo, utilizan con frecuencia los ácidos grasos provenientes de los triglicéridos, guardados en el tejido adiposo, como fuente energética. a. GENERALIDADES
- Los depósitos de TAG (triacilglicéroles) del tejido adiposo, sirven como la mayor reserva de combustibles del organismo y tanto el glicerol como los ácidos grasos, pueden ser utilizados para producción de energía por diferentes vías metabólicas.
- Los ácidos grasos son fácilmente movilizados para proporcionar energía, cuando existe ejercicio extremo o ayuno prolongado, y son ricos en calorías, debido a que contiene un gran número de átomos de hidrógeno, favoreciendo así la formación de ATP.
- La oxidación de los ácidos grasos producen alrededor de 9Kcal/g, comparado con 4 Kcal/g que producen las proteínas y los carbohidratos.
- Los AGL (ácidos grasos libres) pueden ser degradados para obtener energía o bien para ser utilizados en la biosíntesis de diversos lípidos. Ej. El ácido araquidónico (poli insaturado) puede usarse para síntesis de prostaglandinas y leucotrienos.
- Las enfermedades relacionadas con el deterioro de la oxidación de AG, conducen a hipoglucemia, infiltración grasa de órganos e hipocetonemia.
- La alta degradación de AG (ácidos grasos), puede conducir a la acumulación de moléculas de acetil Co-A y estas al mismo tiempo puede generar cuerpos cetónicos, que son moléculas ácidas y en gran cantidad producen cetosis, ésta es leve en la inanición, pero grave en la diabetes sacarina no tratada.
- La oxidación de ácidos grasos es un proceso aerobio, que exige la presencia de oxígeno. ANÀLISIS Después de haber analizado las generalidades y con los conocimientos adquiridos de otros metabolismos, responda las siguientes preguntas:
- Durante un ayuno prolongado ¿Qué sucede con las siguientes vías metabólicas? ¿se activan o se inhiben?: a. Síntesis de TG b. Síntesis de Proteìnas c. Degradación de TG (Lipólisis) d. Degradación de Proteínas (Proteólisis) e. Gluconeogénesis f. Glucogenólisis g. Glucogènesis
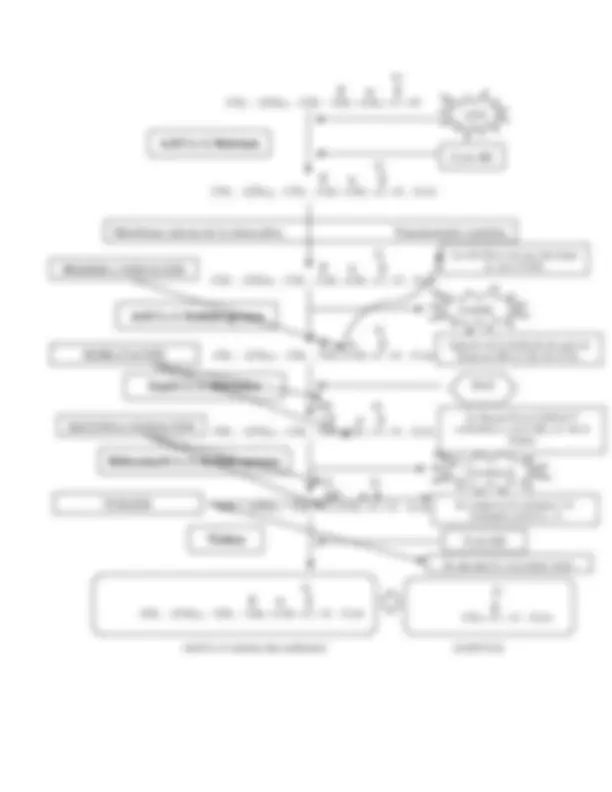
b. MOVILIZACIÓN DE LÍPIDOS: R AC H Glicerol cAMP ATP PK TAGL
Aco.A
ATP
CO 2
Albúmina
T
T
GRÁFICA No. 1 Movilización de lípidos. La movilización de los lípidos desde el tejido adiposo hasta el hígado, se encuentra mediado por un proceso hormonal, en donde el glucagón y la epinefrina, favorece el movimiento de ácidos grasos y la insulina lo inhibe. También la norepinefrina secretada por el sistema nervioso simpático en el tejido adiposo, favorece esta vía. El proceso se realiza permitiendo la formación de segundos mensajeros, como lo es el AMPc, los pasos implica:
- Primero: Unión de la hormona al receptor de membrana, este utilizando proteína G, permite que se active el adenilato ciclasa.
- Segundo: El adenilato ciclasa utilizando una molécula de ATP, favorece la formación de AMPc
- Tercero: El AMPc, activa la proteín quinasa, para que pueda inducir la reacción de una primera enzima que es la Triacilglicerol lipasa, esta enzima es la encargada de la ruptura de los TAG y por consiguiente la liberación de ácidos grasos, para que puedan
De manera que, si tengo un ácido grado de 12 carbonos, cuyo nombre común es el láurico, al momento de oxidación completa, se obtienen 6 moléculas de acetil- Co A. Pregunto: Si tengo un ácido mirístico y palmítico ¿Cuántas moléculas de acetil- Co A se obtienen cuando se oxida de forma completa el ácido graso? Antes de que este proceso de inicio, los ácidos grasos son activados por esterificación con CoA y la enzima tioquinasa, (este tema no se tratará a detalle, pues ya se impartió cuando se analizó la síntesis de lipoproteínas) Después de activa las moléculas los ácidos grasos son transportados a la matriz mitocondrial por una lanzadera de carnitina, ya adentro de la mitocondria, son oxidados por una serie de reacciones que producen energía y el producto que es acetil Co A, que entra al ciclo de Krebs para producción de energía en forma de ATP y Electrones. d. LANZADERA DE LA CARNITINA La carnitina es una proteína conformada por lys y met, sirve como acelerador para la obtención de energía a través de la degradación de ácidos grasos; posee dos estructuras espaciales, de las cuales la forma L es la biológicamente activa En el caso de la oxidación de ácidos grasos, esta molécula es necesaria para introducirlos y ser degradados en la matriz mitocondrial. Para ingresar a la mitocondria, los ácidos grasos ya activados en forma de acil Co-A, pueden atravesar la membrana externa, pero no así la membrana interna, entonces para poder acceder al interior, se sustituye la Coenzima A por una proteína llamada Carnitina, y poder utilizar la proteína transportadora específica que permitirá el ingreso de la molécula a la matriz mitocondrial. La enzima de nombre Carnitina acil transferasa I, que se encuentra en la membrana externa, favorece la sustitución de la coenzima A del ácido grados por una molécula de carnitina, generando como producto la molécula acil-carnitina. La acil-carnitina, puede atravesar la membrana interna, por medio de una proteína transportadora integral de nombre carnitina-acilcarnitina translocasa y así acceder a la matriz mitocondrial. Se hace la acotación, que únicamente puede ingresar a la matriz por medio de la proteína translocasa, no puede utilizar otra vía
ANALISIS
1. ¿Por qué se necesitan enzimas del citosol y de la mitocondria de la célula para
que los ácidos grasos se puedan utilizar como fuente de energía?
2. ¿Por qué los ácidos grasos deben ser activados, antes de que el proceso de β-
oxidación de inicio? (este tema ya lo tratamos en metabolismo de lipoproteínas.
3. ¿Por qué no se realiza β-oxidación, en todos los tejidos del cuerpo?
Ya en el interior de la matriz, la molécula acil-carnitina, no puede ser degradada de esta forma, para ello necesita que se le sustituya otra vez la molécula carnitina por una coenzima A, entonces la enzima carnitina acil transferasa II, elimina la molécula de carnitina y coloca una molécula de Coenzima A, para obtener otra vez el producto Acil-Co A y poder proceder al proceso de β-oxidación. Esquema de repaso Membrana Externa Membrana Interna
- Acil
- Co-A
- Carnitina acil transferasa I
- Carnitina
- Carnitina-acilcarnitina-translocasa
- Carnitina acil transferasa II e. ETAPAS DE LA β-OXIDACIÓN. Se le llama β- oxidación, ya que el proceso oxidativo de realiza entre el carbono α y el carbono β de la molécula. Es un ciclo de 4 reacciones y cada cuadro reacciones, se obtiene como producto una molécula de acetil co-A y un grupo acilo con 2 carbonos menos. Ejemplo: Suponiendo que tenemos un ácido graso de 14 carbonos, O β α CH 3 -- (CH 2 ) 9 -- CH 2 -- CH 2 – CH 2 – C – OH CH 2 – C –
Sustituye Co- A por carnitina.
Sustituye carnitina por Co-A Transporta
Atraviesa 2
O
CH 3 -- (CH 2 ) 7 -- CH 2 -- CH 2 – CH 2 – C – OH
Co-A
O
Acil Co-A menos 2 carbonos (ácido graso de 12 carbonos) Molécula de acetil Co- A (molécula de 2 carbonos)
1. Degradación (β-oxidación): ACIDOS GRASOS SATURADOS DE CADENA PAR a. Las moléculas de acil CoA dentro de la matriz mitocondrial sufren β-oxidación, que es una secuencia cíclica de cuatro reacciones: b. OXIDACIÓN: La oxidación del acil-CoA introduce un doble enlace entre los átomos del C2 y C3 (carbono α y β respectivamente), con la reducción de una molécula de FAD a FADH 2 , que entra en la cadena transportadora de electrones para producir 2 ATP. La enzima que cataliza esta reacción es un Acil-Co A deshidrogenasa. Esta reacción se acopla a la reacción del succinato del ciclo de Krebs Pregunto ¿Qué efecto genera este mecanismos de acoplarse al ciclo de Krebs? c. HIDRATACIÓN: Es la adición de agua a través del doble enlace entre C2 y C mediante la enoil-CoA-hidratasa específicamente para cada ácido graso. Esta reacción, se acopla a la fumarasa en el ciclo de Krebs. d. OXIDACIÓN La 3 - hidroxiacil-CoA deshidrogenasa, es oxidada por una reacción dependiente de NADH para producir 3 - cetoacil CoA, con la consecuente reducción de NAD a NADH+H+, que entra en la cadena transportadora de electrones para producir 3.5 ATP. e. TIOLISIS: La enzima tiolasa rompe la molécula de acil-CoA para liberar acetil- CoA y Acil-CoA, El proceso consiste en cortar en dos carbonos y la molécula restante, queda lista para sufrir otra ronda de β-oxidación, en donde se repiten las cuatro reacciones y se libera otra molécula de acetil Co A. El último paso de la oxidación produce dos moléculas de acetil- Co A.
ANALISIS
La β-oxidación se encuentra estrechamente relacionada con el Ciclo de Krebs
1. ¿Qué relación se infiere en la primera oxidación?
2. ¿Qué relación se infiere en la segunda reacción?
O
β α CH 3 -- (CH 2 )n -- CH 2 -- CH 2 – CH 2 – C – O- O β α CH 3 -- (CH 2 )n -- CH 2 -- CH 2 – CH 2 – C – S -- CoA O β α CH 3 -- (CH 2 )n -- CH 2 -- CH 2 – CH 2 – C – S -- CoA O β α CH 3 -- (CH 2 )n -- CH 2 -- CH 2 = CH – C – S -- CoA OH O β α CH 3 -- (CH 2 )n -- CH 2 -- CH – CH 2 – C – S -- CoA O O β α CH 3 -- (CH 2 )n -- CH 2 -- C—CH 2 – C – S -- CoA Acil-Co A (menos dos carbonos) Acetil-CoA
ATP
CoA-SH
Acil Co A Sintetasa
Membrana interna de la mitocodria Transportador carnitina
Acil Co A Deshidrogenasa^ FADH^2
Enoil Co A Hidratasa^ H 2 O
NADH+H
Hidroxiacil Co A Deshidrogenasa
Tiolasa CoA-SH
O
β α CH 3 -- (CH 2 )n -- CH 2 -- CH 2 – CH 2 – C – S -- CoA
O
CH 2 – C – S -- CoA
PRIMERA OXIDACIÓN
Los H 2 libres con sus electrones se van a FAD+
HIDRATACIÓN
Ingresos de la molécula de agua en forma de OH al C β e H al Cα.
SEGUNDA OXIDACIÓN
Se liberan H en el OH del C carbonílico y en el Cβ y se van al NAD+
TIOLISIS Se rompe en los primeros 2 C
formando acetil Co- A Se une una Co-A al grupo Acilo.
g. OXIDACIÒN DE ÁCIDOS GRASOS DE CADENA IMPAR: La mayoría de los ácidos grasos de la naturaleza posee cadena par de átomos de carbono, sin embargo la oxidación de ácidos grasos de cadena impar es en esencial igual que para los de número par, excepto porque la última β-oxidación produce una molécula de acetil Co-A y una molécula de 3 átomos de carbono llamado propioni-CoA. Molécula que es metabolizada a succinil co A, este último compuesto es un metabolito del ciclo de Krebs. El propionil co A es la porción del ácido graso de cadena impar que es capaz de producir glucosa por gluconeogénesis por lo que se le llama parte glucogénica. h. OXIDACIÒN DE ÀCIDOS GRASOS INSATURADOS: Los ácidos grasos poli-insaturados son muy abundantes y comunes en la naturaleza y durante la oxidación de ácidos grasos insaturados la mayoría de las reacciones son las mismas que para los saturados, excepto en la insaturación ya que cuando es trans intervienen dos enzimas adicionales, la enoil co A-isomerasa, que permite que la estructura se vuelva en conformación cis y la 2,4-dienoil- reductasa que permite que la molécula se satura y se degrade de igual forma que los demás. i. REGULACIÒN DE LA β-OXIDACIÓN: Cuando se están oxidando cantidades importantes de ácido grasos, no se utiliza glucosa como fuente de acetil-CoA, pues el ciclo de Krebs lo está recibiendo de la β-oxidación. Para evitar la conversión de glucosa a acetil-CoA, se inhibe la piruvato deshidrogenasa por la acil carnitina y la acetil CoA, provenientes de la β-oxidación. En la mayoría de las células, la oxidación de los ácidos grasos se controla por la cantidad de sustratos y esta disponibilidad a su vez, se controla mediante las hormonas para permitir movilización de las grasas en los adipocitos. Dado que la función del tejido adiposo consiste en almacenar las grasas para que sea utilizada en otras células, tiene sentido desde el punto de vista metabólico, que la degradación y la liberación de esta grasa almacenada se regule por hormonas, que son mensajeros extracelulares. La acción del glucagón o de la adrenalina, causa la degradación y liberación de las grasas, que da lugar en última instancia a una acumulación de ácidos grasos en otras células. En el hígado, la malonil-CoA constituye un mecanismo regulador que controla la captación de acil-CoA, desde el citosol a la mitocondria, así pues, la malonil-Co A activa la biosíntesis de los ácidos grasos e inhibe su oxidación, al impedir el transporte a la mitocondria. Sería un derroche sintetizar ácidos grasos en el hígado y al mismo tiempo estarlos degradando. Por tanto cuando el exceso de glucosa se transforma en ácidos grasos, se inhibe la β-oxidación por acción de la malonil- CoA, un intermediario de la síntesis de ácidos grasos.
Identifique el rendimiento de ATP del ácido esteárico y palmítico
ANALISIS
1. ¿Quién produce más energía: un àcido graso de cadena impar o uno de cadena
par? Explique
2. ¿Quién produce más energía: un ácido graso saturado o un ácido graso
insaturado? Explique
f. REGULACIÒN DE LA GLUCÓLISIS Y LA β-OXIDACIÒN g. CETOGENESIS Cuando el cuerpo excede la cantidad de acetil-CoA frente a oxalacetato, la mitocondria hepática inicia la formación de algo llamado cuerpos cetónicos. Los compuestos clasificados como cuerpos cetònico son
- Acetona, acetoacetato y β-hidroxibutirato El primer cuerpo cetónico, es un gas que a temperatura del organismo, llega a su punto de ebullición y se mantiene libre en el organismo, esto se caracteriza por un olor descrito a manzana. El segundo y el tercer cuerpo cetónico son los funcionales, estos se pueden transportar por la sangre a los tejidos periféricos, pudiéndose convertir en Acetil-CoA que se oxida en el ciclo de Krebs. Los cuerpos cetónicos pueden convertirse en fuentes energéticas para los tejidos periféricos porque:
- Son solubles en solución acuosa
- Se presentan cuando el acetil-Co A supera la capacidad oxidativa del cuerpo.
- El mùsculo esquelético, cardiaco y la corteza suprarrenal, los utilizan en función de la concentración de sangre. Incluso el cerebro los puede utilizar para satisfacer su necesidad energética y ahorran glucosa. Esto es importante en el período prolongado de de ayuno. h. SINTESIS DE CUERPOS CETÓNICOS Durante el ayuno, inanición, diabetes o consumo excesivo de grasas, el hígado recibe una avalancha de ácidos grasos movilizados por el tejido adiposo. Los elevados niveles de Acetil-CoA hepática resultante, se producen principalmene por la degradación de ácidos grasos, inhiben la piruvato deshidrogenasa y activan la piruvato carboxilasa. El oxalacetato se usa para gluconeogénesis en lugar de para el ciclo de Krebs de manera que la acetil-CoA se aprovecha para la síntesis de cuerpos cetónicos. El proceso metabólico para la activación o inhibición de cuerpos cetónicos implica:
- La oxidación de ácidos grasos reduce el cociente NAD+/NADH
- El aumento de NADH desplaza desplaza el oxalacetato hacia malato. Esto aleja la acetil CoA de la gluconeogénesis y la dirige a la cetogénesis.
Oxidación de Ácidos grasos
Ácido graso
Acil-CoA
Acil-Carnitina
Acetil Co-A
Β-Oxidación
Glucosa
Piruvato
Complejo PDH
i. CETÓLISIS Aunque el hígado sintetiza constantemente niveles bajos de cuerpos cetònicos, su producción se vuelve más significativa en el ayuno, cuando se necesitan cuerpos cetònicos para suministrar energía a los tejidos periféricos Los tejidos extrahepáticos, entre ellos el cerebro, pero no las células que carecen de mitocondria (ej. Eritrocitos), oxidan con eficacia el acetoacetato y el Β-hidroxibutirato. Por el contrario, el hígado aunque produce activamente cuerpos cetónicos no posee una enzima que se llama tioforasa, por lo tanto, es incapaz de utilizar cuerpos cetònicos como combustible.
j. CUERPOS CETÒNICOS EN LA DIABETES Cuando la velocidad de formación de los cuerpos cetònicos es mayor que la de su utilización, sus niveles comienzan a subir en sangre (cetonemia) y finalmente también en la orina (cetonuria). Esto se observa más a menudo en casos de Diabetes Mellitus tipo I no controlada. En las personas diabéticas con cetosis interna, la excreción urinaria de cuerpos cetònicos puede ascender. Un síntoma frecuente de cetoacidosis diabética es un alimento de olor afrutado que proviene de la mayor producción de acetona. El aumento de la concentración de cuerpos cetónicos en la sangre provoca acidosis o academia, que por ser medida por cuerpos cetónicos se llama cetosis Cetosis Diabética Inanición^ Alcohol Tipo I Dependiente de insulina 48 horas Sistema enzimático CAT Lipólisis Y cetogénesis Crónico ß-hidroxibutirato k. TEORIA BIHORMONAL Es una relación que se obtiene cuando la insulina y glucagón afectan el organismo.
Los transportadores Glut 2 se encuentran en las células más sensibles a la glucosa,
como lo son las células β de los islotes de Langerhans (Figura No. 4).
Figura No. 4 Células beta de los Islotes de Langerhans. Tomado de http://dtc.ucsf.edu/es/tipos-de-diabetes/diabetes-tipo-1/comprension-de-la-diabetes-tipo-1/como- controla-el-azucar-el-cuerpo/control-del-azucar-en-sangre/
Dentro de la estructura de las células β, estas poseen un dominio glucocinasa (Un
dominio, es una región proteica que posee una función específica). Este dominio posee
función enzimática, cuya función es semejante a un policía, ya que detecta cuando aumenta
la concentración de glucosa en sangre, permitiendo que ingrese glucosa a la célula β y
aumenta la concentración de calcio, para activar el catabolismo y este mecanismo permite
que se generen monedas energéticas conocida como ATP (adenosin trifosfato).
Al aumentar el ATP, los canales de potasio de la membrana se cierran, esto genera
despolarización de la membrana y activación de los canales de calcio. Este efecto induce a
que se liberen vesículas con insulina hacia el torrente sanguíneo, para que identifiquen
receptores e inicien el proceso metabólico en otras células diana (Figura No. 5).
Figura No. 5 Proceso de liberación de vesículas con Insulina a. Ingreso de Glucosa a la Célula β. b. Aumento de concentración de calcio, para formar ATP. c. Despolarización de la membrana por aumento de K+ y activación de canal de calcio para que ingrese más calcio y d. liberación de vesículas con insulina al torrente para buscar células diana. Tomado de http://commons.wikimedia.org/wiki/File:Glucose_Insulin_Release_Pancreas.svg
En la célula diana, el transportador GluT 4, se regula bajo el efecto de la insulina,
permitiendo el reclutamiento de transportadores desde la parte interna de la célula, hacia la
parte externa y así ingresar glucosa al hígado, músculo y tejido adiposo.
Ahora que ya se analizaron las características normales, describamos que efecto tiene
la Diabetes sobre el ingreso de glucosa a la célula.
Es importante recordar, que la diabetes se caracteriza por: carencia de insulina
causada por ataque autoinmune, disminución del número de islotes β de Langerhans en el
páncreas, resistencia a la insulina en las células diana, obesidad o cambios genéticos en la
GluT 2, causando diversos síntomas que van desde poliuria hasta falta de visibiliad.
Estos síntomas pueden aparecer cuando se han destruido entre el 80 y 90 % de las
células, es por ello que el páncreas no responde; por consiguiente, la insulina no es secretada
y no cumple con sus funciones específicas.
Si recordamos la forma normal como ingresa la glucosa a la célula, los
transportadores que permiten el ingreso hacia adentro de la célula se llaman GluT y en la
diabetes, dos son los importantes, primero el GluT 2, por la regulación de la insulina y
segundo el GluT 4, por la ubicación sistémica en la que se encuentran.
Por lo tanto, si se disminuye el porcentaje de las células β de los Islotes de
Langerhans, también disminuye la presencia GluT 2, que es nuestro sensor de glucosa, no
genera ningún efecto y la glucosa simplemente aumenta en la sangre.
a
b
c. c. d.
que se llama tioforasa, por lo tanto, es incapaz de utilizar cuerpos cetònicos como
combustible.
La cetogénesis (nombre que se le da a la vía metabólica que sintetiza los cuerpos
cetónicos), produce tres moléculas, de las cuales dos son utilizables en el organismo y una
tercera que por su bajo punto de ebullición, rápido se evapora, se vuelve gas, por lo que se
expele aliento afrutado muy semejante a manzana, estas moléculas son: la acetona (producto
secundario no metabolizado) y los metabolizables: el acetoacetato y 3-hidroxibutirato o
también llamado β-hidroxibutirato.
Según Vasudeván “En una persona sana, los niveles de cuerpos cetónicos no excede
1 mg/dl en sangre, sin embargo en una persona diabética no controlada, excede el miligramo
por decilitro y se acumulan en la sangre, generando cetonemia. El cuerpo trata de normalizar
esta patología favoreciendo la excreción de estos por la orina (cetonuria). Estas tres
condiciones juntas (cetogénesis, cetonemia y cetonuria) constituyen la condición conocida
como cetoacidosis” (Figura No. 8).
Figura No. 8 Tomada del Libro de Bioquìmica de Harper, específica cuales son las formas de metabolizar los cuerpos cetónicos, indicando la cetogenesis en el hígado, su desdoblamiento en los órganos para generar acetil Co-A y su eliminación por la orina o pulmones, pudiendo identificar cetosis
La cetoacidosis es un estado patológico que se relaciona con la acumulación de cuerpos
cetonicos en la sangre, esto provoca que disminuya el pH de la sangre menor de 7.35, por lo
que genera acidosis metabólica, provocando taquipnea profunda para poder expulsar CO 2 ,
deprime en general la actividad mental y puede llegar a ser mortal.
A manera de resumen, las ideas generales tratadas hasta el momento implican: Primero
falta de insulina; Segundo no ingreso de glucosa a la célula y acumulación en la sangre;
Tercero diuresis osmótica por la alta cantidad de glucosa en sangre y producción de sed para
disminuir la concentración de sales; Cuarto gluconeogénesis y acumulación en la sangre que
produce hipeglicemia; Quinto falta de energía y movilización de lípidos, para producir β
oxidación; Sexto acumulación de Acetil Co A y formación de cuerpos cetónicos.
Ahora bien, no todos los ácidos grasos que son movilizados del tejido adiposo hacia el
hígado, producen β-oxidación o cetogénesis, pues la cantidad de lípidos que se movilizan es
alta y el mismo organismo no los puede degradar a todos juntos, por ellos en la persona
diabética, existe una tercera vía asociada a los lípidos y es formación de nuevas moléculas
en triacilgliceroles (TAG).
Esta vía metabólica se produce en el citoplasma de los hepatocitos y necesitan
transportarse por la circulación, para ello existen ciertos ayudadores que los agrupan y
favorezcan su degradación, a esta moléculas se les llaman lipoproteínas (Figura No. 9)
Figura No. 9 Estructura de una lipoprotéina Tomado de http://www.saludymedicinas.com.mx/centros-de-salud/colesterol/esquemas/lipoproteinas-colesterol- bueno-y-malo.html
Las lipoproteínas son estructuras quinarias de una proteína, pues poseen un grupo
prostético (grupo no proteico), más una parte proteica conocida como apoproteína. Las
Lipoproteìnas que permiten transportar TAG desde el hígado hacia el tejido adiposo reciben
el nombre de Lipoproteínas de muy baja densidad o sus siglas en inglés VLDL; como el
hígado las está sintetizando, estas moléculas deben regresar al tejido adiposo.
En una persona sana, cuando las VLDL atraviesan la sangre, se van degradando los TAG,
para ello, se necesita de una enzima que se llama lipoprotein lipasa, esta es activada por la
apoproteína CII que posee la lipoproteína y que a mismo tiempo es activada por la presencia
de insulina.
El problema radica en que una persona diabética, no tiene la facilidad que la insulina
pueda activar esta apoproteína, por consiguiente no puede degradar los TAG, generando
aumento de VLDL con TAG a nivel sanguíneos.
Al aspecto anterior, sumemos el consumo de alimentos que poseen lípidos, estos
ingresan por la boca hacia el intestino y para poder ser transportados por el torrente sanguíneo
necesitan de un segundo tipo de lipoproteína llamada quilomicrones.
Si sumamos la cantidad de TAG que posee la VLDL y los quilomicrones, se identifica
aumento en la concentración de TAG en sangre y es por eso que las personas diabéticas
pueden llegar a padecer hipertrigliceridemia (aumento de la concentración de TAG en
sangre) (Figura No. 10).