¡Descarga celular nucleolo y más Apuntes en PDF de Biología Celular solo en Docsity!
3. Cromatina .-
3.1. Concepto.- Este material debe su denominación a su avidez tintorial por los colorantes básicos en microscopía óptica. Se trata de un complejo de nucleoproteinas que representa el genoma de las células eucarióticas y, por tanto, toda la información genética que esta posee (no se considera el genoma de los plastos y de las mitocondrias al hacer esta afirmación). Es el componente más abundante del Núcleo. Está constituida por DNA unido principalmente a proteínas del tipo de HISTONAS. La cromatina tanto del núcleo interfásico como en los cromosomas se tiñe con colorantes básicos y de acuerdo con la naturaleza del DNA, absorbe la luz ultravioleta a 260nm. También muestra una reacción positiva a los siguientes tests histoquímicas:
- Test de Feulgen.- Esta técnica produce: a. Hidrólisis de RNA mediante una solución ácida débil de ClH. b. Sobre el DNA produce la eliminación de las bases púricas quedando libres los radicales aldehídos de las pentosas. c. Tratamiento con leuco-fuchina (colorante que en estado reducido es incolora y cuando es oxidada por los grupos aldehídos que han quedado libres se torna coloreada. - Tinción con verde metilo-pironina.- El verde metilo pironina tiñe al DNA, mientras que la pironina tiñe al RNA. Con esta técnica la cromatina y los cromosomas quedan teñidos en verde. - Tratamiento enzimático con nucleasa.- La DNAasa disuelve la cromatina permaneciendo intacto el RNA y al contrario.
3.2. Composición .- La cromatina aislada presenta tres componentes principales: ADN, ARN y proteinas.
3.2.1. El ADN.- - Es una molécula de doble hélice helicoidal con 2,5 nm de Ø.
- Cada cadena es la unión de nucleótidos. Cada nucleótido está formado por: bases, pentosa y fosfato.
- Las bases son: Púricas (A y G) y Pirimidinicas (T y C).
- La pentosa es la desoxirribosa.
- Las dos cadenas son antiparalelas.
- Es la sede de los genes.
- Cada región de ADN que produce una molécula de ARN funcional es un gen. En eucariotas es bastante común que un gen pueda tener más de 100.000 PB. Puesto que solo son necesarios unos 1.000 nucleótidos para codificar una proteina de tamaño medio, es obvio que existen dentro del gen fragmentos que no codifican. Las secuencias que codifican se llaman exones y las secuencias que no codifican se denominan intrones. El primer ARN (transcrito primario) generado a partir de un gen ha de sufrir un procesamiento (splicing) en el que le son retinados los intrones. Además cada gen posee
una serie de secuencias regulatorias que sirven para la transcripción adecuada del gen. Cada gen puede tener unas I00.000 Pb aunque hay genes con 2.106.
Fig. 1.
Fig. 3.-
Fig. 4. Cada uno de estos tipos tiene 102 a 135 aa y su estructura se ha mantenido muy constante durante la evolución, sobre todo la de las histonas H3 y H4. Las historias se diversifican aún más mediante algunas modificaciones de los aminoácidos, como fosforilación o acetilación, que ocurren después de la síntesis de los polipépti- dos. Las secuencias de aminoácidos de las histonas, particularmente H3 y H4, han sufrido muy, pocos cambios a través de periodos muy prolongados de la evolución. Por ejemplo, la histona H4 de los guisantes y de las vacas contiene 102 aminoácidos y su secuencia sólo difiere en dos residuos animo, ¿por qué están tan conservadas las histonas?.
Una razón es que las histonas interactuan con el esqueleto de la molécula de DNA, que es idéntico en todos los organismos. Además casi todos los aminoácidos de una molécula de histona Participan en una interacción con otra molécula, ya sea DNA, ya sea otra histona. Como resultado, muy pocos aminoácidos de una histona pueden ser reemplazados por otros aminoácidos sin que se vea afectada de manera
proteínas no histónicas represtan al conjunto de enzimas involucrados en la replicación, transcripción y regulación de la transcripción del ADN. Además, destacan otras proteínas, como una proteína contráctil del tipo de la actina con un peso molecular de 45.000 Da, tubulinas α y β (propias de los microtúbulos) y una miosina de elevado peso molecular, 225.000 Da. La presencia de esta; proteínas contráctiles se interpreta como clave en los procesos de condensación de la cromatina en cromosomas y posteriormente en los movimientos de los mismos en las fases de la división nuclear y celular. El ADN está unido a las histonas en relación 1:1 y con las no-histónicas en relación 1:0,6. La relación entre ADN/ARN es de 1:0,1.
Existen una pequeña proporción de otras proteínas, principalmente – ACIDAS del tipo de las fosfoproteinas que son muy abundantes en la eucromatina. Estas proteínas tienen una alta velocidad de recambio (alto turnover) y porque se sintetizan durante todo el ciclo celular a diferencia de las histonas que solo se sintetizan durante la fase S. Las proteínas ácidas del núcleo mejor conocidas son: la nucleoplasmina que se une a la s histonas H2A y H2B y la proteina N1 que se une a las H3 y H4. En el núcleo hay también proteínas ácidas que no se unen al DNA ni al RNA y que se denominan proteínas ácidas residuales. Otras proteínas que aparecen unidas al DNA son diversas enzimas, acetilasas de las histonas, etc.
A lo largo del ciclo celular, la cromatina sufre una serie de cambios bastante importantes, aun cuando desde el punto de vista bioquímico se puede decir que no hay alteraciones. Existe un cambio cuantitativo pues la cantidad de cromatina se duplica si consideramos G1 y G2. Además, también existe un cambio de aspecto pues toda la cromatina interfásica se transforma en cromosomas mitóticos. Este cambio de aspecto es el reflejo de un mayor grado de condensación, es decir, de un mayor grado de estructuración. A continuación se analizará la estructura de la cromatina interfásica y la estructura del cromosoma mitótico.
3.3. Estructura. -
3.3.1. La cromatina interfásica .- La cromatina que forman los cromosomas interfásicos se haya estructurada en una serie de niveles de organización que determinan su mayor condensación. El nivel de condensación de la cromatina guarda una estrecha relación con la actividad transcripcional del ADN que contiene. El empaquetamiento del ADN se realiza a varios niveles, piensese que el ADN humano mide aproximadamente 2m y que ha de empaquetarse en el núcleo que tiene un volumen medio de 25 μ3.
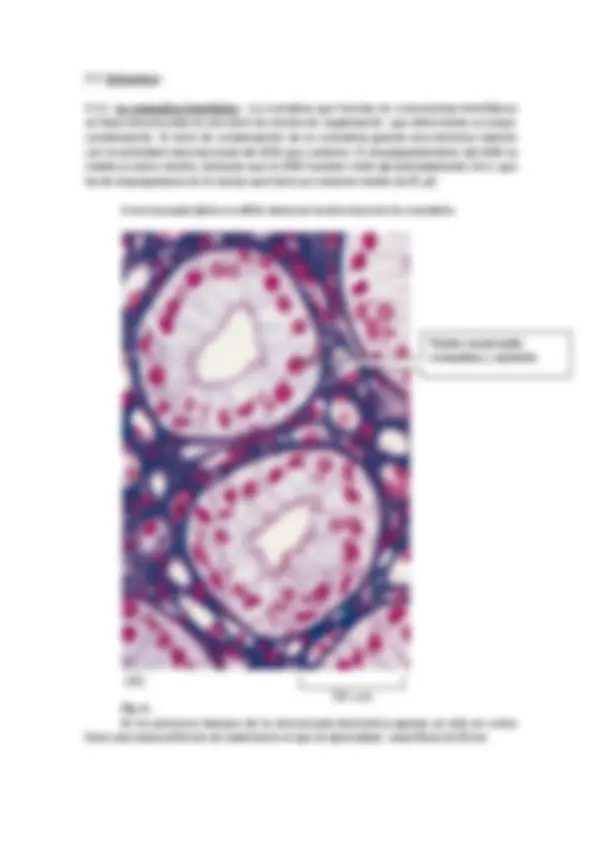
A microscopía óptica es difícil observar la estructura de la cromatina.
Fig. 6.- En los primeros tiempos de la microscopía electrónica apenas se veía en cortes finos una masa uniforme de material en el que se apreciaban unas fibras de 30nm.
Núcleo mostrando cromatina y nucleolo
FFf Fig. 8. F
Cuando se estudia la cromatina o los cromosomas “in toto” y no en cortes mediante la técnica de explotar los núcleos y dispersar la cromatina en una interfase aire-agua, en donde las fuerzas de tensión superficial separan las fibras. Se pudo deducir de estos estudios que: “La cromatina está constituida por una larga hélice de DNA que al asociarse a proteínas – principalmente histonas- formaría una fibra de 10nm de, pero a su vez sufriría una serie de plegam ientos dando lugar a una fibra de 30 nm. Finalmente plegamientos más complejos darían lugar a fibras más gruesas de Ø variable”.
La gran dificultad residía en explicar, ¿cómo se dispone el doble hélice de DNA de 2,5nm y su relación con las proteínas para dar primero la fibra de 10nm y posteriormente la de 30nm? Una técnica permitió, en parte, dar respuesta a estas cuestiones. Cuando los núcleos como los de oocitos de anfibios, son explotados y rota su cubierta nuclear (la envoltura nuclear se rompe por choque osmótico, los núcleos aislados se colocan en un medio hipotónico que los hace hincharse y estallar o por sonificación, mediante ultrasonidos, se rompen las membranas de la envoltura). La cromatina se dispersa en una interfase agua-aire, posteriormente esta cromatina recogida en una rejilla y teñida con tinción negativa o bien sombreado con metales pesados, se aprecia que esta cromatina presenta una secuencia de glóbulos de 30nm de Ø.
Si la preparación de la cromatina se hace en un tampón de baja fuerza iónica, en ausencia de iones divalentes, (se extrae la H1) aparecen glóbulos separados unos 14nm entre sí y unidos por filamentos de DNA de unos 14nm de longitud. “La imagen es como la de un collar de perlas. A los glóbulos se les denominó NUCLEOSOMAS y a la hilera de cuentas cadena nucleosómica o NUCLEOFILAMENTO.
Fig. 9. When the contents of amphibian erythrocyte nuclei were prepared for electron microscopy by spreading on the surface of water, examination of whole mounts of the dissociated material revealed a tangle of 20 to 30 nm fibers (upper micrograph). After pretreatment with chelating agents, the chromatin in such preparations appeared as knobby 10 nm filaments (Ris, 1968). If the isolated chromatin was pretreated with urea, 2 to 4 nm filaments were observed.
Nucleofilaments from chicken erythrocyte nuclei lysed on low ionic buffer in the absence of divalent metals. (Micrograph courtesy of Ada Olins.)
región central (la unión de los 4 extremos compactos de las 4 histonas) y 4 colas (los 4 extremos distendidos) de unión al DNA. De la unión de dos tetrámeros resulta el nucleosoma completo.
Fig. 12.-
La compresión del empaquetamiento de DNA avanzó de manera notable en años recientes gracias a la importantes datos de la partícula del núcleo del nucleosoma obtenidos por cristalografía de Rayos X. Las 8 moléculas de histona se organizan en cuatro hetrodímeros: 2 dímeros H2A-H2B y 2 dímeros H3-H4. La dimerización de las moléculas de histona es mediada por sus dominios C-terminal, que constan sobre todo de hélices α plegadas en una masa compacta en el núcleo del nucleosoma. En contraste, el segmento amino terminal (N-terminal) de cada núcleo histónico (y también el segmento C-terminal de H2A) toman la forma de una cola flexible y larga que se extiende más allá de la hélice de DNA y hacia los alrededores. Estas colas son blancos de una variedad de modificaciones covalentes cuyas funciones clave se explicarán más adelante.
El DNA y los núcleos de histonas se mantienen juntos mediante distintos tipos de enlaces no covalentes, inclusive los enlaces ionicos entre los fosfatos con carga negativa del esqueleto de DNA y los residuos con carga positiva de las histonas.
Fig.13.
Las dos moléculas hacen contacto en sitios donde el surco menor del DNA se relaciona con el núcleo de la histona, lo que ocurre a intervalos aproximados de 10 pares de bases (los ganchos blancos en la figura 12-11C). Entre estos puntos de contacto las dos moléculas están separadas por un espacio considerable, que debe brindar acceso a los factores de transcripción del DNA y otras proteinas que unen al DNA. Aunque por muchos años se pensó que las histonas eran moléculas estructurales inertes, como se verá en las siguientes secciones, estas pequeñas proteinas, tienen funciones de importancia crítica en la determinación de la actividad del DNA con el que se relacionan. Los residuos básicos de aminoácidos del núcleo central de las histonas se agrupan sobre uno o ambos extremos de la molécula, y por lo tanto el resto de la molécula permanece con carácter relativamente hidrofóbico. Esta separación es ideal para la organización del nucleosoma. Las partes más hidrofóbicas, partes no cargadas de las histonas, ocupan el centro de la partícula del nucleosoma y por lo tanto promueven su agregación en un núcleo apretado. Por lo contrario, las partes básicas polares del núcleo de las histonas forman apéndices flexibles dirigidos hacia el exterior de la partícula, donde los residuos con carga positiva se encuentran disponibles para generar interacciones iónicas con grupos fosfato cargados negativamente del esqueleto de DNA. Aun cuando el DNA se encuentra íntimamente asociado con el núcleo de la histona sobre la superficie interna de la fibra helicoidal, la superficie externa del DNA todavía está expuesta y accesible para interactuar con moléculas reguladoras.
Los octámeros forman una unidad fija, es decir, no se desplazan por la cadena. Esta fibra de 10nm corresponde a la cromatina totalmente desespirilizada, tal como se encuentra en el momento de actividad funcional (replicación o trascripción. Pero
modelo en “zigzag” mostrado en la figura 15- b en la que los nucleosomas sucesivos a lo largo del DNA se disponen en distintas pilas y los nucleosomas alternados se convierten en vecinos interactivos.
Fig.
La figura 15- b,c muestra dos modelos en los que el filamento nucleosómico se enrolla en un orden superior, una fibra más gruesa. Los modelos difieren en la posición relativa de los nucleosomas dentro de la fibra. La investigación reciente favorece el modelo en "zigzag" mostrado en la figura 12-10b, en el que los nucleosomas sucesivos a lo largo del DNA se disponen en distintas pilas y los nucleosomas alternados se convierten en vecinos interactivos.
El modelo del solenoide explica el paso de la fibra de 10nm a la fibra de 30 nm por un enrollamiento en hélice levógira en el cada vuelta o cuparia 6- nucleosomas (fig 16)
Fig. 16.-
Sin considerar en que lo anterior se realiza , el ensamblaje de la fibra de 30nm incrementa el índice de empaquetamiento unas 6 veces más, o cerca de 40 veces el total
Fig .- 17
Se estima que cada asa contiene entre 10 y 150 kilobases (kb) de DNA y que está unida en su base a proteínas específicas, incluyendo una topoisomerasa tipo II que presumiblemente regula el grado de superenrollamiento del asa de DNA. Sería de esperar que la topoisomerasa desenrede las moléculas que el DNA de un asa hubiera enredado. Las asas de DNA pueden actuar para dividir el genoma en "dominios" que contengan un solo gen o un pequeño conjunto de genes enlazados expresados por un mecanismo regulador común. Con asas de tamaño promedio de 50 kb, un conjunto haploide de cromosomas humanos contiene casi 60 000 asas; este es el mismo orden de magnitud del número de genes que se cree contiene el genoma humano.
El contenido de un núcleo interfásico tratado con soluciones de baja fuerza iónica que extraen totalmente todas las histonas, libera un esqueleto proteico alrededor del cual irradian núcleofilamentos. Muchos de estos nucleofilamentos están dispuestos en bucles de alrededor de 50KB, cuyos dos extremos están insertados en el entramado proteico. A nivel de sus puntos de inserción, los bucles son ricos en secuencias repetidas
que tienen las propiedades de ADN satélites. En la parte distal de los bucles se observan complejos de transcripción. La cromatina difusa proviene del desplegamiento del nucleofilamento en bucles a nivel de los cuales se efectua la transcripción. Las diferentes regiones cromosómicas axiales estarían asociadas a un esqueleto nuclear conectado con la lámina protéica que está adosada contra la membrana interna de la envoltura nuclear.
Fig. 18.-
(a) (b)