¡Descarga citoplasma - celular y más Apuntes en PDF de Histología solo en Docsity!
Apunte – CELULA La célula se divide en dos comportamientos principales, citoplasma y el núcleo. El citoplasma es la región que se separa del núcleo. Este contiene organelas u orgánulos, Citoesqueleto que son proteínas polimerizadas en Microtúbulos, fibras intermedias y filamentos de actina; y también encontraremos inclusiones suspendidas en la matriz mitocondria. La matriz está formada por varios solutos, como Na+, K-^ y Ca+2^ y moléculas orgánicas como glucosa, proteínas, lípidos y ARN. Tanto núcleo y citoplasma si bien desempeñan tareas diferentes actúan de forma conjunta para controlar el metabolismo, concentraciones de sustancias y llevar adelante la viabilidad celular. De los componentes del citoplasma podremos diferenciar los membranosos donde mencionamos, la membrana plasmática, complejo de Golgi, RER, REL, endosomas, lisosomas, vesículas, mitocondrias, peroxisomas que separan el contenido interno de estas con el externo; los no membranosos mencionamos microtúbulos, filamentos intermedios, filamentos de actina, centriolos, ribosomas y proteosomas, estas estructuras conforman el Citoesqueleto. También podremos encontrar inclusiones que en general no poseen membrana plasmática, estas pueden ser cristales, lípidos, proteínas, glucógeno etc. Orgánulos membranosos Membrana plasmática: la membrana también mencionada como plasmalema, es una estructura que se describe como bicapa fosfolipidica que solo es visible por microscopia electrónica de transmisión. El espesor de esta es de aprox. 8 a 10nm. Las porciones de esta membrana las podemos describir químicamente como hidrofilicas que tiene afinidad por el agua por lo que su posición quedara del lado externo y la porción hidrofóbica queda hacia adentro aislada. La membrana plasmática cumple con el modelo del mosaico modificado, que significa esto? significa que la predisposición de la bicapa lipídica y su tridimensionalidad permite que las estructuras que están insertas en ella no están rígidas o estáticas, sino que presenten cierta fluidez para poder predisponerse en las funciones específicas que posean. La membrana principalmente está compuesta por moléculas de fosfolípidos, y en la zona hidrófoba de esta encontraremos de forma aleatoria colesterol que cumplirá funciones de estructura y le dará la característica de fluida. También aclaremos que este colesterol en altas concentraciones ara que la membrana se vuelva rígida, por lo cual esta debe estar en una concentración indicada, también mencionaremos que este colesterol evitara el congelamiento de esta y conservando la fluidez. A modo de resumen el colesterol tiene un papel vital en la vida de la célula. Si clasificamos a las proteínas que podremos observar en la membrana son proteínas integrales que están incluidas en la bicapa lipídica atravesándola, y superficialmente podremos observar proteínas periféricas que se asocian a la membrana mediante interacciones iónicas fuertes. A su vez hay hidratos de carbono que pueden unirse a estas proteínas formando glicoproteínas o también uniéndose a lípidos denominándose glicolípidos. Estas moléculas perfiladas hacia afuera de la bicapa lipídica se denominara glucocaliz o cubierta celular, que sirven como método de comunicación entre el medio intracelular y el extracelular, también cumplirá con funciones de reconocimiento celular y reconocimiento de hormonas.
Balsas lipídicas: Son micro dominios de la membrana plasmática comúnmente formados por colesterol, glucoesfingolipidos y presencia de ácidos grasos saturados. Son consideradas plataformas de señalización que están equipadas con receptores, factores de acoplamiento, enzimas y sustratos. En infecciones bacterianas y víricas la interacción se produce en estas balsas. [ En algunos casos algunas bacterias usan este mecanismo para ingresar al interior de la célula sin ser detectado como un agente peligroso y se libran de la fagocitosis.] Existen distintos tipo de balsas lipídicas:
- Balsas lipídicas planas: además de colesterol y lípidos estas contienen proteínas denominadas fotilinas que tienen función de marcadores moleculares y son consideradas proteínas de andamiaje.
- Balsas caveolares: tienen forma de botella, tienen proteínas integrales de membrana llamadas caveolinas que tienen la capacidad de unirse al colesterol y a otras señales que participan en la transducción de señales. Proteínas estructurales: Podemos definir 6 tipos de proteínas de membranas, aunque ninguna de las 6 son excluyentes entre si ya que una puede tomar la propiedad de enzima, bomba o receptor etc. Bombas: transportan activamente ciertos iones como Na+^ y K+^ a través de la membrana. Pueden transportan precursores metabólicos, macromoléculas solas, o macromoléculas ligadas a un catión sódico. Canales : permiten el paso de iones y solutos pequeños en cualquier dirección por difusión pasiva. Estos también van a estar formando las uniones de hendidura, que conectan los citoplasmas de una célula con otra que esta contigua. Proteínas receptoras: permiten la único y reconocimiento de un ligando, como hormonas, reacción de endocitosis o reacción de anticuerpos. Estas proteínas pueden comunicarse a segundos mensajeros y producir respuestas fisiológicas. Proteínas ligadoras: estas fijan el Citoesqueleto a la matriz extracelular. Podemos mencionar a las integrinas que vinculan a los filamentos de actina del Citoesqueleto con las moléculas de fibronectina de la matriz. Enzimas: poseen muchas funciones, pero si nombramos específicamente a las ATPasas tiene funciones específicas de bombeo de iones en mitocondrias y enzimas digestivas. Se las considera proteínas integrales de membrana. Proteínas estructurales: forman uniones de células adyacentes. Algunos ejemplos pueden verse en células polares como las células epiteliales.
ligandos como en las células musculares, o canales iónicos activados por fuerzas mecánicas como los del oído interno. Transporte vesicular: este tipo de transporte está relacionado directamente con el ingreso o egreso de sustancias grandes de la célula. El mecanismo que se implementa es la brotacion o gemación vesicular, que consta en la formación de una vesícula que recubra y contenga el contenido a desplazar. En términos más puntuales, cuando algo grande necesita ingresar a la célula lo hace mediante la endocitosis y si algo grande necesita salir lo hace mediante exocitosis. Tanto la endocitosis como la exocitosis trabajan paralelamente y dependen una de otra. Si una se detiene la otra también. Y esto se debe a que las proteínas implementadas en que se genere la exocitosis ( proteínas SNARE ) son las que median el inicio de la endocitosis. [ Esto se demostró en un estudio en neuronas afectadas por la neurotoxina botulínica que bloqueaban la exocitosis también bloqueaban la endocitosis.] Endocitosis: es un mecanismo que media el ingreso de sustancias a la célula, como lípidos, proteínas, nutrientes o moléculas de señalización. Estos una vez dentro de la célula son denominados endosomas que serán degradados en el futuro. El método más conocido y más común de ingreso es mediante proteínas de membrana denominadas clatrinas que median el ingreso celular, aunque actualmente se sabe que no son fundamentales ya que también pueden utilizarse caveolinas o fototilinas para ello. Por lo que podemos decir que la endocitosis se clasifica si es dependiente de clatrinas o independiente de estas. En general la endocitosis tiene 3 mecanismos de acción: pinocitosis, fagocitosis y endocitosis mediada por receptores. Micropinocitosis: es una ingesta inespecífica de sustancias liquidas y proteínas por medio de vesículas pequeñas. La presencia de estas suele está relacionada con las proteínas caveolinas y fototilinas de las balsas lipídicas. En la observación mediante el MET se las describe lisas y se la encuentra mayormente en el endotelio vascular y células musculares lisas. La Micropinocitosis tampoco requiere de modificación del Citoesqueleto de actina, por lo cual también se la denomina como endocitosis independiente de clatrinas o de actinas. Macropinocitosis: este mecanismo media el ingreso de solutos proteínas y anticuerpos y se lo considera de captación inespecífica pero este si requiere de las actinas del Citoesqueleto debido a que deberá realizar pliegues en la membrana de la superficie. Estas generan grandes vacuolas endociticas llamadas macropinosomas. Este mecanismo es utilizado mayormente por células del sistema inmune como los macrófagos y células dendríticas. Este mecanismo responde mucho a factores de crecimiento como por ejemplo: factor de crecimiento 1 de macrófagos, factor de crecimiento dérmico, y factor de crecimiento derivado de plaquetas. La Micropinocitosis también se la conoce como endocitosis independiente de clatrinas pero dependiente de actinas. (Debido a la remodelación de membrana que se produce por el ingreso de sustancias grandes). Fagocitosis: la fagocitosis es la ingesta de partículas grandes como residuos celulares. Se considera un sistema específico donde la célula crea seudópodos para engullir a las sustancias extrañar que se quieren degradar haciéndolas ingresar como un fagosoma. Generalmente cuando estas sustancias ingresar mediante la señal de un anticuerpo estos se unen en la región del anticuerpo que no recepta a la partícula extraña (en su región Fc). La fagocitosis también se desencadena mediante patrones
moleculares asociados con patógenos. No todas las partículas van a necesitar de este anticuerpo o clatrinas para su ingreso debido a la creación de los seudópodos por parte de la célula, las sustancias que no utilizan estas clatrinas suelen ser partículas de polvo, sustancias carbonadas asbesto inhalado. Para la formación del fagosoma el Citoesqueleto debe modificarse por lo que la actina se despolimeriza y vuelve a polimerizarse, por lo tanto la fagocitosis es considerada como independiente de clatrinas pero dependiente de actinas. Endocitosis mediada por receptores: se genera por receptores de carga que se acumulan en las regiones donde se encuentran las balsas lipídicas que luego se convertirán en fositas recubiertas. En estas fositas se acumulan las clatrinas donde producirán una depresión en la membrana formando una especie de jaula para recibir a la sustancia a ingresar. Las clatrinas interactúan con las adaptinas produciendo el ingreso de la vesícula. Después una enzima llamada dinamina media la liberación de las clatrinas que recubren a la vesícula dejándola sin cubierta lista para fusionarse con un lisosoma. Exocitosis: proceso en el cual las sustancias salen de la célula. Se realiza a través de dos mecanismos: Constitutivo: las proteínas que salen de la célula son excretadas de forma automática después de sus síntesis. Es un mecanismo de todas las células. De secreción regulada: Es cuando las células acumulan sus secreciones en el interior en vesículas y las liberan en un momento dado, mediado por un estímulo externo. Inclusiones: Son considerados componentes no vivos de la célula, que puede o no estar rodeado por membrana. Las inclusiones contienen productos de la actividad metabólica de la célula y consisten principalmente en orgánulos de pigmento, gotitas lipídicas y glucógeno.
- Lipofucinas: Pigmentos dorado pardo que se tiñe con H&E. Se observa de forma fácil en célula que no se dividen como en neuronas, células musculares cardiacas y esqueléticas. Se las denomina como pigmento del desgaste debido a que se acumulan acorde pasa el tiempo y envejece la célula. Las células fagociticas como los macrófagos suelen contener Lipofucinas que se acumulan por la digestión célula de bacterias, partículas extrañas y detritos celulares.
- hemosiderina: es un complejo de almacenamiento de hierro. Se suele formar de residuos no digeribles de la hemoglobina y se relaciona con la presencia de fagocitosis de eritrocitos.
- glucógeno: es un polímero ramificado que usa para almacenar energía. No se tiñe con técnicas de rutina domo H&E suele encontrarse en altas concentraciones en musculo estriado esquelético.
- Inclusiones lipídicas: o también llamadas gotitas de grasa son nutrientes que proporcionan energía al metabolismo celular.
- Inclusiones cristalinas: estas inclusiones se suelen encontrar en las células de sertoli y células de Leydig del testículo. (sin importancia biológica conocida)
proceso inespecífico que produce la degradación de proteínas citoplasmáticas de forma lenta y fisiológica. Y por último la autofagia mediada por chaperonas que básicamente estás median la especificidad de lo que se va a degradar cuando hay carencia de nutrientes, generando señales de reconocimiento para estas. Degradación mediada por proteosomas: Los proteosomas son complejos proteínicos que actúan sin la participación de lisosomas. La degradación mediada por estos se rige en la utilización de proteasas dependientes de ATP que destruyen proteínas que han sido marcadas específicamente para ser destruidas. Aunque también actúan en proteínas de corta duración como las proteínas que participan en la división celular. Retículo endoplasmático rugoso: Es el también llamado ergastoplasma por cómo se lo observa en el MET. Es una serie de sacos aplanados denominadas cisternas que se encuentran contiguos a membrana nuclear. Su característica de rugoso es debido a la presencia de ribosomas que están adheridos, estos están compuestos por una subunidad menor y una mayor. Cada subunidad está compuesta de ARNribosomal, de distintas longitudes, así como varios tipos de proteínas. Los grupos de ribosomas forman arreglos espirales cortos denominados polirribosomas o polisomas que están adosados a un mismo ARNmensajero. S íntesis proteica: la síntesis proteica comienza en el núcleo en la traducción del gen correspondiente. Posteriormente a la traducción de ADN hacia el pre-ARN. Se generan modificaciones postraduccional (que incluye el corte, empalme adición de las Poli-A en el extremo 3’ y un capuchón de metilguanosina en el extremo 5’, que le den estabilidad a la molécula para que no sea degradada en el citoplasma), luego viene la traducción donde el ribosoma lee el mensaje codificado para crear un Polipeptido. Luego los ribosomas libres en el citoplasma tienen las mismas características que los ribosomas y polirribosomas pero no están adheridos a una membrana plasmática. Estos traducen ARNm para sintetizar proteínas para la propia célula, donde previo a ser sintetizadas son marcadas o etiquetadas para que estas se dirijan a su destino correcto. Ahora si la proteína es de exportación celular, su síntesis es en el RER, donde se producirán modificaciones postraduccionales como glucosilacion central , formación de enlaces hidrogenados disulfuro y plegamiento de la molécula minutos después esta viajara al Golgi. El RER está muy desarrollado en células secretorariamente activa. El RER y el Golgi poseen un trasporte de vesículas del tipo anterógrado y retrogrado que estas están mediadas por proteínas del tipo COP-I (para el trasporte retrogrado) y la COP-II (para el transporte anterógrado) y estos procesos están implicado en el reciclado de estas vesículas [ los corpúsculos basófilos de las neuronas denominados corpúsculos de nissl están compuestos por el RER y una gran cantidad de ribosomas libres] Retículo endoplasmático Liso: son túbulos anastomosados que no están asociados con los ribosomas, presentan gran eosinofilia al microscopio óptico, son abundantes en células que sintetizan lípidos (ácidos grasos y fosfolípidos). Este orgánulo se encuentra muy desarrollado en células que sintetizan esteroides, como en la corteza suprarrenal y las células intersticiales del testículo (Leydig), en células musculares cardiacas y esqueléticas se lo denomina retículo sarcopasmatico este retículo secuestra el calcio que es esencial para la contracción muscular. El REL participa en la desintoxicación por lo que está muy desarrollado en las células hepáticas relacionado con enzimas con el citocromo P450. El REL participa también en, el metabolismo de lípidos y esteroide, metabolismo de glucógeno y la formación y el reciclado de la
membrana. [Según el papel funcional también se la relaciona con otras enzimas como las hidrolasas, metilasas, glucosa-6-fosfatasa, ATPasa, y oxidasas de lípidos.] Aparato de Golgi : Esta organela está muy desarrollada en células del tipo secretora y no se tiñen con H&E. En la célula se presenta como una red de sacos aplanados y cisternas de membrana denominada rimeros. El aparato de Golgi esta polarizado con una región cis de Golgi de cisternas aplanadas, y una región trans. Las redes que se encuentran en la red cis y la red trans se denomina red intermedia del Golgi. El aparato de Golgi participa en modificaciones postraduccionales en la clasificación y empaquetado de proteínas. Esas proteínas son glucosiladas, fosforiladas y sulfatadas. el Golgi ocupa cuatro vías de exportación celular según el destino. Membrana plasmática apical: las vesículas que salen de la TGN a la membrana apical es sin clatrinas. Membrana plasmática basolateral: Las proteínas enviadas al dominio basolateral también tiene una señal de clasificación específica. Esta vía utiliza laguna proteína específica que luego se asociara con otra adaptadora del epitelio. Endosomas y lisosomas: La mayoría de las proteínas de las proteínas que salen del Golgi están etiquetadas específicamente para convertirse en endosomas y lisosomas. Pero cuando no ocurre esto, las proteínas deben utilizan marcadores M-6-P destinados a los endosomas tardíos y posteriormente se transforman en lisosomas maduros. Citoplasma Apical: es un sistema muy característico de células secretoras. Las vesículas liberadas de la zona trans de Golgi sufren un proceso madurativo consecuencia del cambio de PH y de las concentraciones de calcio. Luego se fusionan a la membrana y son expulsadas por exocitosis. Mitocondria: organela relacionada directamente con la producción de energía celular, se encuentran en altas concentraciones en células como las de músculo estriado, tanto cardiaco como esquelético. Su origen se le atribuye a la teoría endosimbiotica, donde se explica que las mitocondrias comenzaron como organismos procariotas del tipo bacterias de vida libre, que ingresaron a células de mayor porte por endocitosis que no podían generar energías por si sola o que tenían una producción deficiente de la misma. Estos indicios se confirmaron debido a que las mitocondrias tenían sus propios ribosomas, sintetizan 13 de sus propias enzimas, y que su ADN es propio y circular distinto al que se encuentra en el núcleo. Podemos afirmar que las mitocondrias están presentes en casi todas las células excepto en eritrocitos y queratinocitos terminales. La mitocondria posee compartimentos y limites bien definidos que podemos clasificarlos en: membrana mitocondrial externa, membrana mitocondrial interna, espacio o matriz intramitocondrial y espacio intermembrana. Peroxisoma: son orgánulos limitados por una sola membrana que contienen en su interior eximas del tipo oxidativas. Estas en su interior producen peróxido de hidrogeno (H 2 O 2 ) y catalasas. Como el peróxido de hidrogeno es muy toxico en el organismo es escindido en agua y oxigeno (H 2 O + O 2 ) por la catalasa. Además también los peroxisomas contienen D- aminoácidos-oxidasas y enzimas de la B-oxidación entre otras. Los peroxisomas participan en la beta-oxidación que puede igualar a la de las mitocondrias. Si bien los peroxisomas son abundantes en células hepáticas y en células renales pueden están presentes también en gran medida en respuesta a la dieta, fármacos y a estimulación hormonal.
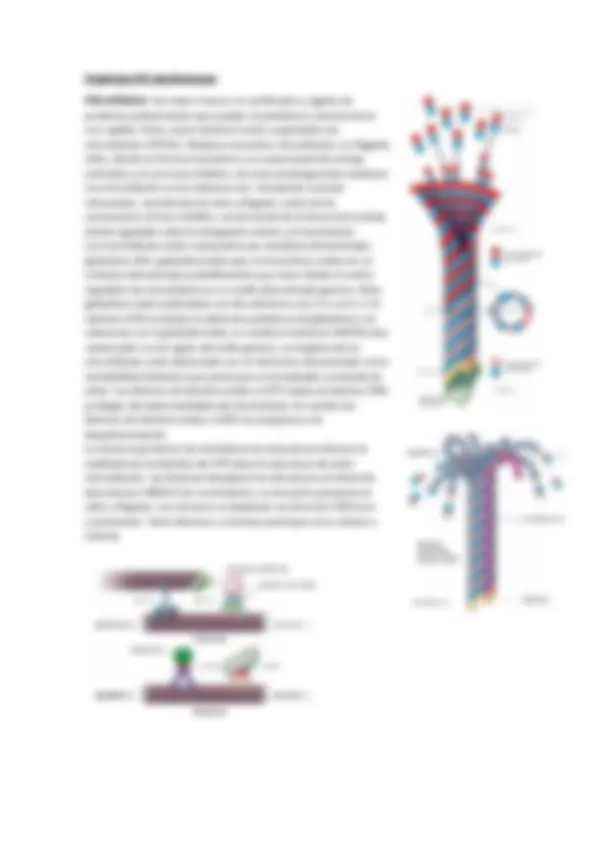
Filamentos de Actina: son abundantes y constituyen alrededor del 20% de las proteínas de algunas células no musculares. Estos filamentos también se ensamblan de forma espontánea, en predisposición helicoidal. Son más flexibles que los microtubulos. Las proteínas que conforman los filamentos de actina son actina globular G que cuando se encuentran unidas se conocen como actina filamentosa f (actina F). Su extremo de crecimiento rápido se llama extremo positivo o (espiculado), y su extremo de crecimiento negativo (puntiagudo). Los filamentos de actina presentan proteínas responsables del ensamblado y organización de estos. Podemos nombrar a proteínas formadoras de fascículos de actina (fascina y fimbrina) y proteínas cortadoras de filamentos que cortan las largas fibras en filamentos cortos. (Gelsolina), y las proteínas Cofilina está implicada en la remodelación rápida del Citoesqueleto de actina creando extremos libres positivos y negativos, las proteínas formadoras de casquetes que bloquean la adición de más moléculas de actina en el extremo libre. (Tropomodulina). Proteínas formadoras de enlaces cruzados en la actina.(presentes en eritrocitos como espectrinas, aductina, proteína 4.1 y 4.9) y por último, la proteína motoras de la actina. Los filamentos de actina están relacionaos con las siguientes funciones.: anclaje y movimiento de las membranas, formación del núcleo estructural de las microvellocidades, locomoción celular y emisión de evaginaciones celulares. Filamentos intermedios: Los filamentos intermedios en general tienen función de sostén, se los considera intermedios porque tienen medidas intermedias entre microtubulos y filamentos de actina. Este dominio se caracteriza por ser en forma de varilla con extremos globulares diversos. Estos filamentos intermedios se ensamblan partir de un par te monómeros helicoidales que se enroscan entre si para formar dímeros enrollados. Este enrosque es antiparalelo a partir de tetrámeros escalonados de dos dímeros. Podemos nombrar varios tipos de filamentos intermedios: clase 1 y 2, clase 3, clase 4, clase 5 y clase 6. Los filamentos intermedios se asocian fuertemente a uniones célula-célula y de célula-matriz extracelular. (desmosomas y hemidesmosomas). Orgánulos microtubulares Centro organizador de microtubulos o centrosoma: suelen estar formados un por material denso y amorfo, comúnmente rodeado por aparato del Golgi y contiguo al núcleo celular este espacio ce lo suele denominar “pericentriolar” y está contenido por centriolos en una posición ortogonal, y conformados por nueve tripletes de microtubulos con un centro vacío. (9+0). Aquí es donde se originan y organizan la mayoría de los microtubulos formados en la interface celular, controlando su cantidad polaridad y dirección. Dentro de las funciones primordiales del centrosoma es la formación de los cuerpos basales (donde estos se forman sin establecer contacto con los pres existentes y otros se forman a partir de otros existentes, ambas vías dan
origen a pro centriolos que posteriormente maduraran a cuerpos basales) y de la formación del uso mitótico. Cuerpos basales: Cuando los centriolos son recién formados en una célula, estos tienden a ir hasta la superficie apical de la célula para trabajar como centros organizadores para el ensamblado de los microtubulos de los cilios, la configuración de estos cilios es de 9 dobletes más un doblete central (9+2). Matriz citoplasmática: solución acuosa concentrada que contiene moléculas de diferentes tamaños y formas, como electrolitos, metabolitos, ARN, y proteínas sintetizadas. NUCLEO CELULAR Estructura y organización del núcleo interfásico: El núcleo es un compartimiento limitado por membrana el cual contiene el genoma (info genética) de las células eucariotas. Cuando este no se está dividiéndose, este periodo se conoce como Interfase, y está formado por los siguientes componentes: cromatina (que a su vez puede ser eucromatina o heterocromatina), nucléolo, envoltura nuclear y nucleoplasma. Cromatina: Está compuesta por complejos de nucleoproteínas. Un complejo de cromatinas con ADN y proteínas estructurales cuando se pliega produce las estructuras denominadas cromosomas. Además las cromatinas contienen proteínas del tipo histonas y no histonicas. Esta cromatina se puede clasificar en heterocromatina y eucromatina. La heterocromatina es la que se encuentra compactada y se considera inactiva por que no se está replicando. A su vez esta heterocromatina se subclasifica en constitutiva y facultativa; La constitutiva contiene secuencias repetitivas y genéticamente activas del ADN y la facultativa no participa en el proceso de trascripción. Por otro lado la eucromatina es la región clara del núcleo y es activa porque se produce la transcripción genética del ADN. Las unidades más pequeñas de la estructura de la cromatina son complejos macromoleculares de ADN e histonas, llamados nucleosomas. Estos nucleosomas se encuentran tanto en cromatina como heterocromatina, están compuestos de ocho moléculas de histonas denominadas octameros que se agrupan como cuentas de collar, y esas cuentas en fibrillas de 30nm. Esas fibrillas a su vez forman dominios en formas de bucles, esos bucles comenzaran a condensarse y acumularse en paquetes de 700nm hasta formar un cromosoma de 1400nm. Cromosomas: Los cromosomas se forman a partir de fibras de cromatina, se pueden observar cromátides que están unidas por medio de un centrómero dándole un aspecto doble. Esta naturaleza doble es genera en la fase S del ciclo celular donde se produce el duplicado del ADN. El área de los extremos de los cromosomas se lo denomina telonero, que se acortan con cada división celular, que indica la vida útil de la célula. [ Las enzimas telomerasas añaden secuencias de nucleótidos repetidos al extremo del telómeros, haciendo que la vida útil de la célula se extienda.] En el ser humano se han contado 46 cromosomas los cuales 44 cromosomas son del tipo autosomas y los dos restantes son del tipo sexuales. Los dos cromosomas sexuales pueden ser X e Y. Estos son 23 pares de cromosomas homólogos, y cuando los cromosomas sexuales se expresan de forma homologa para el cromosoma XX el sexo del nuevo ser será femenino, de caso contrario cuando los cromosomas se presentes de forma heterogametica o sea XY el sexo
FASE S: Es donde se produce la fase de síntesis del ADN, aquí se duplica el material genético formándose nuevas cromátides que se harán evidentes en la profase o metafase de la división mitótica. La replicación se produce en sitios de nominados replicones a lo largo del ADN cromosómico El punto de control en esta fase vigila la calidad de la duplicación del ADN. FASE G2: Esta fase la célula examina su ADN en preparación para la mitosis. En esta fase la célula se agranda y reorganiza a nivel de sus estructuras, aquí se verifican dos puntos de control del daño del ADN no duplicado. FASE M: Esta fase se caracteriza por la cariocinesis y la citocinesis con previas etapas distinguibles. Regulación del ciclo celular: La regulación es impulsada por dos proteínas denominadas ciclinas y cinasas dependientes de ciclinas que contribuyen en el paso de los puntos de control de la división del ciclo celular. El aumento de la actividad del complejo Cdk-ciclina se logra mediante la acción estimuladora de las ciclinas y es equilibrado por la acción inhibitoria de proteínas como las inhibidoras de cinasas. Tipo de ciclina Proteína cinasas dependiente de la ciclina asociada Fase del ciclo celular sobre la que actúan Proteínas efectoras sobre las que actúan. Ciclina D Cdk 4 y Cdk 6 Progresión de la fase G Proteínas p supresora de tumores, proteína de susceptibilidad al retinoblastoma. Ciclina E Cdk 2 Ingreso en la fase S Proteína cinasas ATM o ATR, proteína supresora de tumores p Ciclina A Cdk 2 Progresión de la fase S Proteína de duplicación A, ADN- polimerasa, proteína de mantenimiento de mini cromosoma. Ciclina A Cdk 1 Fase S a fase G y entrada a la fase M Ciclina B Ciclina E Cdk 1 Progresión de la fase M Proteínas asociadas con cromatina , histona H1, láminas de núcleo, proteínas centrosomas, factores de transcripción, proteínas cinasas, caseína cinasas II, proteínas cinasas c-mos. Mitosis: Es un proceso de segregación cromosómico y división nuclear seguido por la división celular que produce dos células hijas con la misma cantidad de cromosomas y el mismo contenido del ADN que la célula progenitora. La mitosis posee 4 fases, la profase, metafase, anafase y la telofase. Profase: Cromosomas duplicados, orgánulos duplicados, ensamblado del uso mitótico, los cromosomas se anclan a ese uso mitótico, las cromátides hermanas se mantienen juntas por la proteína cohesina y por el centrómero. Se desintegra la membrana nuclear. Metafase: los cromosomas son redirigidos hacia el ecuador de la célula anclada a los
microtúbulos del uso mitótico. Se reconocen tres tipos de microtubulos: Microtubulos antrales, Microtubulos polares y microtubulos cinetocoricos. Anafase: Las proteínas dineinas arrastran los cromosomas en sentido hacia los polos celulares mediantes los microtubulos. Telofase: Los orgánulos se encuentras distribuidos, se degrada el uso mitótico, se reconstituye la membrana nuclear, los cromosomas toman aspecto de cromatina relajadas, los nucléolos reaparecen. La separación de las células hermanas se debe a un anillo contráctil compuesto por un fascículo de filamentos de actina. Meiosis: La meiosis incluye dos divisiones nucleares secuenciales, seguidas de divisiones celulares que producen gametos que contienen la mitad del número de cromosomas y la mitad del ADN encontrado en las somáticas. Proface I: se observa el apareamiento, la sinapsis y la recombinación genética. Contiene 5 fases: Leptoteno (condensación de la cromatina, y aparición de cromosomas), Cigoteno (aquí comienza la sinapsis y se forma el complejo sinaptotenico), Paquiteno (se completa la sinapsis y comienza la recombinación genética), Diploteno (los cromosomas homólogos comienzan a separarse y parecen estar conectados por medio de un quiasma), Diacinesis (los cromosomas homólogos se condensan y se acortan para alcanzar su espesor máximo, el nucléolo desaparece y la envoltura nuclear se desintegra). Metafase: los cromosomas son redirigidos hacia el ecuador de la célula anclada a los microtúbulos del uso mitótico. Se reconocen tres tipos de microtubulos: Microtubulos antrales, Microtubulos polares y microtubulos cinetocoricos. Anafase: Las proteínas dineinas arrastran los cromosomas en sentido hacia los polos celulares mediantes los microtubulos. Telofase: Los orgánulos se encuentras distribuidos, se degrada el uso mitótico, se reconstituye la membrana nuclear, los cromosomas toman aspecto de cromatina relajadas, los nucléolos reaparecen. La separación de las células hermanas se debe a un anillo contráctil compuesto por un fascículo de filamentos de actina. Meiosis II: la meiosis II es una división ecuatorial semejante a la mitosis, y durante esta fase la proteasa llamada separasa rompe los complejos de cohesina en la región del centrómero rompe la unión entre ambos centrómeros. Esta escisión separa las cromátides hermanasen la anafase II moviéndose a los polos celulares. El resto del proceso es idéntico que la mitosis.