Vista previa parcial del texto
¡Descarga bioenergetica y más Apuntes en PDF de Bioquímica solo en Docsity!
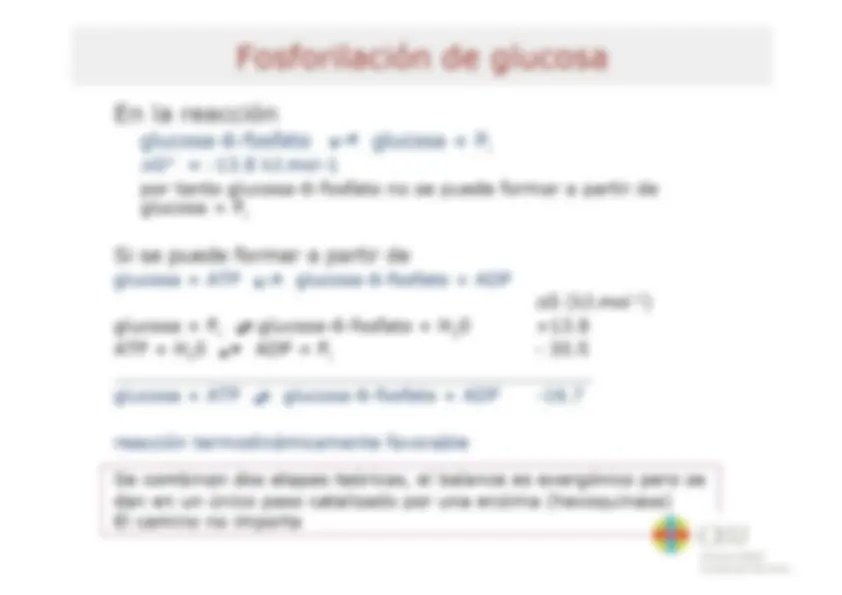
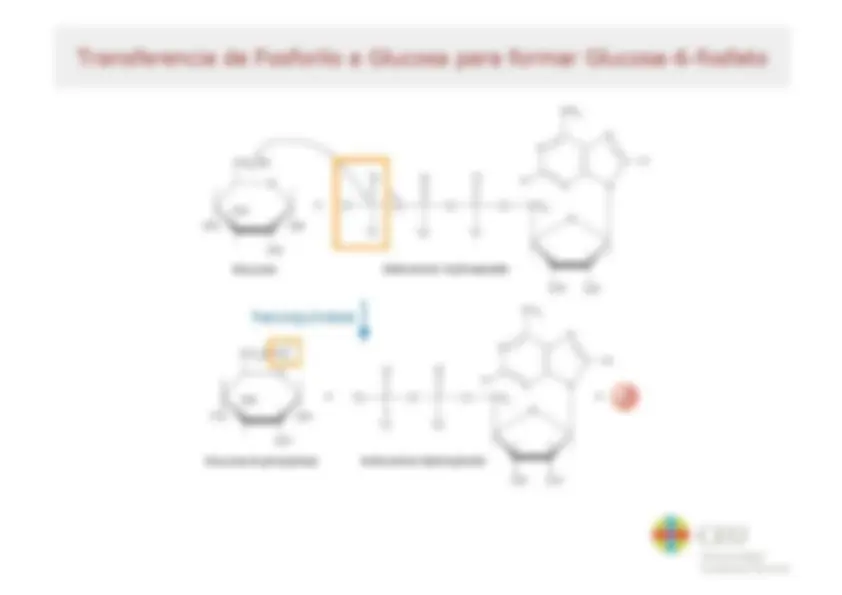
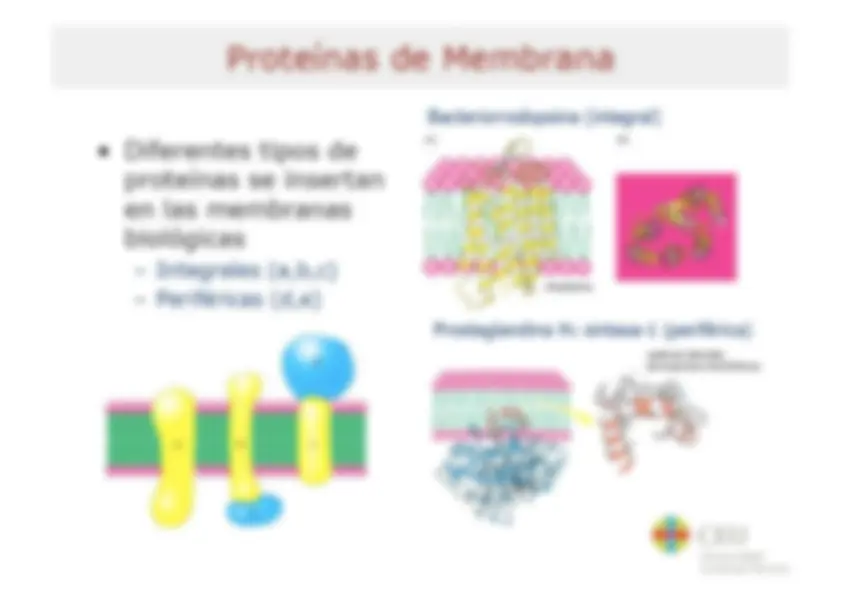
Bioenergética Tema 8 Resumen del Tema El flujo de energía en la biosfera Bases termodinámicas de las reacciones bioquímicas - Energía libre - Relación entre AG y Kg Reacciones acopladas Bioquímica del ATP - Energía libre de hidrólisis del ATP - Factores que contribuyen a la energía libre de hidrólisis del ATP - Potencial de transferencia del grupo fosforilo Sistema ATP-ADP Síntesis de ATP en las células Membranas transductoras de energía Características y propiedades de membranas biológicas Proteínas de membrana Las membranas biológicas como barreras semipermeables - Termodinámica del transporte a través de membrana - Mecanismos para el transporte a través de membrana Bomba de sodio-potasio Acoplamiento quimiosmótico ATP sintasa LB cru Universidad Cardenal Herrera Bases Termodinámicas de las Reacciones Bioquímicas Los procesos físicos y químicos que ocurren en los seres vivos cumplen las leyes de la termodinámica + Primer principio: principio de + Segundo principio la conservación de la energía - Todos los procesos - Enun proceso cualquiera la espontáneos tienden a energía total del universo realizarse de manera tal que (sistema + entorno) la entropía del universo permanece constante (sistema + entorno) siempre aumenta (S, medida del grado de desorden o de azar de un sistema) - La energía ni se crea ni se destruye, solo se transforma - El cambio de energía depende solo del estado inicial y final no del modo como ocurre la ASuniv = ASsist + ASent >0 transformación para un proceso espontáneo Los organismos vivos son sistemas abiertos, intercambian materia y energía con el entorno La entropía de un organismo disminuye a medida que crece a base de aumentar la del entorno Bos Universidad Cardenal Herrera Cambio de Energía Libre y Espontaneidad de una Reacción En sistemas abiertos como criterio de espontaneidad se utiliza la disminución en la energía libre - Energía libre como medida de la capacidad de una reacción para producir trabajo cuando el sistema se va desplazando al equilibrio a temperatura y presión constantes AG = Gprod - Greac < O - Gibbs combina el 10 y 20 principio de la termodinámica: AG = AH - TAS AG: variación de energía libre AH: variación de entalpía AS: variación de entropía T(0k) Tanto AH como AS contribuyen aAGP (simil hidraúlico) Llave abierta S se transforma en P El nivel desciende y la rueda gira (se libera energía y se efectúa trabajo) Llave abierta No hay transformación neta de Sen P No se puede efectuar trabajo AG > O no espontáneo Llave abierta Se requiere energía para convertir S en P db ceu Universidad Cardenal Herrera Relación entre AG” y K'eq AG“ y K'eq dan la misma información: una vez alcanzado el equilibrio la sustancia que mas abunda Koo AG”? (kJ/mol) A | 10% 34.2 1079 28.5 [P]<>[S] 102 11.4 y 103 17.1 dB ceu Universidad Cardenal Herrera Reacciones acopladas Muchas reacciones bioquímicas que requieren energía (endergónicas) se realizan gracias al acoplamiento a reacciones que liberan energía (exergónicas) (a) Mechanical example - La base del acoplamiento entre reacciones exergónicas y endergónicas es la existencia de un intermediario común (MA +B=>2C+D AG, > 0 (2)D + ES F+6G AG,<0 ¡3 Endergonic Exergonic MN (b) Chemical example Si |AG,|>|AG, lel proceso global será Reaction 2: termodinámicamente posible o y ANA Pl - El intermediario común puede 3 |gucose éphorphate ser un compuesto quimico, un 2 gradiente iónico o una E AG, s6 modificación de una enzima | 4%--- A6y=461+ 262 Reaction coordinate Ha Universidad Cardenal Herrera Transferencia de Fosforilo a Glucosa para formar Glucosa-6-fosfato OH HO OH OH Glucose hexoquinasa | N oroor] Qs O jo] o Ye OH + E + o HO OH 0 mo) OH Glucose-6-phosphate Adenosine diphosphate OH OH db ceu Universidad Cardenal Herrera Bioquímica del ATP El concepto de reacción acoplada sirve también para entender el papel del ATP en el metabolismo y la función termodinámica de esta molécula (“moneda energética”) Phosphoester NH2 - El ATP contiene tres grupos fosfato k 1 . - y N (a, $ y y) unidos a la adenosina Phosphoanhydride . , bonds Ñ - El fosfato a. esta unido al C5' de la o o N ribosa mediante un enlace fosfoéster zo—p!0*p."0*p, 0—CH, 0 - Los enlaces entre los fosfatos a-f y $ O O O SH H/ -y son enlaces fosfoanhídrido y H . ny frecuentemente se denominan (no Adeñosine muy correctamente) como enlaces e rico-energéticos” (=) Abe - In vivo el ATP* forma complejos con ro ¡ones Mg?* (ATP+Mg?-) B cru Universidad Cardenal Herrera Factores que contribuyen a la Energía Libre de Hidrólisis del ATP - Reducción de la repulsión electrostática entre las cargas negativas de los oxígenos en los productos de la reacción 0 0% O*- A l | Ri-0—P— 0—P— ATP+ _, ADP3-+P2 +H* ATP+ —> AMP2 + PP? + H* OS - Mayor grado de ¡onización y mejor solvatación de los productos de reacción - Mayor estabilización por resonancia en los productos respecto a la molécula intacta 0 0 La variación de energía libre (AG) de la o-p=o PA o-to reacción de hidrólisis del ATP in vivo es d- ¿- aún más negativa ya que las concentraciones de sustratos y | | productos no son 1M e influye también la 0 0 concentración de magnesio O-P-0" <> 0=P-0 | ¿- AG varia entre -50 y -60 kJ.mol* LB cru Universidad Cardenal Herrera Potencial de Transferencia del Grupo Fosforilo - Otros compuestos fosforilados TABLE 10.3 Standard free energies poseen gran tendencia a ceder el of hydrolysis for common grupo fosforilo, incluso mayor que la metabolites del ATP AG varolysis . 5 1 - Para comparar la tendencia a ceder Metabolite (kJ mol *) el grupo fosforilo de estos , Phosphoenolpyruvate 62 compuestos se utiliza el AG” de la Mia 0 . . ,3-BISphospho cerale - transferencia al H,0 (potencial de A transferencia de grupo fosforilo) Phosphocreatine ss Pyrophosphate -33 -Los compuestos con un potencial de Prosphoargiaios 32 transferencia del grupo fosforilo mayor que el ATP son excelentes dadores de ATP to AMP + PP; 32 grupo fosforilo (compuestos de elevada Acetyl COA -32 energía gía) ATP to ADP + P, -30 -Los compuestos con un potencial de Glucose 1-phosphate -21 transferencia del grupo fosforilo menor Glucose 6-phosphate 14 que el ATP son malos dadores de grupo fosforilo (compuestos de baja energía) Glycerol 3-phosphate -9 LB cru Universidad Cardenal Herrera Síntesis de ATP en las Células La síntesis de ATP requiere energía - Fosforilación a nivel de sustrato e compuestos fosforilados (alto potencial de - Electrofosforilación la disipación de un ADP +Pi —>»ATP AG = 50-60 kJ.mol 1 gluposa transferencia de grupo) ceden el fosforilo al ADP oruvato Presencia Ausencia de oxigeno e Síntesis de ATP acoplada a de ox sono / XxX lactato gradiente iónico generado Acetil-CoA a través de una membrana durante el transporte de electrones - fosforilación oxidativa (energía proviene de proceso de oxidación) - fotofosforilación (energía proviene de la luz solar) ADP + P; ha ADP + P; dB ceu Universidad Cardenal Herrera Membranas Transductoras de Energía Los mecanismos bioenergéticos más básicos no se pueden entender si no se considera la compartimentación celular, el protagonismo activo de las membranas y la esencialidad de los gradientes osmóticos y eléctricos a il tano Las membranas biológicas lipídica están constituidas por una bicapa de fosfolípidos formando un'mosaico fluido” - Modelo "mosaico fluido” de Singer y Nicolson (1972): un 'mosaico” de proteínas flotando en un mar 'fluido” de lípidos (proteínas integrales y periféricas difunden libremente) Cola hidrofóbica LB cru Universidad Cardenal Herrera Acidos Grasos Componentes de Lípidos TABLE 12.1 Some naturally occurring fatty acids in animals Number of Number of carbons double bonds Common name Systematic name Formula 12 0 Laurate n-Dodecanoate CHx(CH»)¡¿COO= 14 0 Myristate n-Tetradecanoate CHx(CH))¡¿COO7 16 0 Palmitate n- Hexadecanoate CHx(CH))4COO7 18 0 Stearate n-Octadecanoate CHx(CH))¡¿COO7 20 0 Arachidate n-Eicosanoate CHx(CH))¡g4C0OO— 22 0 Behenate n-Docosanoate CHx(CH))4COO- 24 0 Lignocerate n-Tetracosanoate CHx(CH)),C0O7 16 1 Palmitoleate cis-A”-Hexadecenoate CHy(CH))¿CH=CH(CH))¿COO= 18 1 Oleate cis-A?-Octadecenoate CHx(CH))¡CH=CH(CH))¿COO— 18 2 Linoleate cis,cis- A? AN. CH(CH 4 (CH=CHCH) (CH) ¿COOT Octadecadienoate 18 3 Linolenate all-cis-A9,412,415- — CH¿CH(CH=CHCH))¿(CH,)¿COO— Octadecatrienoate 20 4 Arachidonate all-cis-A5,A5 A31 A... CHCH))(CH=CHCH))4(CH)),COO Eicosatetraenoate db ceu Universidad Cardenal Herrera Características y Propiedades de Membranas Biológicas + Son asimétricas Fase gel o Fase fluida . E A Transición es + Son semipermeables LAT desfase e e Pueden variar su fluidez en g É => función de la temperatura A E e y y composición en ácidos 5 cada lípido una AS grasos y colesterol Tr Característica e Su fluidez permite el Oo 2000 movimiento lateral de l ' Rápido 1 moléculas l ——> | e Alojan proteínas 00 0008 transportadoras Difusión lateral e Crean compartimentos con 000 co una concentración de [| [| Muy lento | moléculas y carga de ¡ones ON I | distinta del gradiente 2.00 40) Y normal Difusión transversal *flip-flop' “o CEU Universidad Cardenal Herrera