¡Descarga BIOQUÍMICA II y más Apuntes en PDF de Bioquímica solo en Docsity!
TEMA XV
DEGRADACIÓN DE ÁCIDOS GRASOS: ß-OXIDACIÓN,
METABOLISMO DE CUERPOS CETÓNICOS.
UBICACIÓN CELULAR DEL METABOLISMO DE LÍPIDOS
Citosol: producción del NADPH, hace alusión a la ruta de las pentosas fosfato que fabrica NADPH imprescindible para la síntesis de lípidos. También ocurre la síntesis de isoprenoides y esteroles (de los estadios tempranos de su síntesis), también la síntesis de a.g. RE: síntesis de fosfolípidos, síntesis de los estados tardíos de los esteroles, elongación de a.g o desaturación de a.g (en plantas ocurre lo mismo). Mitocondria: oxidación de los a.g., producción de acetil-CoA, la síntesis de cuerpos cetónicos y elongación de a.g. (en plantas no ocurre la oxidación de a.g., en estas ocurre en los peroxisomas). Cloroplasto: producción de NADPH y síntesis de aa.
CATABOLISMO DE ÁCIDOS GRASOS
ORÍGENES DE LOS AG
Exógeno: triacilglicérdios que están en los alimentos y son ingeridos. Endógeno: triacilglicéridos que tenemos en el tejido adiposo y la síntesis de “novo” a partir de acetil-CoA. Los TAG están en el tejidos adiposos y cuando son degradados se generan: Glicerol: (precursor gluconeogénico) que pasa al hígado donde puede entrar en la ruta de la glucólisis generando piruvato o en la ruta de gluconeogénesis generando glucosa. Ácidos grasos que por medio de la oxidación forman Acetil-CoA que entra en el ciclo del ácido cítrico que finalmente genera CO2 + H2O. MOVILIZACIÓN DE LOS TAG DEL TEJIDO ADIPOSO Cuando se movilizan las reservas de grasa se hace para obtener energía, entonces los ácidos grasos se degradarán para obtener ATP. Cuando los a.g. salen a la sangre van unidos por la seroalbumina (proteína) y son transportados a través de la sangre hasta los tejidos. Lo que hace que nuestro organismo movilice las grasas: LAS HORMONAS (glucagón, adrenalina) las dos se producen bajo situaciones diferentes. Su mecanismo de acción es el mismo, mediadas por proteínas G que generan AMPc, que a su vez activa a la PROTEÍN KINASA A, que fosforila la LIPASA sensible a hormonas ( TRIACILGLICEROL LIPASA ), se activa esta y necesita que la PKA fosforile a las perilipinas que están recubriendo a las gotículas de lípidos para evitar que se degraden y cuando las fosforilan se facilita el acceso de la LIPASA a las gotículas y la hidrólisis de los ácidos grasos (se necesita la actuación de las tres GLICEROL LIPASAS para dar lugar a a.g libres: TRIACILGLICEROL LIPASA , DIACILGLICEROL LIPASA y MONOACILGLICEROL LIPASA.
TEMA XV: DEGRADACIÓN DE ÁCIDOS GRASOS: β-
OXIDACIÓN, METABOLISMO DE CUERPOS CETÓNICOS
UBICACIÓN INTRACELULAR DEL METABOLISMO DE LÍPIDOS La oxidación y elongación de ácidos grasos, producción acetil-coA y síntesis de cuerpos cetónicos ocurren en la célula animal, en la mitocondria. Pero no la oxidación de ácidos grasos. En el retículo endoplasmático ocurre la síntesis de fosfolípidos, sisntesis de esteroides, elongación de ácidos grasos y desnaturación de ácidos grasos. En el citosol ocurre la producción de NADPH, NADPH/NADP+, isoprenoides, sintesis de colesterol y ácidos grasos. En la célula vegetal; en los peroxisomas ocurre la oxidación de los ácidos grasos y en los cloroplastos la producción de NADPH, ATP y síntesis de ácidos grasos. ORIGEN DE LOS ÁCIDOS GRASOS Los ácidos grasos tienen un origen exógeno generalmente son los triglicéridos de las grasas. En el tema anterior ya se vió como se transportan con quilomicrones en la vía exógena. Los triacilglicéridos están en el tejido adiposo y cuando se degradan generan:
- Glicerol (precursor gluconeogénico) que pasa al hígado donde puede entrar en la ruta de la glucolisis generando piruvato o en la ruta de gluconeogénesis generando glucosa.
- Ácidos grasos que por medio de la oxidación forman acetil-CoA que entra en el ciclo del ácido cítrico que finalmente genera CO 2 + H 2 O MOVILIZACIÓN DE TAG DEL TEJIDO ADIPOSO Para la movilización de las TAG se necesita de una orden que viene dada por la producción de hormonas que actúan con repceptores mediados por la proteína G que genera AMPc, que a su vez activa la protein quinasa a que fosforila la triglicerol lipasa que se convierte en diacilglicerol. Con niveles bajos de glucosa en sangre activan la liberación de glucagón.
- La hormona se une a su receptor en la membrana del adipocito
- Asi se estimula la adenil ciclasa, vía una proteína G, para producir AMPc. Esto activa la Triacilglicéridos glicerol piruvato glucosa Ácidos grasos Acetil-CoA CAC CO 2 + H 2 O glucolisis gluconeogénesis Oxidación ac. grasos
MECANISMO DE MOVILIZACIÓN DE LOS A.G
Con niveles bajos de glucosa en sangre se activa la liberación de glucagón: 1º La hormona se une a su receptor en la membrana del adipocito. 2º Así se estimula la ADENIL CICLASA , vía una proteína G, para producir AMPc. Esto activa a la PKA. 3º La PKA fosforila la LIPASA sensible a hormona. 4º La PKA también fosforila a moléculas de perilipina de la superficie de la gotícula del lípido. La fosforilación de la perilipina permite el acceso de la LIPASA sensible a hormona a la superficie de la gotícula de lúpulo. 5º Aquí es donde se hidrolizan los TAG a ácidos grasos libres. 6º Los ácidos grasos abandonan el adipocito, se unen a la albumina sérica en la sangre y se transportan por el torrente sanguíneo. 7º Se liberan de la albumina y entran en un miocito mediante un transportador específico de ácidos grasos. 8º En el miocito, los ácidos grasos se oxidan a CO2, al tiempo que la energía de oxidación se conserva en forma de ATP, el cual promueve la contracción muscular y otros procesos metabólicos que requieren energía en el miocito. OXIDACIÓN DE ÁCIDOS GRASOS La ß-oxidación consiste en generar cadenas de 2 carbonos de manera que: 16 C -> 8 acetil-CoA. Leningher demostró que ocurría en la mitocondria y se empiezan con la oxidación del Cß. En cada vuelta del ciclo se oxida y se libera acetil-CoA, así repetidamente. La ß-oxidación es una ruta que puede dividirse en 3 etapas: 1~ Activación de los ácidos grasos: los ácidos grasos son activados en el citosol mediante su conversión a tioésteres de coenzima A ( ACIL-COA ) catalizada por ACIL-COA SINTETASA , en una reacción que consume ATP. Este enzima se encuentra en la membrana mitocondrial externa. sensible a hormona.
- La PKA también fosforila a
moléculas de perilipina de la superfície de la gotícula de lípido. La fosforilación de la perilipina permite el acceso de la lipasa sensible a hormona a la superficie de la gotícula de lúpudo.
- Aquí es donde se hidrolizan los
triacilgliceroles a ácidos grasos libres
- Los ácidos grasos abandonan el
adipocito, se unen a la albumina sérica en la sangre y se transportan por el torrente circulatorio
- Se liberan de la albumina y entran
en eun miocito mediante un transportador específico de ácidos grasos.
- En el miocito, los ácidos grasos se
oxidan a CO2, al tiempo que la energía de oxidación se conserva en forma de ATP, el cual promueve la contracción muscular y otros procesos metabólicos que requieren energía en el miocito. OXIDACIÓN DE ÁCIDOS GRASOS La β-oxidación consiste en generar cadenas de 2 carbonos de manera que: 16 C → 8 acetil-CoA. Leningher demostró que ocurría en la mitocondria y se empiezan con la oxidación del Cβ. En cada vuelta del ciclo se oxida y se libera Acetil-CoA, así repetidamente. De manera que la oxidación de los ácidos grasos tienen 3 fases:
- Activación de los ácidos grasos : formación
de acetil-CoA grasos.
- Transporte de ácidos grasos a la
mitocondria : en los ácidos grasos de cadena larga se necesita de una molécula llamada carnitina y la cadena pasa al interior de la mitocondria y ahi es dónde ocurre la β- oxidación.
- Reacciones de la β-oxidación.
ACTIVACIÓN La activación está catalizada por Acetil-CoA sintasa (ácido graso toquinasa): ácido graso + HS-CoA Acetil-CoA Este proceso ocurre en dos pasos, ambos están próximos al equilibrio pero en la práctica esta reacción está dirigida a la síntesis de Acetil-CoA porque se hidroliza PPi por la pirofosfatasa inorgánica formando 2Pi. Para la formación del Acetil-CoA consumimos 2 enlaces fosfato ricos en energía de modo que ha que tener en cuenta el balance: R-COO- + CoA +ATP + H 2 O → RCO-SCoA + AMP + 2Pi +ATP AMP+ PPi
ACTIVACIÓN
La activación está catalizada por Acetil-CoA sintasa (ácido graso toquinasa): ácido graso + HS-CoA Acetil-CoA Este proceso ocurre en dos pasos, ambos están próximos al equilibrio pero en la práctica esta reacción está dirigida a la síntesis de Acetil-CoA porque se hidroliza PPi por la pirofosfatasa inorgánica formando 2Pi. Para la formación del Acetil-CoA consumimos 2 enlaces fosfato ricos en energía de modo que ha que tener en cuenta el balance: R-COO- + CoA +ATP + H 2 O → RCO-SCoA + AMP + 2Pi +ATP AMP+ PPi te el acceso de la lipasa le a hormona a la superficie otícula de lúpudo. s donde se hidrolizan los gliceroles a ácidos grasos idos grasos abandonan el cito, se unen a la albumina en la sangre y se ortan por el torrente atorio eran de la albumina y entran miocito mediante un ortador específico de ácidos . miocito, los ácidos grasos se n a CO2, al tiempo que la a de oxidación se conserva ma de ATP, el cual eve la contracción lar y otros procesos ólicos que requieren energía iocito. OXIDACIÓN DE ÁCIDOS GRASOS n consiste en generar cadenas de 2 carbonos de manera que: 16 C → 8 acetil-CoA. ostró que ocurría en la mitocondria y on la oxidación del Cβ. En cada vuelta ida y se libera Acetil-CoA, así . e la oxidación de los ácidos grasos ación de los ácidos grasos : formación etil-CoA grasos. porte de ácidos grasos a la ondria : en los ácidos grasos de cadena se necesita de una molécula llamada tina y la cadena pasa al interior de la ondria y ahi es dónde ocurre la β- ción. iones de la β-oxidación.
3~ ß-oxidación con 4 tipos de reacciones: Oxidación: la enzima que cataliza esta reacción es la ACIL-COA DESHIDROGENASA que utiliza como coenzima el FAD. Hidratación: la enzima que cataliza esta reacción es la ENOIL-COA HIDRATASA , se introduce agua que rompe el enlace. Oxidación: la enzima implicada es la ß-HIDROXIACIL-COA DESHIDROGENASA , el coenzima que utiliza es el NAD* que retira 2 hidrógenos. Tiolisis: es una rotura en la que está implicada la ACIL-COA ACETILTRANSFERASA (tiolasa) que coge el S de CoA-SH y Acetil-CoA inicial queda reducido en 2C. Ese compuesto vuelve a sufrir estas reacciones. Con el FADH2 se oxida cediendo los electrones a la proteína ETF ( electrón transferín flavoproteín ) que se reduce. A su vez se reduce otra proteína ETF ubiquinona oxidorreductasa que reduce al CoQ que cede electrones al complejo III y continúa su camino hasta formar agua y produciendo 2 ATP y NADH cede electrones a la cadena respiratoria a nivel del complejo I y produce 3 ATP junto con Acetil-CoA se incorpora el ciclo de Krebs. BALANCE DE LA OXIDACIÓN DE PALMITATO (16C) A CO2 Y H2O En total se generan 108 ATP en la oxidación del ácido palmítico pero gastamos 2 en la activación por tanto el ATP final es 106. Se requieren 7 ciclos antes de llegar a él a través de la secuencia ß-oxidación para oxidar una molécula de palmitoil-CoA a 8 moléculas de acetil-CoA. El acetil-CoA producido en la oxidación de los ácidos grasos puede oxidarse a CO2 y H2O a través del ciclo del ácido cítrico. LA DEGRADACIÓN MITOCONDRIAL DE LOS ÁCIDOS GASOS : ß-OXIDACIÓN Antiguamente, la beta oxidación se le denominó hélice de Lynen. Se llama beta oxidación porque el proceso de degradación empieza con la degradación de un carbono beta. AG SATURADOS => de cadena par: 8 acetil-CoA. eliminación sucesiva de 2 carbones de acetil-CoA. 4 reacciones básicas: oxidación, hidratación, oxidación, tiolisis. Esto se repite n veces hasta convertir todo el a.g. en acetil-CoA. de cadena impar: acetil-CoA y propionil-CoA. El los ácidos grasos de cadena impar en la penúltima vuelta se forma un ácido graso de 5 C que genera un acetil-CoA de 2 C y Propionil-CoA de 3C. El propionil no se puede integra en el ciclo de Krebs, por tanto: La β-oxidación tiene 4 tipos de reacciones:
- oxidación: la enzima que cataliza esta reaccion es la acetil-CoA deshidrogenasa que utiliza como coenzima el FAD.
- Hidratación, la enzima que cataliza esta reacción es la enoil-CoA hidratasa, se introduce agua que rompe el doble enlace.
- Oxidación: la enzima implicada es la β-hidroxiacil- CoA deshidrogenasa, el coenzima que utiliza es el NAD+ que retira 2 hidrógenos.
- Tiolisis: es una rotura en la que está implicada la acil- CoA acetiltransferasa (tiolasa) que coge el S de CoA-SH y Acetil-CoA inicial que reducido en 2 C. Ese compuesto vuelve a sufrir esas reacciones Con el FADH 2 se oxida cediendo los electrones a la proteína ETF (electron- transferin flavoprotein) que se reduce. A su vez se reduce otra proteína ETF ubiquinona oxidorreductasa que reduce al CoQ que cedeelectrones al complejo III y continúa su camino hasta formar agua y produciendo 2 ATP y NADH cede electrones a la cadena respiratoria a nivel del complejo I y produce 3 ATP junto con Acetil-CoA se incorpora el ciclo de krebs. Carbonos Acetil-CoA Vueltas Mitad - 12 6 5 14 7 6 16 8 7 18 9 8 Balance de la oxidación palmitato (16C) a CO 2 + H 2 O. Palmitoil-CoA (7 vueltas) + 7 CoA + 7 FAD + 7 NAD+ + H2O → en total se generan 108 ATP en la oxidación del ácido palmítico pero gastamos 2 en la activación por tanto el ATP final es 106. 8 Acetil-CoA → 80 ATP 7 FADH 2 → 10'5 ATP 7 NADH+7+ → 15'5 ATP Acetil-CoA/ 10 ATP NADH/ 2'5 ATP FADH2/ 1'5 ATP que utiliza como coenzima el FAD.
- Hidratación, la enzima que cataliza esta reacción es la enoil-CoA hidratasa, se introduce agua que rompe el doble enlace.
- Oxidación: la enzima implicada es la β-hidroxiacil- CoA deshidrogenasa, el coenzima que utiliza es el NAD+ que retira 2 hidrógenos.
- Tiolisis: es una rotura en la que está implicada la acil- CoA acetiltransferasa (tiolasa) que coge el S de CoA-SH y Acetil-CoA inicial que reducido en 2 C. Ese compuesto vuelve a sufrir esas reacciones Con el FADH 2 se oxida cediendo los electrones a la proteína ETF (electron- transferin flavoprotein) que se reduce. A su vez se reduce otra proteína ETF ubiquinona oxidorreductasa que reduce al CoQ que cedeelectrones al complejo III y continúa su camino hasta formar agua y produciendo 2 ATP y NADH cede electrones a la cadena respiratoria a nivel del complejo I y produce 3 ATP junto con Acetil-CoA se incorpora el ciclo de krebs. Carbonos Acetil-CoA Vueltas Mitad - 12 6 5 14 7 6 16 8 7 18 9 8 Balance de la oxidación palmitato (16C) a CO 2 + H 2 O. Palmitoil-CoA (7 vueltas) + 7 CoA + 7 FAD + 7 NAD+ + H2O → en total se generan 108 ATP en la oxidación del ácido palmítico pero gastamos 2 en la activación por tanto el ATP final es 106. 8 Acetil-CoA → 80 ATP 7 FADH 2 → 10'5 ATP 7 NADH+7+ → 15'5 ATP Acetil-CoA/ 10 ATP NADH/ 2'5 ATP FADH2/ 1'5 ATP doble enlace.
- Oxidación: la enzima implicada es la β-hidroxiacil- CoA deshidrogenasa, el coenzima que utiliza es el NAD+ que retira 2 hidrógenos.
- Tiolisis: es una rotura en la que está implicada la acil- CoA acetiltransferasa (tiolasa) que coge el S de CoA-SH y Acetil-CoA inicial que reducido en 2 C. Ese compuesto vuelve a sufrir esas reacciones Con el FADH 2 se oxida cediendo los electrones a la proteína ETF (electron- transferin flavoprotein) que se reduce. A su vez se reduce otra proteína ETF ubiquinona oxidorreductasa que reduce al CoQ que cedeelectrones al complejo III y continúa su camino hasta formar agua y produciendo 2 ATP y NADH cede electrones a la cadena respiratoria a nivel del complejo I y produce 3 ATP junto con Acetil-CoA se incorpora el ciclo de krebs.
Carbonos Acetil-CoA Vueltas
Mitad -
Balance de la oxidación palmitato (16C) a CO 2 + H 2 O. Palmitoil-CoA (7 vueltas) + 7 CoA + 7 FAD + 7 NAD+ + H2O → en total se generan 108 ATP en la oxidación del ácido palmítico pero gastamos 2 en la activación por tanto el ATP final es 106. 8 Acetil-CoA → 80 ATP 7 FADH 2 → 10'5 ATP 7 NADH+7+ → 15'5 ATP Acetil-CoA/ 10 ATP NADH/ 2'5 ATP FADH2/ 1'5 ATP doble enlace.
- Oxidación: la enzima implicada es la β-hidroxiacil- CoA deshidrogenasa, el coenzima que utiliza es el NAD+ que retira 2 hidrógenos.
- Tiolisis: es una rotura en la que está implicada la acil- CoA acetiltransferasa (tiolasa) que coge el S de CoA-SH y Acetil-CoA inicial que reducido en 2 C. Ese compuesto vuelve a sufrir esas reacciones Con el FADH 2 se oxida cediendo los electrones a la proteína ETF (electron- transferin flavoprotein) que se reduce. A su vez se reduce otra proteína ETF ubiquinona oxidorreductasa que reduce al CoQ que cedeelectrones al complejo III y continúa su camino hasta formar agua y produciendo 2 ATP y NADH cede electrones a la cadena respiratoria a nivel del complejo I y produce 3 ATP junto con Acetil-CoA se incorpora el ciclo de krebs.
Carbonos Acetil-CoA Vueltas
Mitad -
Balance de la oxidación palmitato (16C) a CO 2 + H 2 O. Palmitoil-CoA (7 vueltas) + 7 CoA + 7 FAD + 7 NAD+ + H2O → en total se generan 108 ATP en la oxidación del ácido palmítico pero gastamos 2 en la activación por tanto el ATP final es 106. 8 Acetil-CoA → 80 ATP 7 FADH 2 → 10'5 ATP 7 NADH+7+ → 15'5 ATP Acetil-CoA/ 10 ATP NADH/ 2'5 ATP FADH2/ 1'5 ATP introduce agua que rompe el doble enlace.
- Oxidación: la enzima implicada es la β-hidroxiacil- CoA deshidrogenasa, el coenzima que utiliza es el NAD+ que retira 2 hidrógenos.
- Tiolisis: es una rotura en la que está implicada la acil- CoA acetiltransferasa (tiolasa) que coge el S de CoA-SH y Acetil-CoA inicial que reducido en 2 C. Ese compuesto vuelve a sufrir esas reacciones Con el FADH 2 se oxida cediendo los electrones a la proteína ETF (electron- transferin flavoprotein) que se reduce. A su vez se reduce otra proteína ETF ubiquinona oxidorreductasa que reduce al CoQ que cedeelectrones al complejo III y continúa su camino hasta formar agua y produciendo 2 ATP y NADH cede electrones a la cadena respiratoria a nivel del complejo I y produce 3 ATP junto con Acetil-CoA se incorpora el ciclo de krebs.
Carbonos Acetil-CoA Vueltas
Mitad -
Balance de la oxidación palmitato (16C) a CO 2 + H 2 O. Palmitoil-CoA (7 vueltas) + 7 CoA + 7 FAD + 7 NAD+ + H2O → en total se generan 108 ATP en la oxidación del ácido palmítico pero gastamos 2 en la activación por tanto el ATP final es 106. 8 Acetil-CoA → 80 ATP 7 FADH 2 → 10'5 ATP 7 NADH+7+ → 15'5 ATP Acetil-CoA/ 10 ATP NADH/ 2'5 ATP FADH2/ 1'5 ATP
El succinil-CoA ya se puede integrar en el ciclo de Krebs, ya que es un intermediario de este. La mayoría de vitaminas actúan como coenzimas sobre todos los hidrosolubles. AG INSATURADOS: la oxidacion de los acidos grasos que presentan dobles enlaces requiere la acción de dos enzimas: una ISOMERASA y una REDUCTASA. monoinsaturados: requiere un enzima a mayores, una ISOMERASA , que convierte el isómero cis en trans, un intermediario normal de la ß-oxidación. Ácido oleico : se usa una isomerasa. Hay 3 ciclos de ß- oxidación normal y se liberan 3 moléulas de Acetil-CoA. Al estar en cis no es sustrato de la HIDRATASA. Aquí, es dónde actúa la ENOIL-COA ISOMERASA, pasa de cis a trans, esto es sustrato de la ENOIL-COA HIDRATASA. El succinil-CoA ya se puede integrar en el ciclo de krebs, ya que es un intermediario de este. La mayoría de vitaminas actúan como coenzimas sobre todo los hidrosolubles. + Ácido graso monoinsaturado Los ácidos grasos santurados son los que siguen las fases de oxidación-hidratación-oxidación-tiolisis en la oxidación. En los ácidos grasos insaturados hay 2 tipos:
- Monoinsaturados (ácido oleico): se usa una isomerasa. Hay 3 ciclos de β-oxidación normal y se liberan 3 moléculas de Aceil-CoA. Al estar en cis no es sustrato de la hidratasa. Aqui es dónde actúa la enoil-CoA isomerasa que lo hace pasar de cis a trans, esto es sustrato de la hidratasa (enoil-CoA hidratasa) Ilustración 1 : monoinsaturados El succinil-CoA ya se puede integrar en el ciclo de krebs, ya que es un intermediario de este. La mayoría de vitaminas actúan como coenzimas sobre todo los hidrosolubles. + Ácido graso monoinsaturado Los ácidos grasos santurados son los que siguen las fases de oxidación-hidratación-oxidación-tiolisis en la oxidación. En los ácidos grasos insaturados hay 2 tipos:
- Monoinsaturados (ácido oleico): se usa una isomerasa. Hay 3 ciclos de β-oxidación normal y se liberan 3 moléculas de Aceil-CoA. Al estar en cis no es sustrato de la hidratasa. Aqui es dónde actúa la enoil-CoA isomerasa que lo hace pasar de cis a trans, esto es sustrato de la hidratasa (enoil-CoA hidratasa)
Ilustración 1 : monoinsaturados
ω-OXIDACIÓN Ocurre en el Retículo endoplasmático de células animales del hígado y riñones. Los sustratos son ácidos grasos de tamaño medio (10-12 C) Tienen importancia cuando la ß-oxidación es defectuosa. En condiciones normales es poco relevante. α-OXIDACIÓN Oxidación de los ácidos grasos ramificados, ocurre en los peroxisomas. Se llama así porque empieza en la oxidación del C alfa, mientras que en la ß-oxidación se empezaba en el C-ß. Se produce porque en el C-ß hay una ramificación metilo, de manera que no se puede empezar en ese C.
- La^ enfermedad de^ RefsuM .: trastorno congénito debido a la ausencia de la^ FITANIL-COA HIDROLASA , de manera
que no se puede reducir el ácido fitánico aumentando la concentración de este en sangre y causando entre, otras cosas ceguera. α-OXIDACIÓN queremos oxidar los ácidos grasos roxisomas. Se llama así porque α mientras que en la β-oxidación se la α-oxidación porque en el Cβ hay anera que no se puede empezar en fsum nito grave debido a fitonil-CoA ta de la α-oxidación es defectuosa o fitónico aumentando la e. Causa entre otras cosas ceguera. IDACIÓN de las células animales, en las n de vertebrados. s de tamaño medio (10-12C) -oxidación es defectuosa. En relevante.
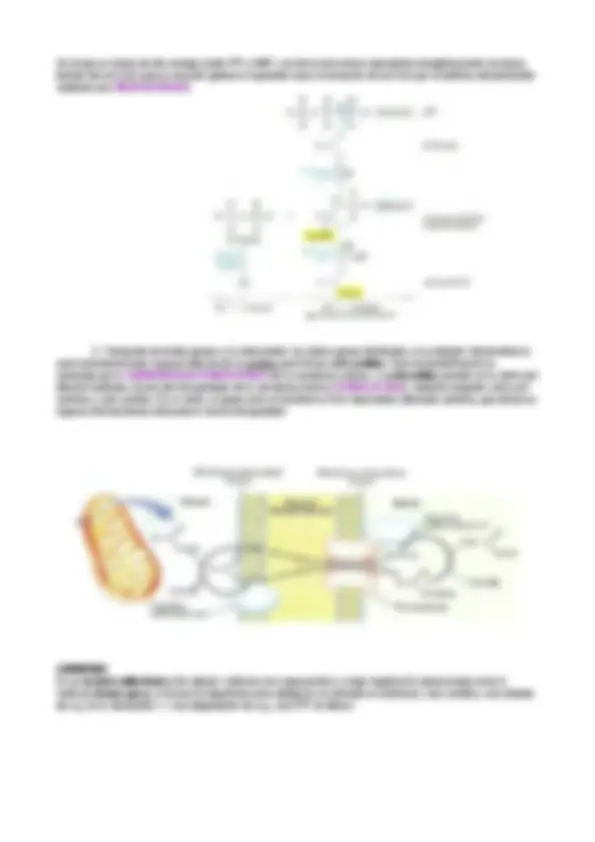
METABOLISMO DE CUERPOS CETÓNICOS
CUERPOS CETÓNICOS
Acetona , aceto-acetato , hidroxibutirato. Su síntesis se hace a partir de Acetil-CoA. Este, formado en el hígado durante la oxidación de ácidos grasos puede entrar en el ciclo del ácido cítrico o puede ser convertido en cuerpos cetónicos. Se producen en la mitocondrias hepáticas y son fuente de energía para el cerebro, corazón, músculo en baja concentración de glucosa en sangre, en estados de ayuno. La acetona se exhala, el aceto-acetato & hidroxibutirato son transportados en sangre a tejidos extrahepáticos. Los cuerpos cetónicos son como ácidos grasos porque se sintetizan a partir del Acetil-CoA que se produce a partir de los ácidos grasos. METABOLISMO
- La degradación: en el hígado no. El hígado puede fabricar cuerpos cetónicos pero no los puede degradar porque carece de la enzima ß- CETOACILTRANSFERASA, enzima que cataliza la reacción de aceto-acetato + succinil-CoA para dar lugar a succinato y acetoacetil-CoA que, a su vez, se rompe en dos acetil-CoA los cuales se integran en el ciclo de Krebs para la obtención de ATP.
- La síntesis: mitocondria. La TIOLASA realiza la condensación de 2 Acetil-CoA y forma el aceto-acetil- CoA. A esta se le añade otra molécula de Acetil-CoA y, mediante una reacción catalizada por la HMG-CoA SINTASA se forma el HMG-CoA , este a su vez, mediante la HMG-CoA LIASA , se rompe en aceto-acetato y por descarboxilación da lugar a acetona (reacción espontánea) o dar lugar al hidroxibutirato (en una reducción en la que interviene el NADH). La acetona se expulsa del cuerpo por la orina o exhalación y los otros dos sufren degradación (en el hígado no) El hidroxibutirato se convierte en aceto-acetato porque la reacción es próxima al equilibrio. CUERPOS CETÓNICOS (ketan Los cuerpos cetńicos son una fuente especial de energía para condiciones. Son 3: a cetona, aceto acetato y hidroxibutira Se produce en la mitocondria hepática cuando el hígado no p de manera que son la principal fuente de energía para el cere recibir energía que consigue en los cuerpos cetónicos. La acetona se va a expulsar, exhalar por el alimento. El aceto transportados vía sanguínea a tejidos extrahepáticos. Los cuerpos cetónicos son como ácidos grasos porque se sin se produce a partir de ácidos grasos.
- Metabolismo de cuerpos cetónicos Ilustración 3 : Ilustración 4 : Degradación (Metabolito del ciclo de krebs) (se rompe en dos moléculas de acetil CoA) (Pasan al Ciclo de Krebs) El hígado carece de tiofarasa por tanto no puede degradar los cuerpos cetónicos y los libera ala sangre que llega a los tejidos ÓNICOS (ketane bodies) l de energía para algunos tejidos en determinadas y hidroxibutirato. do el hígado no puede consumirlas/metabolizarlas ergía para el cerebro en ayuno ya que este no puede cetónicos. limento. El acetoacetato e hidroxibutirato son epáticos. sos porque se sintetizan a partir de Acetil-CoA que os Ilustración 3 : Síntesis bs) s de acetil CoA)