¡Descarga CAPITULO 16 HARPER 30 edicion y más Transcripciones en PDF de Bioquímica Médica solo en Docsity!
El ciclo del ácido cítrico:
la vía central del metabolismo
de carbohidratos, lípidos
y aminoácidos
David A. Bender, PhD & Peter A. Mayes, PhD, DSc
■ ■ (^) Describir las reacciones del ciclo del ácido cítrico y las reacciones que llevan a la producción de equivalentes reductores que son oxidados en la cadena de transporte de electrones mitocondrial para dar ATP. ■ ■ (^) Comprender la importancia de las vitaminas en el ciclo del ácido cítrico. ■ ■ (^) Explicar cómo el ciclo del ácido cítrico proporciona tanto una ruta para el catabolismo de aminoácidos como una ruta para su síntesis. ■ ■ (^) Identificar las principales vías anapleróticas que permiten el reabastecimiento de intermediarios del ciclo del ácido cítrico, y cómo está controlado el retiro de oxaloacetato para gluconeogénesis. ■ ■ (^) Definir la función del ciclo del ácido cítrico en la síntesis de ácidos grasos. ■ ■ (^) Explicar cómo la actividad del ciclo del ácido cítrico está controlada por la disponibilidad de cofactores oxidados. ■ ■ (^) Explicar cómo la hiperamonemia puede llevar a pérdida del conocimiento.
O B J E T I V O S
Después de estudiar este capítulo, usted debe ser capaz de:
C A P í T u l o
ImportancIa bIomédIca
El ciclo del ácido cítrico (ciclo de Krebs, ciclo del ácido tricar- boxílico) es una secuencia de reacciones en las mitocondrias que oxidan la porción acetilo de la acetil-CoA, y reducen coenzimas que se reoxidan por medio de la cadena de transporte de electro- nes (capítulo 13), enlazada a la formación de ATP. El ciclo del ácido cítrico es la vía común final para la oxida- ción de carbohidratos, lípidos y proteínas porque la glucosa, los ácidos grasos y casi todos los aminoácidos se metabolizan hacia acetil-CoA o intermediarios del ciclo. También tiene una función fundamental en la gluconeogénesis, lipogénesis e interconver- sión de aminoácidos. Muchos de estos procesos ocurren en casi todos los tejidos, pero el hígado es el único tejido en el cual todos suceden en un grado significativo. En consecuencia, hay pro- fundas repercusiones cuando, por ejemplo, grandes números de células hepáticas quedan dañadas, como en la hepatitis aguda, o reemplazadas por tejido conjuntivo (como en la cirrosis ). Los pocos defectos genéticos de las enzimas del ciclo del ácido cítrico que se han informado se relacionan con daño neurológico grave
como resultado de alteración muy considerable de la formación de ATP en el sistema nervioso central. La hiperamonemia, como ocurre en la enfermedad hepática avanzada, lleva a pérdida del conocimiento, coma y convulsiones como resultado de actividad alterada del ciclo del ácido cítrico, lo que lleva a formación reducida de ATP. El amoníaco disminuye intermediarios del ciclo del ácido cítrico (al retirar el a-ceto- glutarato para la formación de glutamato y glutamina), e inhibe también la descarboxilación oxidativa del a-cetoglutarato.
El cIclo dEl ácIdo cítrIco
proporcIona sustrato
para la cadEna rEspIratorIa
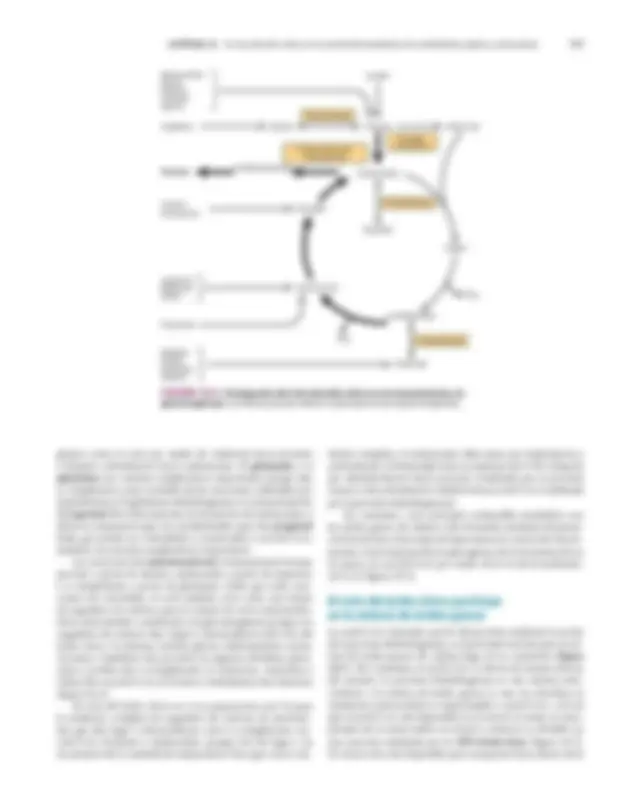
El ciclo empieza con la reacción entre la porción acetilo de la acetil-CoA y el ácido dicarboxílico de cuatro carbonos oxaloa- cetato, lo que forma un ácido tricarboxílico de seis carbonos, el citrato. En las reacciones subsiguientes, se liberan dos molécu- las de CO 2 , y se regenera el oxaloacetato ( figura 16-1 ). Sólo se
162 SEccIón IV Metabolismo de carbohidratos
requiere una pequeña cantidad de oxaloacetato para la oxida- ción de una gran cantidad de acetil-CoA; puede considerarse que desempeña una función catalítica , ya que es regenerada al final del ciclo. El ciclo del ácido cítrico proporciona la principal vía para la formación de ATP enlazada a la oxidación de combustibles metabólicos. Durante la oxidación de acetil-CoA, las coenzimas se reducen y después se reoxidan en la cadena respiratoria, enla- zadas a la formación de ATP (fosforilación oxidativa, figura 16-2 ; capítulo 13). Este proceso es aerobio ; requiere oxígeno como el oxidante final de las coenzimas reducidas. Las enzimas del ciclo del ácido cítrico están ubicadas en la matriz mitocondrial , libres o fijas a la membrana mitocondrial interna y la membrana de las crestas, donde también se encuentran las enzimas y coenzimas de la cadena respiratoria (capítulo 13).
las rEaccIonEs dEl cIclo
dEl ácIdo cítrIco lIbEran
EquIvalEntEs rEductorEs y co 2
La reacción inicial entre la acetil-CoA y el oxaloacetato para formar citrato está catalizada por la citrato sintasa , que forma un enlace de carbono-carbono entre el carbono metilo de la ace- til-CoA y el carbono carbonilo del oxaloacetato ( figura 16-3 ). El enlace tioéster de la citril-CoA resultante se hidroliza, lo que libera citrato y CoASH —una reacción exotérmica—. La enzima aconitasa (aconitato hidratasa) isomeriza el citrato hacia isocitrato; la reacción ocurre en dos pasos: deshi- dratación hacia cis-aconitato, y rehidratación hacia isocitrato. Aun cuando el citrato es una molécula simétrica, con la aconi- tasa reacciona de manera asimétrica, de modo que los dos áto- mos de carbono que se pierden en reacciones subsiguientes del ciclo no son los que se añadieron provenientes de la acetil-CoA. Tal conducta asimétrica depende de canalización : transferencia del producto de la citrato sintasa de manera directa hacia el sitio activo de la aconitasa, sin entrar en solución libre. Esto propor- ciona integración de la actividad del ciclo del ácido cítrico y el suministro de citrato en el citosol como una fuente de acetil-CoA
para la síntesis de ácido graso. El citrato sólo está disponible en solución libre para ser transportado desde las mitocondrias hacia el citosol para la síntesis de ácidos grasos cuando la aconitasa es inhibida por la acumulación de su producto, el isocitrato. El veneno fluoroacetato se encuentra en algunos vegetales y su consumo puede ser mortal para animales que pastan. Algunos compuestos fluorados que se usan como agentes anticáncer y como sustancias químicas industriales (incluso plaguicidas) se metabolizan hacia fluoroacetato. El veneno fluoroacetato es tóxico porque la fluoroacetil-CoA se condensa con oxaloacetato para formar fluorocitrato, que inhibe la aconitasa, lo que hace que se acumule citrato.
Oxaloacetato (C 4 )
Citrato (C 6 )
Acetil-CoA (C 2 ) CoA
CO (^2) CO 2
FIgura 16-1 El ciclo del ácido cítrico, que ilustra el papel catalí- tico del oxaloacetato.
-Cetoglutarato
Ciclo del ácido cítrico
Oxaloacetato (C 4 )
H 2 O Citrato (C 6 ) Cis-aconitato (C 6 )
Isocitrato (C 6 )
α (C 5 )
Succinil-CoA (C 4 )
Succinato (C 4 )
Fumarato (C 4 )
Malato (C 4 )
H 2 O
H 2 O
H 2 O
CO (^2)
2H CO (^2)
2H
NAD
2H
2H Fp
Q
Cit b
Cit c
Cit aa (^3) O (^2)
ATP ADP
H 2 O
Acetil-CoA (C 2 )
Carbohidrato Proteína Lípidos
(^1) / 2 Anaerobiosis (hipoxia, anoxia)
Fosforilación oxidativa
Flavoproteína
Cadena respiratoria Fp Cit Citocromo
FIgura 16-2 El ciclo del ácido cítrico: la principal vía catabólica para la acetil-coa en organismos aeróbicos. la acetil-CoA, el pro- ducto del catabolismo de carbohidratos, proteínas y lípidos, es captada hacia el ciclo y oxidada hacia Co 2 , con la liberación de equivalentes reductores (2H). la oxidación subsiguiente de 2H en la cadena respira- toria lleva a fosforilación de ADP hacia ATP. Para una vuelta del ciclo, se generan nueve ATP por medio de fosforilación oxidativa, y surge un ATP (o GTP) en el ámbito de sustrato a partir de la conversión de succi- nil-CoA en succinato.
164 SEccIón IV Metabolismo de carbohidratos
fisiológico. Como sucede con la oxidación del piruvato (capí- tulo17), la arsenita inhibe la reacción, lo que hace que se acumule el sustrato, a -cetoglutarato. Altas concentraciones de amoniaco inhiben la deshidrogenasa de a-cetoglutarato. La succinil-CoA se convierte en succinato mediante la enzima succinato tiocinasa (succinil-CoA sintetasa) ; se trata del único ejemplo en el ciclo del ácido cítrico de fosforilación en el ámbito de sustrato. Los tejidos en los cuales ocurre gluconeogénesis (el hígado y los riñones) contienen dos isoenzimas de succinato tiocinasa, una específica para difosfato de guanosina (guanosín difosfato; GDP) y la otra para ADP. El trifosfato de guanosina (guanosín trifosfato; GTP) formado se usa para la descarboxila- ción de oxaloacetato hacia fosfoenolpiruvato en la gluconeogéne- sis, y proporciona un enlace regulador entre la actividad del ciclo del ácido cítrico y el retiro de oxaloacetato para la gluconeogéne- sis. Los tejidos no gluconeogénicos sólo tienen la isoenzima que usa ADP. Cuando los cuerpos cetónicos se están metabolizando en tejidos extrahepáticos, hay una reacción alternativa catalizada por la succinil-CoA-acetoacetato-CoA transferasa (tioforasa), que comprende transferencia de CoA desde la succinil-CoA hacia el acetoacetato, lo que forma acetoacetil-CoA y succinato (capítulo 22). El metabolismo anterógrado de succinato, que lleva a la rege- neración de oxaloacetato, es la misma secuencia de reacciones químicas que ocurre en la b-oxidación de ácidos grasos: deshi- drogenación para formar un doble enlace de carbono-carbono, adición de agua para formar un grupo hidroxilo, y deshidrogena- ción adicional para dar el grupo oxo del oxaloacetato. La primera reacción de deshidrogenación, que forma fuma- rato, es catalizada por la succinato deshidrogenasa , que está unida a la superficie interna de la membrana mitocondrial interna. La enzima contiene FAD y proteína hierro-azufre (Fe:S), y reduce de manera directa la ubiquinona en la cadena de trans- porte de electrones. La fumarasa (fumarato hidratasa) cata- liza la adición de agua a través del doble enlace del fumarato, lo que produce malato. La malato deshidrogenasa convierte a este último en oxaloacetato, una reacción que requiere NAD^1. Aunque el equilibrio de esta reacción favorece con fuerza al malato, el flujo neto es hacia el oxaloacetato debido a la elimina- ción continua de este último (para formar citrato, como un sus- trato para la gluconeogénesis, o para pasar por transaminación hacia aspartato), y a la reoxidación continua de NADH.
por cada vuElta dEl cIclo
dEl ácIdo cítrIco sE Forman
10 atp
Como resultado de oxidaciones catalizadas por las deshidroge- nasas del ciclo del ácido cítrico, se producen tres moléculas de NADH y una de FADH 2 por cada molécula de acetil-CoA cata- bolizada en una vuelta del ciclo. Estos equivalentes reductores se transfieren hacia la cadena respiratoria (figura 13-3), donde la reoxigenación de cada NADH origina la formación de ~2. ATP, y de FADH 2 , ~1.5 ATP. Además, 1 ATP (o GTP) se forma mediante fosforilación en el ámbito de sustrato catalizada por la succinato tiocinasa.
las vItamInas dEsEmpEñan
FuncIonEs clavE En El cIclo
dEl ácIdo cítrIco
Cuatro de las vitaminas B (capítulo 44) son esenciales en el ciclo del ácido cítrico y, por ende, en el metabolismo productor de energía: la riboflavina , en forma de flavina adenina dinucleótido (FAD), un cofactor para la succinato deshidrogenasa; niacina , en forma de nicotinamida adenina dinucleótido (NAD), el acep- tor de electrón para la isocitrato deshidrogenasa, a-cetoglutarato deshidrogenasa, y malato deshidrogenasa; tiamina ( vitamina B 1 ), como difosfato de tiamina, la coenzima para la descarboxilación en la reacción de a-cetoglutarato deshidrogenasa, y ácido pan- toténico , como parte de la coenzima A, el cofactor fijo a residuos ácido carboxílicos “activos” como acetil-CoA y succinil-CoA.
El cIclo dEl ácIdo cítrIco
dEsEmpEña una FuncIón
crucIal En El mEtabolIsmo
El ciclo del ácido cítrico no sólo es una vía para la oxidación de unidades de dos carbonos, sino que también es una vía impor- tante para la interconversión de metabolitos que surgen por transaminación y desaminación de aminoácidos (capítulos 28 y 29), y proporciona los sustratos para la síntesis de aminoácidos mediante transaminación (capítulo 27), así como para gluconeo- génesis (capítulo 20) y síntesis de ácidos grasos (capítulo 23). Dado que funciona en procesos tanto oxidativos como sintéticos, es anfibólico ( figura 16-4 ).
El ciclo del ácido cítrico participa en la gluconeogénesis, transaminación y desaminación Todos los intermediarios del ciclo son en potencia glucogénicos , porque pueden dar lugar a oxaloacetato y, por ende, a producción neta de glucosa (en hígado y riñones, los órganos que llevan a cabo la gluconeogénesis; capítulo 19). La enzima clave que cataliza la transferencia neta hacia afuera del ciclo hacia la gluconeogénesis es la fosfoenolpiruvato carboxicinasa , la cual cataliza la descar- boxilación de oxaloacetato hacia fosfoenolpiruvato; el GTP actúa como el donador de fosfato (figura 19-1). El GTP que se requiere para esta reacción es proporcionado (en el hígado y los riñones) por la isoenzima dependiente de GDP de la succinato tirosinasa. Esto asegura que el oxaloacetato no se retire del ciclo para gluco- neogénesis si esto llevaría a la disminución de intermediarios del ciclo del ácido cítrico y, por ende, generación reducida de ATP. La transferencia neta hacia el ciclo ocurre como resultado de varias reacciones. Entre las más importantes de esas reaccio- nes anapleróticas , está la formación de oxaloacetato mediante la carboxilación de piruvato, catalizada por la piruvato carboxilasa ( figura 16-4 ). Esta reacción es importante para mantener una concentración adecuada de oxaloacetato para la reacción de con- densación con acetil-CoA. Si esta última se acumula, actúa como activador alostérico de la piruvato carboxilasa y como inhibidor de la piruvato deshidrogenasa, lo que asegura un aporte de oxa- loacetato. El lactato, un importante sustrato para la gluconeo-
capíTulO 16 El ciclo del ácido cítrico: la vía central del metabolismo de carbohidratos, lípidos y aminoácidos 165
génesis, entra al ciclo por medio de oxidación hacia piruvato y después carboxilación hacia oxaloacetato. El glutamato y la glutamina son sustratos anapleróticos importantes porque dan a-cetoglutarato como resultado de las reacciones catalizadas por la glutaminasa y la glutamato deshidrogenasa. La transaminación de aspartato lleva directamente a la formación de oxaloacetato, y diversos compuestos que son metabolizados para dar propionil CoA , que puede ser carboxilada e isomerizada a succinil CoA, también son sustratos anapleróticos importantes. Las reacciones de aminotransferasa (transaminasa) forman piruvato a partir de alanina, oxaloacetato a partir de aspartato, y a-cetoglutarato a partir de glutamato. Dado que estas reac- ciones son reversibles, el ciclo también sirve como una fuente de esqueletos de carbono para la síntesis de estos aminoácidos. Otros aminoácidos contribuyen a la gluconeogénesis porque sus esqueletos de carbono dan origen a intermediarios del ciclo del ácido cítrico. La alanina, cisteína, glicina, hidroxiprolina, serina, treonina y triptófano dan piruvato; la arginina, histidina, gluta- mina y prolina dan a-cetoglutarato; la isoleucina, metionina y valina dan succinil-CoA; la tirosina y fenilalanina dan fumarato (figura 16-4). El ciclo del ácido cítrico en sí no proporciona una vía para la oxidación completa de esqueletos de carbono de aminoáci- dos que dan lugar a intermediarios como a-cetoglutarato, suc- cinil-CoA, fumarato y oxaloacetato, porque esto da lugar a un incremento de la cantidad de oxaloacetato. Para que ocurra oxi-
dación completa, el oxaloacetato debe pasar por fosforilación y carboxilación a fosfoenolpiruvato (a expensas de GTP) y después por desfosforilación hacia piruvato (catalizada por la piruvato cinasa) y descarboxilación oxidativa hacia acetil CoA (catalizada por la piruvato deshidrogenasa). En rumiantes, cuyo principal combustible metabólico son los ácidos grasos de cadena corta formados mediante fermenta- ción bacteriana, tiene especial importancia la conversión de pro- pionato, el principal producto glucogénico de la fermentación en el rumen, en succinil-CoA por medio de la vía de la metilmalo- nil-CoA (figura 19-2).
El ciclo del ácido cítrico participa en la síntesis de ácidos grasos La acetil-CoA, formada a partir del piruvato mediante la acción de la piruvato deshidrogenasa, es el principal sustrato para la sín- tesis de ácidos grasos de cadena larga en no rumiantes ( figura 16-5 ). (En rumiantes, la acetil-CoA se deriva de manera directa del acetato). La piruvato deshidrogenasa es una enzima mito- condrial, y la síntesis de ácidos grasos es una vía citosólica; la membrana mitocondrial es impermeable a acetil-CoA; a fin de que el acetil-CoA esté disponible en el citosol, el citrato es trans- portado de la mitocondria al citosol y entonces es dividido en una reacción catalizada por la ATP-citrato liasa (figura 16-5). El citrato sólo está disponible para transporte hacia afuera de la
Hidroxiprolina Serina Cisteína Treonina Glicina
Lactato
Triptófano Alanina Piruvato Acetil-CoA
CO 2
CO 2
Citrato
Aspartato
a-Cetoglutarato
Glutamato
Transaminasa
Transaminasa
Transaminasa
Succinil-CoA
Fumarato
Glucosa Oxaloacetato
Tirosina Fenilalanina
Isoleucina Metionina Valina
Propionato
Histidina Prolina Glutamina Arginina
Fosfoenolpiruvato
Fosfoenolpiruvato carboxicinasa
Piruvato carboxilasa
FIgura 16-4 participación del ciclo del ácido cítrico en la transaminación y la gluconeogénesis. las flechas gruesas indican la principal vía de la gluconeogénesis.
capíTulO 16 El ciclo del ácido cítrico: la vía central del metabolismo de carbohidratos, lípidos y aminoácidos 167
deshidrogenasa y a-cetoglutarato deshidrogenasa. Las deshidro- genasas son activadas por Ca^21 , que aumenta de concentración durante la contracción muscular y secreción por otros tejidos, cuando hay aumento de la demanda de energía. En un tejido como el cerebro, que depende en su mayor parte de carbohidratos para el aporte de acetil-CoA, el control del ciclo del ácido cítrico puede ocurrir en la piruvato deshidrogenasa. Varias enzimas se encargan de la situación en cuanto a energía, como se demuestra por las proporciones [ATP]/[ADP] y [NADH]/[NAD^1 ]. De este modo, hay inhibición alostérica de la citrato sintasa por el ATP y acil-CoA grasa de cadena larga. La activación alostérica de la iso- citrato deshidrogenasa dependiente de NAD, mitocondrial, por ADP, es contrarrestada por ATP y NADH. El complejo de a-ce- toglutarato deshidrogenasa está regulado de la misma manera que la piruvato deshidrogenasa (figura 17-6). La succinato des- hidrogenasa es inhibida por el oxaloacetato, y la disponibilidad de este último, según se controla por la malato deshidrogenasa, depende de la proporción [NADH]/[NAD^1 ]. Dado que la Km para oxaloacetato de la citrato sintasa es del mismo orden de magnitud que la concentración intramitocondrial, es probable que la concentración de oxaloacetato controle el índice de for- mación de citrato. La hiperamonemia, como ocurre en la enfermedad hepática avanzada y en varias enfermedades genéticas (raras) del metabo- lismo de aminoácidos, lleva a pérdida del conocimiento, coma y convulsiones, y puede ser mortal. Esto se debe mayormente al retiro de a-cetoglutarato para formar glutamato (lo cual es catalizado por la glutamato deshidrogenasa) y después gluta- mina (lo cual es catalizado por la glutamina sintetasa), lo que da pie a concentraciones reducidas de todos los intermediarios del ciclo del ácido cítrico y, por ende, menor generación de ATP. El equilibrio de la glutamato deshidrogenasa está finamente esta- blecido, y la dirección de la reacción depende de la proporción de NAD^1 :NADH y la concentración de iones amonio. Además, el amoníaco inhibe la a-cetoglutarato deshidrogenasa, y posible- mente también la piruvato deshidrogenasa.
rESumEn
■ ■ (^) El ciclo del ácido cítrico es la vía final para la oxidación de carbohidratos, lípidos y proteínas. Su metabolito terminal común, la acetil-CoA, reacciona con el oxaloacetato para formar citrato. Mediante una serie de deshidrogenaciones y descarboxilaciones, el citrato es degradado, lo que reduce coenzimas, libera dos CO 2 y regenera oxaloacetato. ■ ■ (^) Las coenzimas reducidas se oxidan mediante la cadena respiratoria enlazada a la formación de ATP. De este modo, el ciclo es la principal vía para la formación de ATP, y está ubicado en la matriz de mitocondrias adyacente a las enzimas de la cadena respiratoria y la fosforilación oxidativa.
■ ■ (^) El ciclo del ácido cítrico es anfibólico, puesto que además de oxidación, es importante en el suministro de esqueletos de carbono para la gluconeogénesis, la síntesis de ácidos grasos y la interconversión de aminoácidos.
rEFErEncIaS
Baldwin JE, Krebs HA: The evolution of metabolic cycles. Nature 1981;291:381. Bender DA: The metabolism of “surplus” amino acids. Br J Nutr 2012;108(suppl 2): S113. Bowtell JL, Bruce M: Glutamine: an anaplerotic precursor. Nutrition 2002;18:222. Briere JJ, Favier J, Giminez-Roqueplo A-P, et al: Tricarboxylic acid cycle dysfunction as a cause of human diseases and tumor formation. Am J Physiol Cell Physiol 2006;291:C1114. Brunengraber H, Roe CR: Anaplerotic molecules: current and future. J Inherit Metab Dis 2006;29:327. De Meirleir L: Defects of pyruvate metabolism and the Krebs cycle. J Child Neurol 2002;(suppl 3):3S26. Depeint F, Bruce WR: Mitochondrial function and toxicity: role of the B vitamin family on mitochondrial energy metabolism. Chem Biol Interact 2006;163:94. Gibala MJ, Young ME: Anaplerosis of the citric acid cycle: role in energy metabolism of heart and skeletal muscle. Acta Physiol Scand 2000;168:657. Grunengraber H, Roe CR: Anaplerotic molecules: current and future. J Inherit Metab Dis 2006;29:327. Hertz L, Kala G: Energy metabolism in brain cells: effects of elevated ammonia concentrations. Metab Brain Dis 2007; 22:199–218. Jitrapakdee S, St Maurice M, Rayment I, et al: Structure, mechanism and regulation of pyruvate carboxylase. Biochem J 2008;413:369. Jitrapakdee S, Vidal-Puig A, Wallace JC: Anaplerotic roles of pyruvate carboxylase in mammalian tissues. Cell Mol Life Sci 2006;63:843. Kay J, Weitzman PDJ (editors): Krebs’ Citric Acid Cycle—Half a Century and Still Turning. Biochemical Society, 1987. Kornberg H: Krebs and his trinity of cycles. Nat Rev Mol Cell Biol 2000;1:225. Ott P, Clemmesen O, Larsen FS: Cerebral metabolic disturbances in the brain during acute liver failure: from hyperammonemia to energy failure and proteolysis. Neurochem Int 2005;47:13. Owen OE, Kalhan SC: The key role of anaplerosis and cataplerosis for citric acid cycle function. J Biol Chem 2002;277:30409. Pithukpakorn M: Disorders of pyruvate metabolism and the tricarboxylic acid cycle. Mol Genet Metab 2005;85:243. Proudfoot AT, Bradberry SM: Sodium fluoroacetate poisoning. Toxicol Rev 2006;25:2139. Rama Rao KV, Norenberg MD: Brain energy metabolism and mitochondrial dysfunction in acute and chronic hepatic encephalopathy. Neurochem Int 2012;60:697. Sumegi B, Sherry AD: Is there tight channelling in the tricarboxylic acid cycle metabolon? Biochem Soc Trans 1991;19:1002.