¡Descarga Mitochondrias y ADN: Transcripción, Sinapsis y Herencia y más Apuntes en PDF de Biología solo en Docsity!
INTRODUCCIÓN
Araceli Lantigua Cruz Mencionar la relación de hechos históricos que han confluido desde el redescubrimiento de las leyes de Mendel hasta nuestros días, requeriría de lar- gas horas de búsqueda. Cada aporte científico ha sido un importante eslabón en esta larga cadena que nos lleva desde la Genética General hasta la Genética Clínica actual y que sin dudas ha tenido un impacto impresionante en las Cien- cias Médicas. En este primer capítulo exponemos un cuadro que menciona apenas pincela- das sobre hechos relevantes que han dejado una impronta en el desarrollo actual de las disciplinas de las Genética Humana, Médica y Clínica.
ANTECEDENTES
Es práctica habitual comenzar los recuentos sobre la historia de la Genética con los trabajos de Mendel a mediados del siglo XIX y su redescubrimiento en los inicios del siglo XX. Sin embargo, observaciones sobre la herencia biológica en humanos habían sido realizadas desde el siglo XVII, aunque no tuvieron ni el significado ni la trascendencia de los trabajos de Mendel, pues el estudio de la Genética Humana presenta cierto grado de complejidad. Estos intentos pioneros estuvieron mejor encaminados ya a principios del siglo XIX. Como muestra de ello se mencionan los siguientes. En el año 1814 Joseph Adams, médico inglés, publicó un libro en el que se destacan las siguientes observaciones:
- Diferencias entre congénito, familiar y condiciones hereditarias.
- Las enfermedades hereditarias no están presentes al nacimiento sino que se manifies- tan por si mismas a diferentes edades.
- Hay predisposiciones para enfermedades que determinan que estas solo se manifies- ten bajo la influencia de factores ambientales. 1
- La reproducción de personas afectadas puede estar disminuida y propuso establecer registros de familias con enfermedades hereditarias. Qué no hubiera hecho este científico con la automatización actual de datos. El profesor alemán de medicina C. F. Nasse hizo el reconocimiento en el año 1820 de la transmisión de la hemofilia a través de mujeres. Debe tenerse presente que en el siglo XIX el nivel de reconocimiento de una enfer- medad genética en el humano se basaba solamente en el análisis clínico de los signos y síntomas de enfermedades, el reconocimiento del carácter familiar y la historia de la enfermedad, en especial la edad de comienzo de los primeros síntomas. Sin embargo, Mendel hizo sus experimentos con guisantes de líneas puras lo cual le permitió llegar a sus trascendentales conclusiones. Estas no fueron comprendidas por las limitaciones de conocimientos del momento. Durante las últimos décadas del siglo XIX se producen algunos descubrimientos im- portantes. En 1875 Hertwig observó la fertilización animal y la continuidad de células nucleadas. Cinco años después Fleming observa y descubre las cromátidas hermanas de los cromosomas en la mitosis. Ya en 1883, von Beneden establece la regularidad de los cromosomas en los núcleos de células hijas. Y para 1888 Boveri establece la individuali- dad de cada par cromosómico y en ese mismo año Waldeyer utiliza el término cromosoma (cuerpo que toma color) por las características de tinción de estas estructuras celulares. También para ese año habían culminados los estudios sobre la mitosis. Todos estos acon- tecimientos hicieron posible la rápida aceptación de los trabajos de Mendel cuando fue- ron redescubiertos en los albores del siglo XX. A partir del redescubrimiento de las leyes de Mendel comienzan a emerger nuevos conocimientos sobre la genética de caracteres humanos que vertiginosamente finalizaron el sigo XX con el proyecto Genoma Humano. En el año 1902 Archibald Garrod publicó un trabajo titulado "La incidencia de la alcaptonuria: Un estudio en individualidad química". Con este trabajo se da inicio al aná- lisis de las leyes de Mendel en humanos e hizo las siguientes observaciones:
- Una persona tiene o no alcaptonuria, son dos alternativas claras.
- El defecto metabólico está presente desde el nacimiento.
- Se observa en hermanos, no en los padres.
- Los padres con frecuencia son primos hermanos.
- Existen, además de la alcaptonuria, otros defectos de este tipo, como el albinismo que puede estar en esta categoría. En el año 1900 Landesteiner descubre el sistema de grupos sanguíneos ABO y en 1911 se comprueba su herencia mendeliana.
El año 1956 marca el inicio de la Genética Clínica ya que paralelo al desarrollo de la citogenética se producen nuevos descubrimientos de defectos metabólicos y se produce un importante avance en la Genética Bioquímica. Entre ellos se destaca el descubrimien- to del defecto bioquímico de la sicklemia por Pauling y sus colaboradores en el año 1949. En 1953 se produjo un hecho trascendental no sólo para la Genética sino para toda la Biología, cuando aparece el modelo molecular del ADN propuesto por Watson y Crick. A partir de ese momento el gen dejó de ser sólo una intuición para tomar materialidad en una molécula específica. Este trabajo, considerado por muchos, como la hipótesis más brillante de la Biología contemporánea, no sólo aportó el conocimiento sobre la estructura del ADN sino que además dejó demostrado fehacientemente que esta molécula era la portadora material de la información genética. La última década del siglo XX ha desbordado la imaginación en recursos técnicos, automatización, nuevos conocimientos, nuevas posibilidades para personas afectadas, familiares y para la sociedad. La manipulación del Genoma Humano ha requerido incor- porar a la Etica Médica principios Bioéticos, .surgidos por la necesidad de tomar decisio- nes que no dañen la integridad de nuestra propia especie. El futuro de las Genética Humana, Médica y Clínica es difícil de predecir. La incerti- dumbre sobre nuevos tratamiento y utilización de nuevos fármacos dirigidos a enfermos con genotipos específicos, bajo el control de las grandes industrias farmacéuticas, pone en peligro el principio ético de la justicia, pues los recursos financieros para la producción de estos fármacos es muy elevada y habría que preguntarse que personas con un tipo específico de defecto genético tendrían la posibilidad de ser tratados. ¿Qué nuevos recursos técnicos se inventarán? ¿Qué nuevos conocimientos surgirán? ¿Beneficiarán o perjudicarán esos nuevos conocimientos a las poblaciones del Tercer Mundo? Una verdad se abre, para contribuir en lo adelante al desarrollo del Proyecto Genoma Humano, es necesario tener presente que el punto de partida de ahora en lo adelante es conocer la epidemiología de lo que es raro y tener el consentimiento de estudio de las personas afectadas a fin de dar la respuesta a ¿por qué es raro? y para eso se requiere del desarrollo de la Genética Comunitaria, que será el pilar del futuro desarrollo de las Genética Humana y Médica y que dará respuesta a las exigencias de diagnóstico, tratamiento, prevención y pronóstico que comprometen al desarrollo de la Genética Clíni- ca. Las Ciencias Médicas no escapan a tan elevado volumen de información y de exigencias prácticas tanto a nivel preclínico como a nivel clínico. Es necesario preparar- se para enfrentar un futuro insospechado en el campo de la Genética del siglo XXI y la repercusión que es de esperar en la atención médica.
PERINATALES INFECCIONES ECTOS CONGÉNITOS 3 2 1 1970 1980 2001 2002 DEF 10 X 1000 15
LAS ENFERMEDADES GENÉTICAS
Los datos que ofrece la epidemiología de las enfermedades genéticas y defectos congénitos se caracterizan por su estabilidad en el tiempo en regiones específicas, Sus variaciones obedecen fundamentalmente a la edad del grupo poblacional que se estudie ya que muchos defectos genéticos se ponen de manifiesto a diferentes edades o causan mortalidad en diferentes periodos de la vida, pero la tendencia una vez realizados los estudios y conocida su epidemiología es a permanecer con prevalencias similares durante años. Solamente varían si se producen factores ambientales que cambien la tasa de nuevas mutaciones o por la ocurrencia de otros factores que pueden estudiarse en el capítulo16; de ahí la importancia de los registros de enfermedades genéticas y de defec- tos genéticos ya recomendados por científicos desde el siglo XIX. Ocurre con frecuencia que dentro de los indicadores de mortalidad infantil se miden los defectos congénitos, pero las frecuencias de estos, cuando hay otros factores ambientales que elevan las causas de mortalidad, impiden reconocer exactamente que las variaciones en las frecuencias de los defectos congénitos son casi constantes. Es por eso que cuando se trata en el tema de Salud, el análisis epidemiológico los defectos congénitos no alcanzan a identificarse como un problema principal cuando existen otros con frecuencia mucho más elevadas. Solamente después de resolver la prevención de la mortalidad a expensas de enfermedades infecciosas y perinatales y de disminuir la incidencia de éstas, es que los defec- tos congénitos manifiestan un aumento relativo de ellas que pueden incluso causar alarma. Un ejemplo puede apreciarse en el gráfico en el que se analizan los tres primeros facto- res de mortalidad en Cuba en el primer año de vida desde el período de 1970 al 2002
PANORAMA DE LA BIOLOGÌA
CELULAR Y MOLECULAR
Rolando Hernández Fernández En 1953 se produce un hecho trascendental en la historia de la Biología, cuan- do la revista Nature publicó el trabajo sobre estructura del ADN de James D. Watson y Francis H. C. Crick. Este trabajo dio comienzo a una revolución en el campo de la Biología y marcó el inicio de la Biología Molecular. Desde entonces hasta nuestros días el conocimiento acerca de los mecanismos moleculares relacio- nados con la herencia biológica han alcanzado un desarrollo insospechado en épocas anteriores. El conocimiento de la estructura molecular del ADN ha permitido esclarecer los mecanismos relacionados con la duplicación del ADN, la síntesis de los ARN y de las proteínas, la recombinación genética y la mutagénesis, así como los complejos mecanismos que permiten la autorregulación de todos esos procesos. Por si esto fuera poco, a partir de ese descubrimiento se han desarrollado pode- rosos procedimientos experimentales que han rebasado el marco de la genética molecular y han incidido de forma fundamental en el conocimiento de la estructura y las funciones celulares, de los procesos del desarrollo filo y ontogenético, de la rela- ción entre el genoma y el ambiente y de las bases moleculares de las enfermedades. En este capítulo nos proponemos hacer un resumen de los principales aspectos de la Biología Celular y Molecular como una introducción panorámica para el resto de los capítulos del libro.
LA BIOLOGÍA CELULAR
La Biología Celular contemporánea tiene como objetivo explicar las funciones de las células a partir del conocimiento de la estructura, las propiedades y las funciones de las moléculas que la forman, especialmente de los agregados supramoleculares. Desde ese punto de vista la célula eucarionte se presenta como un complejo sistema biomolecular con un alto grado de organización estructural y funcional que constituye la unidad básica de los organismos superiores. Sus estructuras están formadas por agrupaciones de molé- culas o de macromoléculas que forman verdaderos organitos en los cuales se llevan a
cabo todas las funciones celulares de una forma armónica y coordinada. Las moléculas que componen estos organitos son los lípidos, las proteínas, los ácidos nucleicos y los polisacáridos. Dos aspectos destacan en estas células, la existencia del núcleo donde se encuentra confinado el ADN y la existencia de una basta red de estructuras membranosas que dividen la célula en numerosos compartimentos, de manera que los procesos se produzcan con relativa independencia uno de otros. Así, para realizar funciones comple- jas es necesario que se genere un flujo de sustancia, energía e información no sólo entre la célula y su entorno sino además entre los diferentes compartimentos intracelulares.
La membrana plasmática
La membrana plasmática es una estructura laminar que rodea a la célula separándola y comunicándola con el exterior. Está compuesta por lípidos, polisacáridos y proteínas. La estructura básica está formada por una doble capa de moléculas de lípidos que son los fosfátidos de glicerina, los esfingolípidos y el colesterol. Estos lípidos presentan una es- tructura anfipática, es decir, contiene una zona polar pequeña y una zona apolar mucha más grande. En las membranas las zonas apolares se disponen hacia el interior y las zonas polares hacia el exterior, una en contacto con el espacio extracelular y otra hacia el interior de la célula. Hasta hace poco se daba por sentado que los lípidos de la membrana solamente tenían un papel pasivo como parte de la barrera que separa la célula del exte- rior, sin embargo, recientemente se ha descubierto que los componentes lipídicos de la membrana intervienen en complejos mecanismos de comunicación intercelular, unos ge- nerando segundos mensajeros (como el diacilglicerol), otros regulando la actividad de enzimas (como el fosfatitil inositol) y aún unos terceros como precursores de importantes señales moleculares (como el ácido araquidónico que da origen a las prostaglandinas) que comunican a una célula con sus vecinas. Los lípidos además participan en los meca- nismos de difusión a través de la membrana de sustancias apolares y del agua. Los polisacáridos suelen ser cortos (catorce a veinte unidades) y ramificados, con monosacáridos derivados (galactosamina, fucosa, glucosamina, etc.) y siempre están orientados hacia el espacio extracelular. Participan en funciones de reconocimiento intercelular (como los grupos sanguíneos) y se encuentran unidos bien a los lípidos, bien a las proteínas. Las proteínas son los componentes funcionales más importantes de las membranas. Las funciones específicas que una membrana realiza se deben a las proteínas que contie- ne. Las proteínas pueden ser extrínsecas o periféricas si sólo están en contacto con la parte polar de la membrana e intrínsecas o integrales si lo hacen con la zona apolar, entre las que se encuentran las transmembranales que atraviesan la membrana de un lado al otro. Las proteínas pueden constituir poros por donde pasan sustancias polares cuyo
La envoltura nuclear está formada por dos membranas, la externa cubierta temporal- mente de ribosomas y la interna asociada con la lámina nuclear. Estas membranas se fusionan en numerosos puntos dejando una abertura cuyas paredes están cubiertas de proteínas dando lugar al llamado complejo del poro nuclear por donde transitan macromoléculas, tanto desde el núcleo hacia el citoplasma, como en sentido contrario. El aparato de Golgi está formado por un conjunto de sacos membranosos aplanados que está muy desarrollado en las células secretoras. En su interior las proteínas sintetizadas por los ribosomas unidos al retículo endoplásmico rugoso son modificadas, principalmente por glicosilaciones, concentradas y empaquetadas para ser enviadas a diferentes lugares, entre ellos los lisosomas, los peroxisomas y la membrana plasmática. Los organitos citoplasmáticos membranosos son las mitocondrias, los lisosomas y los peroxisomas. Las mitocondrias están formadas por dos membranas, la externa es lisa y permeable a moléculas de hasta 10 kDa, mientras que la interna forma pliegues hacia el interior llamados crestas y es prácticamente impermeable a casi todas las sustancias con excepción del agua, el oxígeno y el dióxido de carbono. El espacio limitado por la mem- brana interna recibe el nombre de matriz y es el asiento de importantes procesos metabólicos principalmente del Ciclo de Krebs. La membrana interna contiene cerca de un 70% de proteínas entre las cuales se encuentran las que forman parte de la cadena transportado- ra de electrones y la ATP sintetasa que cataliza la formación del ATP. Muchas de las otras proteínas actúan como transportadores, lo cual es necesario debido a la poca per- meabilidad de la membrana. Los lisosomas contienen un gran número de enzimas hidrolíticas capaces de degradar un gran número de sustancias complejas por lo que se han identificado como el sistema digestivo de la célula. Su membrana contiene un bomba de protones que permite mante- ner en el interior un pH más bajo que en el exterior. Como la enzimas tienen su mayor actividad a ese pH en caso de ruptura de los lisosomas su contenido sería vertido al citosol, donde el pH más alto inactivaría a las enzimas impidiendo la digestión de los componentes celulares. Los peroxisomas son corpúsculos membranosos redondeados que contienen un buen grupo de enzimas oxidativas, entre ellas la catalasa y la peroxidasa, y otras que participan en la oxidación de los ácidos grasos de cadena muy larga. Se han descrito numerosas enfermedades debidas a la deficiencia genética de mu- chas de las proteínas que forman parte del sistema de endomembranas.
El citoesqueleto
El citoesqueleto forma una especie de armazón de la célula y está integrado básica- mente por tres tipos de estructuras: los microfilamentos, los microtúbulos y los filamentos intermedios. Los microfilamentos se forman por la polimerización de la actina en presen- cia de ATP. Estos filamentos forman una intricada red inmediatamente por debajo de la membrana plasmática y mediante proteínas específicas están en contacto con compo- nentes de la matriz extracelular. Los microfilamentos son estructuras dinámicas. Existe normalmente un equilibrio entre las reacciones de polimerización y despolimerización de la actina que hace posible los movimientos que realiza la membrana plasmática en los movimientos ameboideos y durante el proceso de fagocitosis. Los microtúbulos se forman por la polimerización de la tubulina de la cual existen dos variedades, la y la . Estas forman un dímero que se polimeriza dependiendo de GTP. Los dímeros al polimerizarse dan lugar a una estructura helicoidal que deja una luz central que es el motivo de su nombre. Los microtúbulos se disponene en forma radiada desde el núcleo hacia la periferia de la célula, creando una armazon que da forma y consistencia a la célula. Proteínas que funcionan como motores moleculares se asocian a la superficie de los microtúbulos y transportan sustancias de gran tamaño incluyendo vesículas membranosas y macromoléculas. También constituyen una estructura dinámica que se polimeriza y despolimeriza de acuerdo con las condiciones celulares. Una forma especial de estos microtúbulos es el huso acromático que se forma durante la división celular. Los filamentos intermedios son proteínas fibrilares que se asocian lateralmente y dan a la célula consistencia y resitencia a las tensiones. Mientras la actina y la tubulina son las mismas en todas las etirpes celulares, los filamentos intermedios son específicos de cada tipo, así en los epitelios se encuentran los filamentos de keratina mientas los neurofilamentos se encuentran en el sistema nervioso. Entre los filamentos intermedios se encuentran las lamininas A, B y C que son los componentes moleculares de la lámina nuclear.
Los ribosomas
Los ribosomas no son organitos membranosos pues están formados por ácidos ribonucleicos ribosomales y proteínas. Están formados por dos subunidades de tamaño diferente denominadas L la mayor y S la menor. La subunidad L contiene los ARNr de 28 S, 5,8 S y 5 S y más de cincuenta proteínas. La subunidad S solo contiene el ARNr de 18 S y unas 35 proteínas. Para su funcionamiento requieren además del concurso de un buen número de proteínas no ribosomales. La función de los ribosomas es la traducción genética o sea, la síntesis de proteínas.
EL CICLO CELULAR
En el desarrollo ontogenético de un organismo pluricelular las células más primitivas se multiplican y se van diferenciando de manera que cada célula forma parte de un órgano o tejido especializado. Cada estirpe celular prolifera hasta cierto punto y entonces se detiene. Los órganos sólidos adquieren forma y tamaño fijos y después cesa el creci- miento. En el individuo adulto el crecimiento de muchos tipos celulares transcurre a velo- cidades muy lentas que solo permiten reponer células viejas o dañadas. Al terminar la división celular (fase M) a las células hijas se le presentan dos alterna- tivas, bien entrar en un periodo de reposo (G0) donde la mayor parte de las funciones celulares se expresan a un nivel basal; bien comenzar a prepararse para un nuevo ciclo de replicación (G1). Las células en G1 comienzan su actividad metabólica pero esta puede decrecer si en un momento determinado no son estimuladas por factores de creci- miento del tipo del factor de crecimiento derivado de plaquetas (PDGF)^1 que hace a las células competentes (por eso le llamamos punto C) para continuar su recorrido por G1. Pasado el punto C las células muestran una intensa actividad metabólica. Se incrementa el transporte de nutrientes a través de la membrana. Hay un aumento considerable de la glucolisis y de la síntesis de proteínas. Pero aparece un nuevo punto donde las células deben ser estimuladas por factores como la insulina o el factor insulinoide de crecimiento 1 (IGF?1).^2 Si las células rebasan ese punto continúan su progresión por el ciclo (por eso le denominamos punto P). A partir de aquí se produce un intenso tráfico de proteínas hacia el núcleo, tal vez las que participan en la replicación del ADN. Las interacciones de las células con factores específicos de crecimiento constituyen un control positivo, pues la presencia del factor en el medio estimula la proliferación celular. Los factores de crecimiento influyen sobre la actividad celular porque las células poseen en la membrana plasmática receptores específicos para ellos. Estos receptores son proteínas transmembranales que hacia el lado citoplasmático presentan un dominio enzimático con actividad de tirosil protein kinasa (TPK), es decir, que transfiere grupos fosforilos del ATP hacia proteínas, en las cuales los esterifica con residuos de tirosina. La unión del factor con su receptor específico estimula la actividad de TPK del receptor que en primer lugar se autofosforila y después fosforila a otras proteínas de la membrana y del citosol. Las proteínas fosforiladas por el receptor son, por lo general, kinasas que fosforilan a otras enzimas y así se establece una cadena de fosforilación que comienza en (^1) Las abreviaturas provienen del nombre en inglés, en este caso PDGF: Plateld Derived Growth Factor. (^2) Insulin-like Growth Factor.
la membrana y avanza hacia el núcleo, donde la fosforilación de factores de transcripción activan la expresión de genes específicos, cuyos productos están relacionados con los mecanismos de la proliferación celular. Los estudios de transformación llevados a cabo con virus ADN llevaron a la conclu- sión de que el ciclo celular exhibe igualmente un control negativo. Las células elaboran proteínas cuya función es inhibir la progresión del ciclo. Cuando uno de estos virus infec- ta a una célula, ésta produce proteínas codificadas por el genoma viral. Se ha podido comprobar que algunas de ellas se asocian con proteínas del hospedero y las mantienen inactivas. Libres del freno que significa la actividad de las proteínas inhibidoras las célu- las proliferan incontroladamente. En condiciones normales los mecanismos positivos y negativos forman una intrincada red molecular cuyo funcionamiento armónico permite que el grado de proliferación celu- lar se adapte perfectamente al momento del desarrollo del organismo y a las condiciones ambientales imperantes. Los estudios del ciclo celular en muchos organismos han mostrado que en todos ellos el ciclo celular progresa básicamente por la intervención de un número reducido de fami- lias de proteínas con funciones similares a las de levaduras.
Las ciclinas
Las ciclinas constituyen una familia de proteínas muy diversas de masa molecular relativamente pequeña, 35 a 90 kD, que fueron identificadas y nombradas porque su síntesis fluctúa durante el ciclo celular. Presentan una secuencia de 100 residuos de aminoácidos conocida como motivo de ciclinas. Este motivo es necesario para la unión a, y la activación de las Cdk correspondientes. La concentración intracelular de un tipo particular de ciclina se eleva bruscamente en un momento del ciclo celular y poco tiempo después también disminuye bruscamente. Mucho se ha investigado acerca del sistema regulador capaz de generar esas oscilacio- nes en las concentraciones intracelulares de ciclinas y se han podido identificar al menos dos sitios de regulación: la transcripción génica y la degradación de proteínas. En los mamíferos el conocimiento del control de la transcripción de las ciclinas se limita a la transición entre la fase G1 y la S. La cascada enzimática desencadenada por los factores de crecimiento conduce a la activación de la ERK(^3 ), una proteína de la familia de las MAPK(^4 ). La ERK por medio de la fosforilación produce la activación de (^3) Del inglés E xtracellular signal R egulated protein K inases. (^4) Del inglés M itogen A ctivated P rotein K inases.
Las proteínas kinasas dependientes de ciclina (Cdk)^5
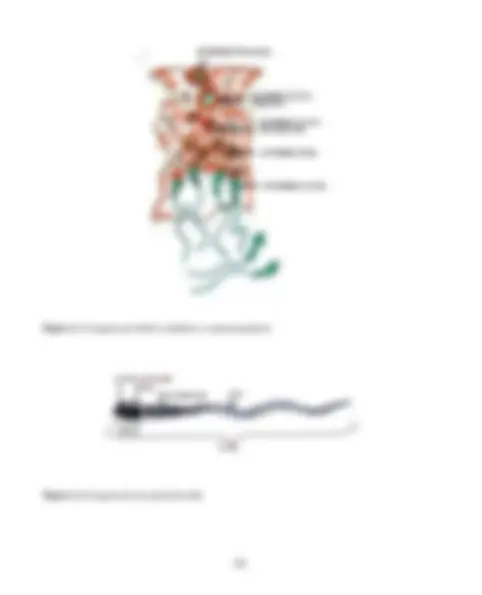
Las protein kinasas dependientes de ciclina (Cdk) son enzimas que transfieren grupos fosforilos del ATP hacia grupos hidroxilos de serina o treonina de las proteínas sustratos. La progresión del ciclo por G1 en los vertebrados requiere la estimulación externa por factores de crecimiento (Figura 2.2). En muchos tipos celulares la respuesta clave a estos factores de cre-cimiento es la activación de la Cdk4 u otra íntimamente relacionada con la Cdk6 las cua-les actúan en asociación con las ciclinas D (D1, D2 y D3). Los complejos Cdk4/Cdk6/ciclina D hacen progresar las células por G1 al suprimir la acción antiproliferante de la proteína pRB, como lo demuestra el hecho de que células carentes de pRB progre- san por G1 en ausencia de Cdk4/ciclina D. F inal d e la m itosis. In icio d e G 1. N o tran scrip ció n. A u m en ta la activ id ad d e C d k 4 /ciclin a D. F o sfo rilación de pR B. B ajo n iv el de tran scrip ció n. T ran sición G 1 - S. A u m en ta la activ idad de C d k 2 /ciclina E. F o sfo rilació n de p R B. A u m en ta el n ivel d e tran scrip ció n. P eríod o S. H iperfo sfo rilació n de pR B , qu e p rovo ca su separació n d el E 2 F. T ran scripció n a n ivel m áx im o. P eríod o S av an zad o. A lta activ idad de C d k2 /ciclina A. F o sfo rilación de E 2 F. S e su p rim e la tran scrip ció n. Figura 2.2. Eventos moleculares que favorecen la progresión del ciclo celular. Más detalles en el textto. (^5) Del inglés C yclin- D ependent K inases.
La Cdk2 se asocia con la ciclina E en la transición de la fase G1 a la fase S y poste- riormente se asocia con la ciclina A haciendo progresar la fase S aunque su papel exacto no está totalmente establecido. La concentración nuclear de la ciclina B se eleva a mediados de la fase G2 y forma complejos con la Cdk1 al cual se ha llamado Factor Promotor de la Mitosis (MPF del inglés mitosis promoting factor). Este complejo es el que fosforila la lámina nuclear en la profase posibilitando la desintegración de la envoltura nuclear. Existen como mínimo cuatro mecanismos principales para la regulación de la activi- dad de las Cdk. El mecanismo primario de activación es la unión con las ciclinas, que se completa en la mayoría de las Cdk por la fosforilación en un residuo específico de treonina de la posición 160. Por otra parte los complejos Cdk/ciclina pueden ser inactivados por al menos dos mecanismos: la unión con proteínas inhibidoras (CDI) y la fosforilación en sitios inhibitorios cerca del extremo N terminal.
Los inhibidores de las Cdk (CDI/CKI)
Todos los mecanismos reguladores de la progresión de un proceso no sólo necesita de factores que lo aceleren sino que también requieren de algunos que lo frenen o retarden. El ciclo celular también posee esos frenos moleculares que son los inhibidores de las kinasas dependientes de ciclina (CDI o CKI). Estos inhibidores forman un grupo de proteínas de masa molecular relativamente pequeña, 15 a 57 kDa, cuya unión a los com- plejos Cdk/ciclina determina la inactivación de éstos. En las células de mamíferos pueden distinguirse dos grupos de CDI: la familia Cip/Kip y la familia INK4. Los primeros son inhibidores de cualquiera de las Cdk, mientras los segundos son específícos para las Cdk que actúan con las ciclonas D. Los distintos miembros de cada familia actúan en tejidos específicos y proporcionan un mecanismo mediante el cual puede retardarse la progre- sión del ciclo celular en respuesta a señales tanto intracelulares como extracelulares. Análisis de ligamiento genético de familias con melanoma hereditario localizaron un gen putativo de susceptibilidad en la región cromosómica 9p21, la cual es un sitio de rearreglos cromosómicos frecuentes y pérdida de heterocigocidad en otros tumores es- porádicos. Esta región incluye los genes INK4a y INK4b ligados en tándem y las muta- ciones encontradas argumentan a favor de que el gen implicado es el INK4a. Reciente- mente se ha demostrado que mutaciones y deleciones que involucran a INK4a son fre- cuentes en diferentes tipos de tumores humanos. Esto significa que INK4a puede funcio- nar como un gen supresor tumoral.
Fosfoproteínas fosfatasas
Como ya fue señalado la actividad de los complejos Cdk/ciclinas depende del grado de fosforilación de la subunidad catalítica. Por lo tanto es el balance entre la actividad de las kinasas y las fosfatasas lo que en última instancia determina el progreso del ciclo celular.
desoxinucleótido y la posición 5´ del otro por lo que se denomina 3´ 5´. De esta forma la hebra posee un extremo con el grupo fosfato de la posición 5´ libre (extremo 5´) y el otro que presenta libre el grupo OH de la posición 3´ (extremo 3´). Cada desoxinucleótido a su vez está formado por una base nitrogenada que puede ser purínica o pirimidínica, por la D- 2 - desoxiribosa y una o más moléculas de ácido fosfórico. Las bases purínicas del ADN son la Adenina (A) y la Guanina (G), mientras que la pirimidínicas son la Citosina (C) y la Timina (T). Cuando se forma el polímero hay una zona con una estructura monótona pues en ella se alternan la desoxiribosa y el grupo fosfato a todo lo larga de la cadena, pero también una zona diversa pues las bases nitrogenadas que sobresalen de la estructura monótona son diferentes en cada sector de la molécula. Es precisamente en el orden o sucesión de esas bases nitrogenadas donde está contenida la información genética (Figura 2.3). 3´ (^) 5´
A T
C G
T A
G C
5´ (^) 3´ Figura 2.3. Diagrama de la estructura secundaria del ADN que muestra el eje covalente principal de desoxirribosa y fosfato, que forman la doble hélice con giro derecho. Las bases nitrogenadas (no representadas) se orientan hacia el interior de la molécula perpendiculares al eje covalente principal. Los pares de bases son obligados adenina con timina y guanina con citosina.
Watson y Crick propusieron que la molécula de ADN estaba formada por dos hebras que se disponían en forma antiparalela, es decir, el extremo 5´ de una coincidía con el 3´ de la otra y adquirían la forma de una doble hélice de giro derecho. La zona monótona estaba dispuesta hacia el exterior mientras que la zona diversa se orientaba hacia el interior de la molécula, de manera que las bases nitrogenada de una hebra se enfrentaban a las bases de la otra. Lo más trascendental del modelo era que la estructura solamente podía acomodar dos pares de bases, los formados por la Adenina y la Timina (A-T) y por la citosina y la Guanina (C-G). Las bases de cada par se dice que son complementarias. Estos pares se mantenían unidos por la formación de puentes de hidrógeno entre las bases, dos puentes en el par A-T y tres en el C-G. No existía restricción alguna para la sucesión de la bases en una de las hebras, pero la de la otra hebra venía determinada debido al carácter complementario del apareamiento. Este modelo permitió ver rápidamente el fundamento de una de las funciones más tras- cendentales de los seres vivos, esto es, su reproducción en seres de su misma especie. Para ello, las moléculas portadoras de la información genética debían duplicarse dando cada una dos moléculas idénticas a las progenitoras. Watson y Crick propusieron que durante ese proceso las dos hebras del ADN se separaban y cada una de ella servía de molde para la formación de la hebra complementaria. Esta idea básica ha resultado ser cierta, aunque el mecanismo es mucho más complejo que lo imaginado en los primeros momentos.
LA TRANSMISION DE LA INFORMACIÓN GENÉTICA
Tal vez la característica más sobresaliente de los seres vivos es la de poder originar organismos esencialmente iguales a sus progenitores, es decir, reproducirse. Esto sucede gracias a que los organismos contienen en su ADN toda la información necesaria para la formación de un organismo similar. Por lo tanto, para reproducirse al organismo dado le basta con obtener una copia lo más exacta posible de su ADN y transferírsela a sus descendientes. Desde el punto de vista molecular este proceso consiste en obtener dos moléculas de ADN idénticas entre sí e idénticas a la que les dio origen. En eso precisa- mente consiste la replicación del ADN que es el mecanismo molecular que sustenta la reproducción de todos los organismos vivos.
La replicación del ADN
Durante la fase S del ciclo celular se produce la replicación del ADN, proceso que consiste en obtener, a partir del ADN celular, dos moléculas idénticas de éste. Para este proceso se requiere el concurso de un gran número de proteínas enzimáticas y no enzimáticas.