



VISIÓN GENERAL DE LA GENERACIÓN OXIDATIVA DE ENERGÍA



Prepara tus exámenes y mejora tus resultados gracias a la gran cantidad de recursos disponibles en Docsity

Gana puntos ayudando a otros estudiantes o consíguelos activando un Plan Premium

Prepara tus exámenes
Prepara tus exámenes y mejora tus resultados gracias a la gran cantidad de recursos disponibles en Docsity
Prepara tus exámenes con los documentos que comparten otros estudiantes como tú en Docsity
Encuentra los documentos específicos para los exámenes de tu universidad
Estudia con lecciones y exámenes resueltos basados en los programas académicos de las mejores universidades
Responde a preguntas de exámenes reales y pon a prueba tu preparación
Consigue puntos base para descargar
Gana puntos ayudando a otros estudiantes o consíguelos activando un Plan Premium
Comunidad
Pide ayuda a la comunidad y resuelve tus dudas de estudio
Ebooks gratuitos
Descarga nuestras guías gratuitas sobre técnicas de estudio, métodos para controlar la ansiedad y consejos para la tesis preparadas por los tutores de Docsity
Asignatura: Bioquimica II, Profesor: jaime gomez, Carrera: Biología, Universidad: USC
Tipo: Apuntes
1 / 14

Esta página no es visible en la vista previa
¡No te pierdas las partes importantes!
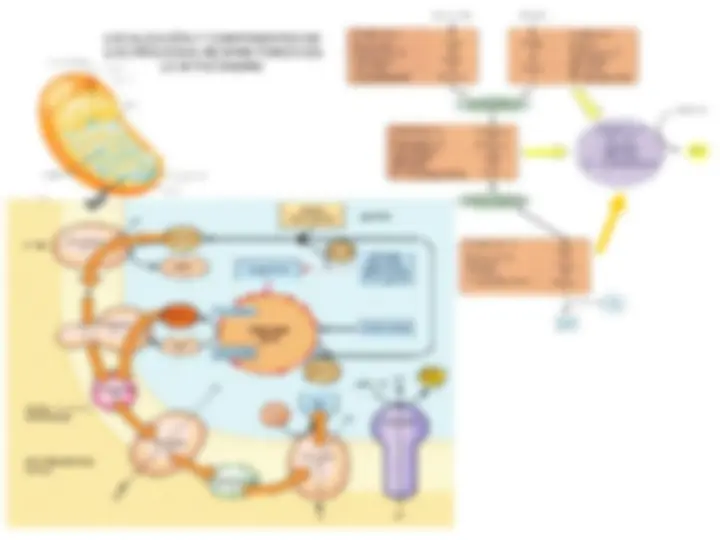
Glicerol-3-fosfato deshidrogenasa citosólica Malato deshidrogenasa citosólica Aminotransferasa Aminotransferasa (^) Malato deshidrogenasa mitocondrial Transportador glutamato/aspartato Transportador malato/ - cetoglutarato a) Lanzadera dihidroxiacetona fosfato/glicerol-3-fosfato b) Lanzadera malato-aspartato Glu= glutamato; Asp= aspartato; CG= -cetoglutarato
a) La lanzadera de la dihidoxiacetona fosfato/ glicerol-3-fosfato es el sistema más primitivo y especialmente activo en el cerebro y en el músculo de vuelo de los insectos. La dihidroxiacetona fosfato (DHAP) se reduce por el NADH en el citosol, tras lo que se produce el paso del glicerol-3-fosfato a la mitocondria, en donde se reoxida por una flavoproteína, la glicerol-3-fosfato deshidrogenasa, unida a la cara externa de la membrana interna. Este proceso comporta la reducción del FAD, seguida de la transferencia de un par de electrones desde el FADH 2 a la coenzima Q (CoQ), de la misma manera que el NADH intramitocondrial transfiere los electrones a la CoQ. Una vez que la DHAP ha regresado al citosol, el efecto neto ha sido la transferencia de dos equivalentes reductores desde el NADH citosólico al FADH 2 mitocondrial y de allí a la cadena respiratoria. b) La lanzadera del malato/aspartato es especialmente activa en el hígado y el corazón. En este caso, una isoenzima citosólica de la malato deshidrogenasa, junto con el NADH, reduce el oxalacetato a malato, que pasa a la matriz a través de un sistema de transporte activo específico de la membrana mitocondrial interna. El malato se reoxida a continuación por la malato deshidrogenasa del ciclo del ácido cítrico, que utiliza también NAD+. Dado que el oxalacetato no puede atravesar la membrana mitocondrial interna, se transamina a aspartato, que se transporta posteriormente al exterior para la reconversión en oxalacetato, con objeto de iniciar un nuevo ciclo. Dada la tansaminación que se produce en este proceso, es necesario que el - cetoglutarato se transporte continuamente al exterior de las mitocondrias y que el glutamato se transporte de manera continua al interior de las mismas.
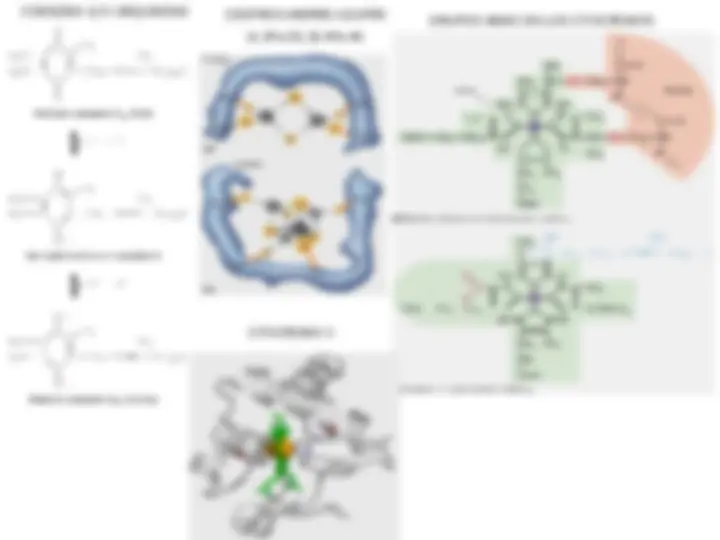
La transferencia de electrones comienza con la reducción del FMN por el NADH, un proceso que requiere 1 protón de la matriz. A continuación el FMNH 2 transfiere un par de electrones a seis u ocho centros Fe-S. Debido a que no se conoce la ruta de los electrones, sólo se muestran cuatro centros Fe-S. La transferencia secuencial de los 2 electrones al primer centro Fe-S libera en última instancia 4 protones al espacio intermembrana. Sin embargo, se cree que en la transferencia de dos de los protones participa una UQ interna. El segundo par de protones se transfiere al pasar secuencialmente los 2 electrones desde la UQ interna a través de una serie de centros Fe-S a una UQ externa. Los electrones desde el succinato se transfieren al FAD en el complejo II, a varios centros Fe-S, y luego a la UQ. Los electrones del NADH citoplasmático se transfieren a la UQ a través de una ruta con participación del glicerol-3- fosfato y la flavoproteína glicerol-3-fosfato deshidrogenasa. Los ácidos grasos se oxidan como derivados de la coenzima A. La acil-CoA deshidrogenasa, una de las enzimas de la oxidación de los ácidos grasos, transfiere 2 electrones al FAD. Luego se ceden a la UQ.
En tres pasos se producen descensos relativamente grandes de la energía libre (lugares I, II y III). En cada uno de ellos se libera energía suficiente para producir la síntesis de ATP. Este esquema de la cadena respiratoria desde el NADH al O 2 indica los lugares de acción de algunos inhibidores útiles (rojo) y algunos aceptores electrónicos artificiales (azul). Cada aceptor está situado según su valor de E ’ 0 (entre paréntesis), identificando el lugar más probable en el que un aceptor retirará los electrones de la cadena respiratoria al añadirlo a las mitocondrias. Inhibidores respiratorios
Este dibujo de los complejos proteicos de la membrana interna muestra la secuencia de transportadores electrónicos desde el NADH hasta el O 2 , así como el bombeo de protones. Los protones bombeados por los complejos I, III y IV no son, probablemente, los eliminados desde los transportadores electrónicos durante la oxidación. La reentrada de los protones en la matriz, a través del canal FO (llamado a veces complejo V), proporciona la energía necesaria para la impulsar la síntesis de ATP. No se han establecido aún las relaciones estequiométricas exactas entre las reacciones de oxidación y el bombeo de protones. En el modelo de Mitchell los protones se impulsan desde la matriz mitocondrial a través de la membrana mitocondrial y dentro del espacio intermembrana por el mecanismo de transporte electrónico. La energía capturada del transporte electrónico se utiliza para crear un potencial eléctrico y un gradiente de protones. Debido a que la membrana interna es impermeable a los protones, sólo pueden atravesar la membrana fluyendo a través de canales específicos de protones. El flujo de protones a través de la ATP sintasa impulsa la síntesis de ATP.
El transporte de H 2 PO 4 -^ a través de la membrana mitocondrial interna por la translocasa de fosfato está impulsado por el gradiente de protones. Por cada 4 protones que se transportan fuera de la matriz, 3 impulsan el rotor de la ATP sintasa y 1 impulsa el transporte de fosfato al interior. El intercambio simultáneo de ADP3-^ y ATP4-, que se requiere para la síntesis continua de ATP y que se produce por el translocalizador ADP- ATP, está impulsado por la diferencia de potencial a través de la membrana interna.
Transportadores de electrones de la membrana interna de las mitocondrias de plantas. Los electrones pueden fluir a través de los complejos I, III y IV, al igual que en las mitocondrias de animales, o a través de transportadores alternativos específicos de plantas mediante las vías mostradas en azul. La proteína desacopladora, termogenina, de las mitocondrias de la grasa marrón, al proporcionar una ruta alternativa para que los protones vuelvan a entrar a la matriz mitocondrial, hace que la energía conservada por el bombeo de protones se disipe en forma de calor.
El L- ascorbato, una molécula hidrosoluble, protege a las membranas del daño oxidativo regenerando el - tocoferol a partir del radical - tocoferilo. El radical ascorbilo que se forma en este proceso se reconvierte en L-ascorbato durante una reacción con GSH.
La glutatión peroxidasa utiliza el GSH para reducir los peróxidos generados por el metabolismo aerobio celular. La glutatión reductasa regenera GSH a partir de su forma oxidada, GSSG. El NADPH, el reductor de esta reacción, lo aportan la ruta de las pentosas fosfato y otras reacciones. El estallido respiratorio proporciona un ejemplo espectacular del efecto destructivo de las ROS. A los pocos segundos de unirse una célula fagocítica a una bacteria (u otra estructura ajena), su consumo de oxígeno aumenta cerca de 100 veces. Durante la endocitosis, la bacteria se incorpora a una gran vesícula que se denomina fagosoma. Los fagosomas se fusionan con los lisosomas para formar fagolisosomas. Tienen lugar entonces dos procesos destructores: el estallido respiratorio y la digestión por enzimas lisosómicas. El estallido respiratorio se inicia cuando la NADPH oxidasa convierte el O 2 en O 2 -.. Dos moléculas de O 2 -.^ se combinan en una reacción que cataliza la superóxido dismutasa (SOD) para formar H 2 O 2. Éste se convierte a continuación en varias clases de moléculas bactericidas por la mieloperoxidasa (MPO), una enzima que se encuentra en abundancia en los fagocitos. Por ejemplo, la MPO cataliza la oxigenación de los iones haluro (p. ej., Cl-) para formar hipohaluros. El hipoclorito (el ingrediente activo de la lejía casera) es extremadamente bactericida. En presencia de Fe2+, el O 2 -.^ y el H 2 O 2 reaccionan para formar · OH y 1 O 2 (oxígeno singlete), ambos muy reactivos. Tras desintegrarse la célula bacteriana, las enzimas lisosómicas digieren los fragmentos que quedan.