¡Descarga comunicacion celular y más Apuntes en PDF de Psicobiología solo en Docsity!
En organismos multicelulares, las células que conforman tejidos especializados fun-
cionan en forma armónica y coordinada por medio de numerosos mecanismos finamen- te regulados. Muchos de estos mecanismos dependen de la comunicación entre células. Esta comunicación se produce cuando ciertas células emiten un mensaje a través de se- ñales químicas que ejercen su efecto en otras células y éstas, como consecuencia, inician una respuesta biológica. Las respuestas son muy variadas: transformaciones morfológicas, modificación en rutas metabólicas o cambios en la expresión de ciertos genes que llevan a la división, a la diferenciación e incluso a la muerte celular. Existen similitudes en estos mecanismos de comunicación intercelular entre distintos grupos de eucariontes multicelulares y también entre bacterias. Esas similitudes sugieren que es probable que el ancestro común a todos esos grupos ya tenía esas características. En organismos unicelulares hay mecanismos de comunicación entre individuos. Evolutiva- mente, cuando se adquirió la multicelularidad, estos mecanismos ya existentes en organis- mos unicelulares se habrían cooptado (véase cap. 19, Aptación, adaptación, exaptación) haciendo más eficiente la coordinación de funciones del organismo.
Mecanismos generales
de comunicación celular
Para analizar el tipo de comunicación que existe entre células de los distintos tejidos de un organismo se establece una clasificación, según la distancia que recorre la molécu-
Los mensajeros moleculares recorrían los vasos sanguíneos como autos en
una autopista. Atravesaban tibios paisajes iluminados por un resplandor difu-
so y ondulante, avanzaban por túneles oscuros que latían rítmicamente, se in-
ternaban por callejuelas cada vez más estrechas. Cuando por fin llegaban a
las profundidades de los tejidos, se acercaban a las delicadas formas que
emergían de las membranas celulares y ejecutaban en torno a ellas una sen-
sual danza exploratoria. Danzaban hasta encontrar a sus parejas perfectas.
Entonces se producía un reconocimiento íntimo y los mensajeros susurraban
sus mensajes al interior de las células. De inmediato se desencadenaba una
avalancha de reacciones y las células despertaban, del mismo modo que des-
pierta el durmiente cuando la luz del sol se derrama sobre su rostro.
Raúl A. Alzogaray
Capítulo 12
Comunicación celular
Capítulo 10. El flujo de información genética: los caminos del DNA a la proteína
Capítulo 11. La regulación de la expresión génica
Capítulo 12. Comunicación celular
Capítulo 13. Elementos genéticos móviles
Capítulo 14. La manipulación de la información genética
Capítulo 15. Desarrollo: la ejecución de un programa genético
Sección 3
Los genes en acción: estructura, expresión y control de la información genética
Capítulo 16. Genética, medicina y sociedad
la señal hasta alcanzar a la célula receptora. Si la molécula señal afecta só- lo a células receptoras que se encuentran en las cercanías de la célula emisora, el mecanismo se denomina comunicación paracrina. Tam- bién puede suceder que la célula receptora sea a su vez la célula emiso- ra, en cuyo caso el proceso se denomina comunicación autocrina. La comunicación entre células muy distantes en el organismo, en la que participan señales liberadas por ciertas células animales que via- jan por el torrente sanguíneo hasta alcanzar a la célula receptora –co- mo ocurre con las hormonas–, se denomina comunicación endocrina (véase cap. 34). La señal también puede recorrer largas distancias den- tro de la célula emisora, como es el caso de la señalización nerviosa. La estimulación de las neuronas produce impulsos eléctricos que llegan hasta la terminal del axón y causan la liberación de neurotransmisores (moléculas señal) al espacio sináptico donde interactúan con la célula receptora (véase cap. 31). También puede haber comunicación por contacto directo entre células. En estos casos, la molécula que actúa como señal puede per- manecer unida a la célula emisora durante su interacción con la célula receptora (en algunos tipos de respuesta inmunitaria) o desplazarse desde el citoplasma de la célula emisora al de una célula receptora ve- cina a través de verdaderos canales generados entre ambas (en mamí- feros, llamadas uniones comunicantes de tipo gap o brecha ) ( 1 fig. 12-1) (véase cap. 30, cuadro 30-1). El conocimiento actual sobre estos mecanismos es incompleto. Las clasificaciones que emplearemos se proponen sobre la base de aquellos procesos que se han descrito hasta el momento. Sin duda, muchos otros mecanismos y sobre todo la complejidad de sus interac- ciones aún no se comprenden. Pese a ello, los conceptos y los mode- los que presentaremos a lo largo de este capítulo ponen en evidencia la importancia que tiene no sólo el flujo de información del DNA hacia las proteínas, sino también aquella información que es transmitida a través de señales químicas, entre células, tejidos, y entre el organismo y su am- biente, en la expresión del fenotipo y en la continuidad de la vida.
El reconocimiento de la señal
En un organismo hay distintos tipos de señales; ciertas señales pueden disparar la división celular, mientras que otras pueden causar la liberación de una enzima para la digestión o llevar a la contracción de un músculo. Las señales son moléculas de diverso tipo. Pueden ser proteínas, péptidos pequeños, aminoácidos, nucleótidos, esteroides, vi- taminas, derivados de ácidos grasos y gases disueltos como el monóxi- do de carbono y el óxido nítrico. La interacción entre la señal y una mo- lécula que actúa como receptor da lugar a un complejo en el que la señal se denomina ligando ; de este modo, cuando la señal alcanza la célula blanco , se forma un complejo ligando-receptor. No todas las células son sensibles a las mismas señales; las interacciones son espe- cíficas de ciertos tejidos y están asociadas con funciones particulares. A su vez, las células no responden a un único tipo de señal, sino a un conjunto de señales. Es posible, a pesar de los grandes avances en este campo de investigación, que existan formas de respuesta a la enorme can- tidad de señales químicas existentes que aún se desconocen. ¿Por qué una molécula señal interactúa con un tipo de células en particular y no con otro? ¿Pueden diferentes moléculas señal transmitir su mensaje a la misma célula receptora? ¿Qué pasa con la molécula señal luego de la recepción del mensaje? Para contestar este tipo de preguntas debemos concentrar la atención en los dispo- sitivos y en los procesos que ocurren en la célula receptora de la señal.
La interacción de la molécula señal con su receptor
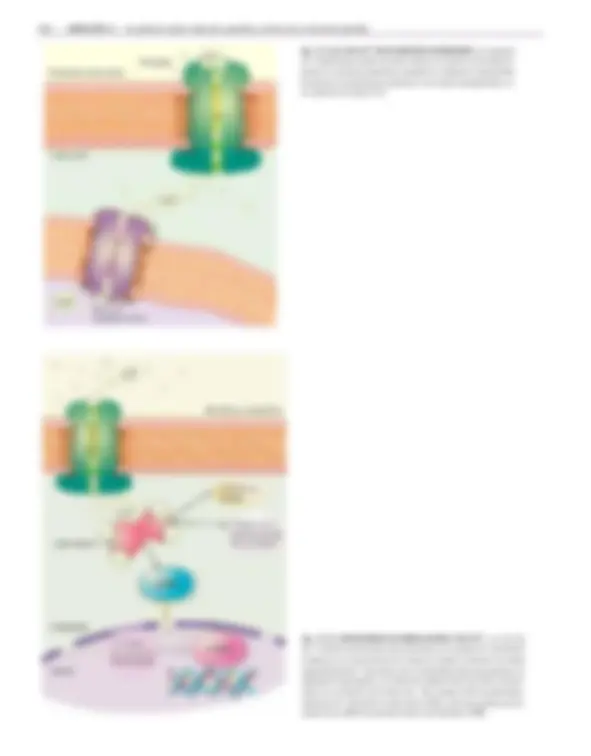
Una molécula actúa como señal cuando la célula receptora puede reconocer ese mensaje. Ese reconocimiento se produce por medio de los receptores, que son macromoléculas de estructura proteica localiza- dos ya sea en la membrana plasmática o en el interior de la célula ( 1 fig. 12-2).
Fig. 12-1. TIPOS DE COMUNICACIÓN CELULAR. La comunicación celular puede establecerse de distintas formas que incluyen: (a) la comunicación paracrina: la señal actúa sobre células vecinas; (b) endocrina: la molécula señal viaja por el torrente san- guíneo y alcanza células lejanas; (c) autocrina: la molécula señal llega a la misma célu- la de la cual salió; (d) neurotransmisión: la señal eléctrica viaja por la célula emisora (la neurona) hasta la terminal sináptica donde se produce la liberación de una molécula se-
ñal al espacio sináptico desde donde es captada por la célula receptora; (e) contactos célula-célula: la molécula señal permanece anclada a la membrana de la célula emiso- ra mientras interactúa con la célula receptora y (f) en células de mamíferos, mediante uniones comunicantes o gap function, la molécula señal pasa desde la célula emisora a la célula receptora por difusión dentro del citoplasma.
Capítulo 12: Comunicación celular / 231
Figura interactiva
Capítulo 12: Comunicación celular / 233
Cuando estos complejos diméricos interactúan con las secuencias específicas del DNA, reclutan a otras proteínas coactivadoras que, entre otras funciones, facilitan el acceso de la RNA polimerasa II, responsable de la transcripción del gen (véase cap. 10, cuadro 10-1). Si bien muchas células tienen los mismos receptores intracelulares, la combinación co- rrecta de proteínas coactivadoras y complejos ligando-receptor es la que hace que en cada célula se activen determinados genes y no otros. En algunos casos, como en el de las hormonas tiroideas o el de los retinoides, los receptores no se encuentran en el citoplasma, sino unidos al DNA pero, en ausencia del ligando, no presentan actividad transcripcional. Cuando interactúan con la hormona, se producen cam- bios conformacionales que provocan su activación.
Los receptores de membrana
Receptores de membrana asociados con canales
iónicos
Algunos receptores de membrana permiten el pasaje de iones al interior o el exterior de la célula. Estos receptores, denominados
ionotrópicos , forman parte de canales iónicos (véase cap. 3, Los ca- nales). En general, los receptores asociados con canales iónicos res- ponden a neurotransmisores y están involucrados en el control de la contracción muscular y en la transferencia neuronal de informa- ción que ocurre en los procesos de memoria y de excitabilidad neu- ronal. En ausencia de estímulo, los canales iónicos se abren y se cierran con una frecuencia determinada y permiten el pasaje de determinados iones. La unión del ligando con su receptor específico produce cambios en la estructura del canal que alteran su frecuencia de apertura, lo cual modifica transitoriamente la permeabilidad de la membrana a determi- nado ion y de esa forma altera el potencial eléctrico de la membrana y la excitabilidad de la célula (véase cap. 31). Esta variación de potencial eléctrico induce la respuesta biológica. Un ejemplo de receptor con función de canal iónico para el Na + es el receptor de acetilcolina. La unión de acetilcolina aumenta la fre- cuencia de apertura del canal y así permite un ingreso mayor de iones Na +^ en la célula. La entrada de especies cargadas positivamente produce la despolarización de la membrana de la célula receptora ( 1 fig. 12-4) ( 2 recuadro 12-2, Patch clamp ).
Fig. 12-3. RECEPTORES CON ACTIVIDAD DE FACTOR DE TRANSCRIPCIÓN. Este tipo de receptores pertenece a la familia de factores de transcripción activados por li- gando. (a) Desde su extremo amino hasta el carboxilo se ubican los dominios funciona- les en el siguiente orden: A/B: dominio que interactúa con coactivadores de la transcrip- ción; C: dominio de unión al DNA; D: señal de localización nuclear y E/F: dominios de unión al ligando, de interacci ón con coactivadores de la transcripción, de unión a las cha- peronas y de dimerización. (b) Mecanismo de activación del receptor de esteroides. La lle-
gada del ligando provoca la transformación del receptor que se encuentra unido a pro- teínas chaperonas. Este complejo se trasloca al núcleo y se asocia con otro complejo si- milar que se une a secuencias específicas del DNA presentes en los promotores de los ge- nes blanco. Cuando los complejos diméricos interactúan con el DNA, participan otras pro- teínas coactivadoras que, entre otras funciones, facilitan la transcripción del gen por la RNA polimerasa II.
Receptores de membrana asociados con enzimas
Dentro de este grupo de receptores asociados con enzimas se dis- tinguen dos tipos: los que se activan y funcionan directamente como enzimas y los que interactúan con enzimas que se encuentran en el la- do interno de la membrana plasmática, activándolas. Todos estos recep- tores son proteínas que atraviesan la membrana plasmática. En todos los casos tienen un sitio de unión para el ligando fuera de la célula y desarrollan actividad catalítica o presentan un sitio de unión para las en- zimas en el lado interno. La actividad enzimática asociada con la gran mayoría de estos receptores es de proteincinasa (véase cap. 4, El ATP en acción), ya que catalizan la transferencia de grupos fosfato del ATP al grupo hidroxilo de ciertos aminoácidos (tirosina, serina o treonina) de determinadas proteínas receptoras. Como vimos, la fosforilación de una proteína provoca cambios en su conformación que influyen directa- mente sobre su función. Los receptores con actividad de proteincinasa participan en vías de transducción, es decir, en cascadas de señalización, cuyas respuestas biológicas se relacionan con la regulación de la proliferación, la diferen- ciación, la producción de la matriz extracelular, la reparación de tejidos y la regulación inmunitaria. También participan en mecanismos que modulan el metabolismo y la migración celular. ¿Cómo es la cascada de señalización que se origina en estos receptores? Luego de la interacción del ligando con su receptor se produce la unión de varios complejos li- gando-receptor en la membrana, lo que provoca la yuxtaposición de sus dominios catalíticos que permite su fosforilación recíproca en tirosina (receptores con actividad de tirosina cinasa) o en serina o treonina (re- ceptores con actividad de serina/treonina-cinasa). La autofosforilación de los receptores incrementa aún más su actividad de cinasa y permi- te el reclutamiento de proteínas señalizadoras citosólicas que interac-
túan con los dominios catalíticos de los receptores y transmiten el men- saje al interior de la célula ( 1 fig. 12-5). En la actualidad se conocen más de 50 miembros de la familia de receptores con actividad de tirosina cinasa. Entre ellos se encuentran re-
234 / S E CC IÓN 3 / Los genes en acción: estructura, expresión y control de la información genética
Recuadro 12-
Patch clamp
La introducción de la técnica de patch clamp por Edwin Neher y Bert Sak-
manna hizo posible el estudio individual de canales iónicos en la membra-
na. Ambos investigadores recibieron el Premio Nobel de Fisiología en 1991
por su contribución al conocimiento en esta área.
La técnica de patch clamp se basa en la utilización de una pipeta de vidrio
de borde muy liso cuya punta, de entre 1 y 5 micrómetros de diámetro,
se apoya sobre el exterior de una célula. Al aplicar una succión muy leve,
se forma un sello muy firme entre la punta de la pipeta y la membrana.
Dentro de la punta de la pipeta queda entonces una porción de la mem-
brana que sólo tiene unos pocos canales. Si se dispone de un dispositivo
de medida de potencial eléctrico, se puede entonces determinar el poten-
cial de la membrana en distintas condiciones experimentales y analizar la
apertura y el cierre de canales individuales.
Técnica de patch clamp_. La figura muestra una fotomicrografía en la que se aprecia la pipeta apoyada sobre la membrana plasmática de una célula don- de puede observarse un canal iónico._
Fig. 12-4. RECEPTORES CON ACTIVIDAD DE CANALES IÓNICOS. El receptor de acetilcolina atraviesa la membrana plasmática de lado a lado. En su centro se abre un canal que permite la entrada del ion Na+^. En este caso, la unión de dos moléculas de acetilcolina produce cambios en la estructura del canal que alteran su frecuencia de apertura y así permite la entrada de Na +^ en la célula. De esta manera, se altera el potencial eléctrico de la membrana de la célula receptora, lo cual provoca su despolari- zación.
tesis de estradiol) y el de la hormona angiotensina II (que estimula la síntesis y la liberación del mineralocorticoide aldosterona) (véanse caps. 34 y 41). Este tipo de mecanismo involucra procesos de terminación de la señal. Por ejemplo, la exposición prolongada del ligando al receptor pro- mueve su inactivación. La desensibilización del receptor lo desacopla del transductor (la proteína G) e induce su internalización y finalmente su degradación, y así disminuye el número de receptores expuestos en la superficie celular.
Transmisión de la señal al interior de la célula
Cuando el ligando interactúa con su receptor de membrana, la se- ñal es transmitida al interior de la célula, lo que genera la respuesta bio- lógica correspondiente. Este proceso involucra una cascada de eventos, como la síntesis de pequeñas moléculas ( segundos mensajeros ) y la activación o la inhibición de enzimas, que constituyen el mecanismo
236 / S E CC IÓN 3 / Los genes en acción: estructura, expresión y control de la información genética
Fig. 12-6. PRODUCCIÓN DE cGMP A PARTIR DE GTP.
Fig. 12-7. RECEPTORES ACOPLADOS A PROTEÍNAS G. (a) Este tipo de receptores consiste en un polipéptido que atraviesa la membrana plasmática siete veces. Este esquema muestra la topolo- gía del receptor β 2 -adrenérgico que se ha estudiado ampliamente. (b) Una señal interactúa con el re- ceptor que se activa y cambia de forma. La proteína G inactiva se une al receptor y se activa. Luego se desplaza hacia otra proteína de membrana que se encuentra en estado inactivo. Cuando la pro- teína G se une a esta proteína, altera su actividad. Esto conduce a una respuesta. (c) Estructura de una proteína G trimérica unida a GDP. Este modelo se obtuvo sobre la base del análisis por cristalo- grafía de rayos X.
(b)
(a)
(c)
Galería de imágenes
Capítulo 12: Comunicación celular / 237
Recuadro 12-
Las proteínas G heterotriméricas: estructura, regulación y mecanismos de señalización
I
Las proteínas G funcionan como un acople entre receptores y enzimas
o canales iónicos. Su denominación se debe a Alfred G. Gilman, uno
de los investigadores pioneros en el área quien, junto con sus colabo-
radores del Hospital Clínico de la Universidad de Texas en Dallas, Esta-
dos Unidos, recibió el Premio Nobel de Fisiología en 1994 por haber
caracterizado en detalle esta proteína. En la actualidad se llaman pro-
teínas G a aquellas que intervienen en el metabolismo de nucleótidos
de guanina.
Todas las proteínas G heterotriméricas presentan tres subunidades di-
ferentes, α , β y γ , y un funcionamiento similar. Como vimos, en su
estado inactivo, la subunidad α de la proteína G lleva GDP unido. La
interacción con el complejo ligando-receptor produce la liberación del
GDP y la unión de GTP. Esto produce un cambio conformacional que
induce la disociación del trímero en dos componentes, la subunidad α
y el complejo βγ. La subunidad α puede entonces interactuar con
otras proteínas. El complejo βγ no cambia su conformación, pero la
zona que antes estaba enmascarada por la subunidad α queda libre
para interactuar con el otro grupo de proteínas. Este proceso es transi-
torio porque la subunidad α es capaz de hidrolizar el GTP unido a GDP
(esto se llama actividad de GTPasa) y una vez que esto ocurre se rea-
socian las subunidades y se reconstituye la proteína G inactiva. Este
mecanismo es de suma relevancia fisiológica, dado que permite volver
a la situación basal una vez transmitido el mensaje.
Cualquier compuesto que pueda bloquear la función de la subunidad
α podría afectar la transducción de la señal. Un ejemplo de este caso
es la acción de la toxina producida por la bacteria Vibrio cholerae que
inhibe la actividad GTPasa de la subunidad αs ; de esta forma, la subu-
nidad α se encuentra siempre unida al GTP y por lo tanto siempre per-
manece activa. En las células del intestino, la consecuencia es la pérdi-
da de agua y cloruro característica de la diarrea del cólera.
Otro ejemplo es la toxina producida por la bacteria Bordetella pertus-
sis , que causa la tos ferina o convulsiva. Esta toxina modifica estructu-
ralmente a la subunidad α, lo cual impide la asociación de la proteína
G al complejo ligando-receptor y como consecuencia bloquea la trans-
ducción de esa señal.
Proteínas G heterotriméricas. Cuando el ligando interactúa con el re- ceptor, este complejo se asocia con la proteína G y le provoca un cam- bio conformacional que induce el intercambio del GDP (que en condi- ciones basales está unido a la subunidad α ) por GTP. En consecuen- cia, el trímero se disocia en α -GTP y β γ. La subunidad α puede enton- ces interactuar con proteínas efectoras. Este proceso se cierra (aunque no se observa en la figura) cuando la subunidad α hidroliza el GTP uni- do a GDP (mediante su actividad de GTPasa), se reasocian las subu- nidades y se reconstituye la proteína G inactiva.
Figura animada
Capítulo 12: Comunicación celular / 239
En el grupo de pequeñas proteínas G se encuentran las denomi- nadas Ras, que están involucradas en el control de procesos de prolife- ración y diferenciación celular. La proteína Ras es una proteína de mem- brana que actúa como intermediario entre una señal, por ejemplo un factor de crecimiento, y la división celular ( 1 fig. 12-9). En algunos ti- pos de cáncer, la proteína Ras anormal siempre está activa, aunque no haya señal, y causa la división continua de la célula (véase cap. 7). Tam- bién pertenecen a este grupo las proteínas Rho, que participan en la re- gulación de la actividad de proteínas del citoesqueleto. El grupo deno- minado Rab, participa en el control del transporte de proteínas y vesí- culas dentro de la célula y el grupo Ran está implicado en la regulación de las transiciones del ciclo celular.
Los segundos mensajeros
Un segundo mensajero es una molécula pequeña que se gene- ra rápidamente (en 1-2 minutos) y en gran cantidad en respuesta a la activación de un receptor y que se aleja pronto de su lugar de sín- tesis llevando la señal a otras partes de la célula, donde interactúa con proteínas específicas. El incremento en los niveles del segundo men- sajero precede a cualquier otra etapa del mecanismo de señalización. El segundo mensajero produce la activación de enzimas como las proteincinasas y de esta forma participa en la amplificación de la se- ñal iniciada por el ligando constituyendo un intermediario en la gene- ración de la respuesta biológica.
El número de segundos mensajeros parece ser muy pequeño frente a la variedad de moléculas señal que controlan las distintas respuestas bio- lógicas. Es decir que las rutas internas de transducción de los mensajes den- tro de las células son sorprendentemente universales. Es por ello que la identidad de las sustancias que actúan como segundos mensajeros es de importancia fundamental para comprender el control de la actividad celular. A continuación presentaremos distintos tipos de segundos mensa- jeros.
Nucleótidos cíclicos
El concepto de segundo mensajero se originó en 1957, cuando se identificó como tal al nucleótido monofosfato cíclico de adenosina (cAMP), una molécula soluble y resistente al calor que se acumula en el tejido hepático luego de su estimulación con glucagón o adrenalina (véase cap. 34, Médula suprarrenal: adrenalina y noradrenalina). Bas- tante tiempo después se caracterizaron las enzimas que catalizan su síntesis (adenilato ciclasa) y su inactivación (fosfodiesterasa). La estimulación del receptor produce en unos pocos segundos un incremento transitorio de los niveles de cAMP en más de 20 veces. El nucleótido es transformado en 5’-AMP por la fosfodiesterasa, y así se cierra el ciclo de señalización ( 1 fig. 12-10). El aumento de cAMP pro- ducido como respuesta a muchas señales extracelulares se debe a un incremento en la actividad de la adenilato ciclasa, más que a una inhi- bición en la actividad de fosfodiesterasa.
Fig. 12-9. REACCIONES EN CASCADA MEDIADAS POR LA PROTEÍNA Ras. Las proteínas Ras pertenecen a la familia de las pequeñas proteínas G monoméricas. Como otras proteínas G, Ras existe en dos estados conformacionales diferentes, activo cuando lleva unido GTP e inactivo cuando tiene GDP. La activación del receptor asegura el reclu- tamiento de un complejo denominado Grb2/Sos. La proteína de los vertebrados Grb2 es una proteína adaptadora que se asocia con el receptor con actividad de tirosina cinasa
fosforilado. A Grb2 se acopla la proteína Sos, que interactúa con Ras estimulando la diso- ciación de GDP y la unión de GTP. La interacción del receptor fosforilado con otra proteí- na denominada GAP incrementa la hidrólisis del GTP unido a Ras y, por lo tanto, su inac- tivación. Ras activada se fija a la proteincinasa Raf que se activa. Raf activada se une y fosforila a ERK, que se trasloca al núcleo y es un activador de la transcripción de los ge- nes tempranos.
La consecuencia del aumento de cAMP depende del tipo de célu- la que se esté estudiando. Sin embargo, si diferentes señales extracelu- lares producen un aumento de cAMP en la misma célula, es posible que la respuesta biológica sea la misma. La mayor parte de las vías de señalización en las que actúa el cAMP involucran también a la proteincinasa A (PKA). La actividad de PKA es similar a las de las cinasas de serina o treonina que describimos antes. En el estado inactivo, esta enzima consta de dos subunidades re- guladoras y dos subunidades catalíticas. En este estado, las subunida- des reguladoras bloquean el sitio catalítico. La unión de cuatro molécu- las de cAMP a las dos subunidades reguladoras de la PKA provoca un
cambio conformacional que desbloquea las subunidades catalíticas y éstas, entonces, pueden fosforilar sus sustratos ( 1 fig. 12-11). Los sustratos naturales de la PKA incluyen enzimas de vías meta- bólicas como la glucógeno sintetasa –que participa en la síntesis de glu- cógeno– y la piruvato cinasa –que actúa en la glucólisis– y algunos ca- nales iónicos (canal de Ca 2+^ ). La fosforilación de enzimas es un ejem- plo de regulación rápida del metabolismo. Por ejemplo, la fosforilación de las enzimas que participan del metabolismo del glucógeno entre dos comidas activa su degradación. La PKA también actúa a largo plazo iniciando la transcripción de genes específicos. En este caso, la PKA fosforila un factor de transcrip-
240 / S E CC IÓN 3 / Los genes en acción: estructura, expresión y control de la información genética
Fig. 12-10. SEÑALIZACIÓN POR NUCLEÓTIDOS CÍCLICOS. (a) El cAMP es sinte- tizado a partir de ATP en una reacción catalizada por la enzima adenilato ciclasa. (b) La disociación de la proteína G heterotrimérica y el intercambio del GDP unido a la subuni- dad α por GTP produce su activación. La subunidad α activada activa la adenilato cicla-
sa que cataliza la síntesis de cAMP a partir de ATP. El incremento en los niveles de cAMP es transitorio, dado que este nucleótido es transformado en 5’-AMP por la fosfodiestera- sa, y así se cierra el ciclo de señalización.
Ion calcio
El ion calcio (Ca 2+^ ) participa como segundo mensajero en diver- sos procesos biológicos, entre los que se encuentran la contracción muscular, los movimientos de cromosomas, la liberación de neurotrans- misores, la endocitosis y la exocitosis (véase cap. 3, Intercambio a tra- vés de vesículas). Un estímulo adecuado produce un incremento en los niveles cito- plasmáticos de Ca2+^ en forma transitoria. Este aumento puede deberse a la apertura de canales presentes en la membrana plasmática, lo que permite la entrada de ese ion desde el medio extracelular o la libera- ción de Ca 2+^ desde depósitos intracelulares como los del retículo endo- plasmático ( 1 fig. 12-13). La apertura de los canales de calcio depende de la unión de un li- gando (como en el caso de los receptores acoplados a canales iónicos) o puede producirse como consecuencia de cambios en el potencial de membrana. Como veremos más adelante, los canales de Ca 2+^ del retí- culo endoplasmático son activados por el lípido inositol trifosfato ( IP 3 ). El aumento de la concentración de calcio en el citosol activa vías de señalización. Para que pueda transmitir el mensaje, el calcio debe unirse y modificar proteínas específicas. Sin embargo, no todas las pro- teínas que pueden unirse al calcio participan de la transducción de se- ñales. Las proteínas involucradas en la transducción deben poder res- ponder a aumentos temporarios y, muchas veces localizados, de la con- centración del ion. Entre las proteínas que pueden cumplir esta función
se encuentra la calmodulina. Si bien la calmodulina no tiene ninguna actividad enzimática, el complejo Ca2+^ -calmodulina puede regular la ac- tividad de más de 100 enzimas diferentes ( 1 fig. 12-14). Una de ellas es la fosfodiesterasa de cAMP. De hecho, la calmodulina fue identifica- da durante la purificación de esa fosfodiesterasa. La calmodulina funciona como un receptor intracelular de calcio, da- do que une cuatro iones calcio por molécula. La unión del Ca2+^ modifica la conformación de la calmodulina y le permite la interacción con otras proteínas. Entre ellas podemos citar la adenilato ciclasa, la óxido nítrico sintetasa, proteínas del citoesqueleto, proteínas transportadoras de calcio como la Ca2+-ATPasa y algunas proteincinasas como la fosforilasa cinasa (que participa en el metabolismo de glucógeno), la cinasa de la cadena liviana de miosina (que participa en la contracción muscular) y otras cina- sas multifuncionales dependientes de Ca2+-calmodulina (CaMK). La CaMK más estudiada es la CaMK II. Cuando los niveles de calcio cito- sólicos son bajos, esta enzima no es activa porque tiene una zona au- toinhibitoria que bloquea el acceso del sustrato al sitio catalítico. Cuan- do los niveles de calcio aumentan, el ion se une a la calmodulina y el complejo Ca2+^ -calmodulina se une a la enzima CaMK II. Esto desblo- quea el sitio catalítico. La unión de Ca 2+^ -calmodulina a la enzima per- mite su autofosforilación, lo que incrementa aún más su actividad de ci- nasa. Entre las proteínas que son sustrato de CaMK II se encuentran factores de transcripción ( 2 recuadro 12-4, ¿Cómo medir el aumento de calcio citoplasmástico? ).
242 / S E CC IÓN 3 / Los genes en acción: estructura, expresión y control de la información genética
Fig. 12-12. EL cGMP Y LA ACTIVACIÓN DE LA PROTEINCINASA G (PKG). El au- mento en los niveles de cGMP, inducido por receptores con actividad de guanilato cicla- sa, provoca la activación de proteínas como la proteincinasa PKG. El cGMP también inte-
ractúa con canales iónicos de cationes monovalentes y divalentes, en particular relacio- nados con el sistema visual. Los niveles de cGMP también son regulados por fosfodieste- rasas que catalizan su transformación en 5’-GMP.
Capítulo 12: Comunicación celular / 243
Lípidos
Otro grupo de segundos mensajeros se produce a partir de fosfo- lípidos que constituyen la membrana plasmática. En particular, la ruptu- ra del fosfatidil inositol difosfato libera por un lado inositol trifosfato (IP 3 ) que conformaba su cabeza polar y por el otro, diacilglicerol (DAG) (que constituía la cola no polar). La enzima que cataliza la escisión del fosfolípido se denomina fosfolipasa C ( 1 fig. 12-15). La liberación de los mediadores IP 3 y DAG conduce a la activación de otro eslabón de la cadena de señalización: la proteincinasa C (PKC). La PKC es una cinasa de serina y treonina que participa en cami- nos de señalización involucrados en la proliferación celular. Como vi- mos, el IP 3 permite la liberación de Ca2+^ de los depósitos intracelulares al citosol. El Ca2+^ se une directamente a la PKC que a su vez es ancla- da a la membrana plasmática al unirse al DAG. Esta interacción aumen- ta la afinidad de la PKC por el fosfolípido fosfatidilserina (PS). Cuando la PKC se une a la membrana plasmática, cambia su conformación y se desbloquea la región catalítica al separarse de la región regulado- ra ( 1 fig. 12-1 6 ). Las proteínas sustrato de la fosforilación por PKC varían según el ti- po celular, pero entre ellas se encuentran los receptores β-adrenérgicos y el receptor del factor de crecimiento epidérmico.
Transducción del mensaje por cascadas de activación
de cinasas
La fosforilación y desfosforilación de proteínas es parte de la trans- ducción del mensaje. Las vías metabólicas pueden ser reguladas por
efectores alostéricos (véase cap. 4, Interacciones alostéricas) o por mo- dificación covalente de sus enzimas clave. Sin embargo, la fosforilación es principalmente una respuesta a la influencia extracelular mediada por receptores, mientras que los efectores alostéricos responden a con- diciones intracelulares. Como vimos desde el comienzo del capítulo, muchas vías de se- ñalización involucran mecanismos de fosforilación y desfosforilación que regulan la actividad de proteínas (enzimas o no). Esto significa que la mayoría de los mensajes que se transmiten al interior celular produ- cen la activación de proteincinasas que regulan su función mediante la fosforilación de diversos sustratos. Estas cinasas son activadas por los segundos mensajeros (como vimos en los casos de PKA, PKG, PKC y CaMK II) o bien por fosforilación mediante una cascada de activación de cinasas.
Interacción entre las distintas vías de señalización
¿Puede una molécula señal determinada regular la respuesta bio- lógica de una célula a través de más de un camino de transducción de señal? Si bien a lo largo de este capítulo en general hemos descrito las principales vías de transducción como cadenas lineales a través de las cuales fluye la información, también hemos visto que existen puntos potenciales de regulación y de intersección entre los distintos eslabo- nes de la cadena. Esto permite que se establezca una comunicación cruzada entre dos o más vías de transducción de señales. Todas las cé- lulas reciben constantemente múltiples señales del medio externo que son reconocidas por receptores específicos. Una misma molécula señal puede ser reconocida en la misma célula por receptores diferentes aso-
Recuadro 12-
¿Cómo medir el aumento de calcio citoplasmático?
Un gran avance en los métodos utilizados para las determinaciones de cal-
cio intracelulares se produjo cuando Roger Tsien diseñó una serie de indi-
cadores sintéticos de calcio. Se trata de compuestos que permiten detec-
tar ciertas fluctuaciones de las concentraciones de calcio porque producen
una fluorescencia característica. Estos indicadores, uno de los más conoci-
dos es el Fura-2, pueden atravesar fácilmente la membrana plasmática y
se utilizan para estudiar variaciones en la concentración de calcio citosóli-
ca en una célula aislada sometida a un estímulo determinado. Otro indi-
cador de calcio es la proteína aequorina (proteína luminiscente de inver-
tebrados marinos), que mediante la metodología de DNA recombinante
se ha logrado introducir en determinados compartimientos intracelulares
para estudiar variaciones en la concentración del ion en organelas intrace-
lulares.
Las células HeLa (de tumor de epitelio cervical humano) fueron teñidas con el indica- dor Fura-2. Se obtuvieron dos imágenes de fluorescencia midiendo a diferentes longi- tudes de onda (340nm y 380nm) y luego se calculó la relación entre ambas para ob- tener los valores de concentración de calcio. En la foto se asignan diferentes colores a distintas concentraciones de calcio (de azul a rojo indica de menor a mayor concen- tración). La imagen de fluorescencia se superpone con la imagen obtenida por micros- copio electrónico.
Capítulo 12: Comunicación celular / 245
Fig. 12-15. LOS FOSFOLÍPIDOS SON SEGUNDOS MENSAJEROS. La hidrólisis del fosfatidilinositol difosfato li- bera por un lado el inositol trifosfato (IP 3 ) y por el otro diacil- glicerol (DAG). La fosfolipasa C es la enzima que cataliza esta reacción. Esta enzima puede ser activada por proteínas Gq, co- mo se muestra en el esquema, o mediante la interacción con receptores. El IP 3 liberado al citoplasma interactúa con recep- tores localizados en los depósitos de Ca2+^ del retículo endo- plasmático y estimula la liberación de este ion al citoplasma. Sobre el margen superior izquierdo del esquema se observa la estructura del fosfatidilinositol difosfato.
Fig. 12-16. ACTIVACIÓN DE LA PROTEINCI- NASA C (PKC). En estado inactivo, la región cata- lítica de la PKC se encuentra bloqueada por una re- gión llamada de seudosustrato (rectángulo verde). El Ca 2+^ citosólico se une directamente a la PKC, que a su vez se ancla a la membrana plasmática por in- teracción con el DAG y el fosfolípido fosfatidilserina (PS). La PKC unida a la membrana plasmática está activa porque cambia su conformación y desblo- quea la región catalítica, al separarse de la región de seudosustrato.
Galería de imágenes
246 / S E CC IÓN 3 / Los genes en acción: estructura, expresión y control de la información genética
ciados con distintos mecanismos de transducción de la señal. En ese sentido puede pensarse que la respuesta celular está conformada por un circuito complejo de activación que involucra más de un camino de señalización. Por otra parte, también puede ocurrir que un mismo re- ceptor active diferentes vías. De esta manera, podríamos mencionar muchos ejemplos de co- municación cruzada, pero tengamos en cuenta que para que esa tra- ma de interacciones se establezca tiene que haber puntos de con- tacto dentro de la célula que permitan que las proteínas que interac- túen se encuentren. Aún no resulta claro cómo una célula individual produce una respuesta específica a tantas señales extracelulares di- ferentes, muchas de las cuales utilizan el mismo tipo de receptores y activan muchas de las mismas vías de señalización. Una estructu- ra asociada con este tipo de procesos de transducción de señales es la de las llamadas “proteínas de andamiaje” sobre las que se organi- zan las proteínas que participan de la vía de señalización. Este tipo de estructura hace que la transducción del mensaje sea más preci- sa, rápida y eficiente y además permite que las interacciones sean específicas. En algunos casos, estos complejos son transitorios y se desarman luego de cumplir su función.
El final de la vía: la respuesta biológica
Cualquiera que sea el mecanismo que interviene en la transmisión de la señal extracelular, la consecuencia es una respuesta por parte de la célula blanco. Esta respuesta es una alteración que puede ser el resultado de la activación o de la inhibición de alguna vía metabólica, tal como el cambio en la utilización de combustibles durante el ayuno nocturno, como consecuencia del incremento de la relación hormonal glucagón/insulina. Estos cambios pueden ser rápidos, cuando se modifica la actividad de las enzimas de la vía, o lentos, cuando se modifican los niveles de expresión de esas enzimas. Modificaciones en la forma o en el movimiento de las células pueden ser otro tipo de respuesta. Como vimos, existe un alto grado de complejidad en los mecanismos de transducción disparados por diferentes señales. Es por ello que defec- tos en el reconocimiento ligando-receptor o en el sistema de señalización pueden producir alteraciones funcionales cuyas consecuencias son muy diversas, como por ejemplo la alteración en el grado de proliferación celu- lar, la transformación de un fenotipo celular normal a uno canceroso o la muerte celular. A lo largo de este texto veremos que el papel de estos procesos en el control de la diferenciación, de la proliferación y del crecimiento celular es esencial para el desarrollo y el funcionamiento normales del organismo.
En síntesis
Mecanismos generales de comunicación celular
- La comunicación celular puede establecerse de distintas formas que incluyen: a) la comunicación paracrina: la señal actúa sobre células vecinas; b) endocrina: la señal viaja por el torrente sanguí- neo y alcanza células lejanas; c) autocrina: la señal llega a la mis- ma célula de la cual salió; d) neurotransmisión: la señal es libe- rada por la célula emisora al espacio sináptico, donde es captada por la célula receptora; e) contactos célula-célula: la señal perma- nece anclada a la membrana de la célula emisora mientras inte- ractúa con la célula receptora y f) mediante gaps : la señal se di- funde desde la célula emisora a la receptora.
El reconocimiento de la señal
- En cada organismo existen distintos tipos de señales químicas que reciben el nombre de ligandos y forman complejos con re- ceptores específicos. Cada tipo celular es sensible a distintas se- ñales y cada interacción ligando-receptor está asociada con una función particular. Cada célula responde a un conjunto de se- ñales.
- El complejo ligando-receptor transmite el mensaje al interior de la célula e inicia un camino que lleva a la ejecución de una respues- ta biológica específica. El proceso completo constituye la trans- ducción de la señal.
- Ciertas moléculas pequeñas y/o hidrófobas atraviesan la membrana celular y se unen a receptores internos. Estos com- plejos suelen unirse al DNA y actuar como factores de trans- cripción. 5. Los receptores de membrana son variados. Pueden formar parte de canales iónicos, presentar actividad enzimática o estar asocia- dos con enzimas. Existen receptores que activan una proteína adaptadora, la proteína G, que transmite el mensaje al siguiente intermediario.
Los receptores intracelulares
- Los glucocorticoides y los mineralocorticoides, las hormonas se- xuales y las hormonas tiroideas son ejemplos de ligandos que se unen a receptores intracelulares que actúan como factores de transcripción. Todos los receptores intracelulares tienen una zona de unión al DNA y otra de reconocimiento del ligando.
- Las proteínas chaperonas mantienen el plegamiento específico de los receptores que se encuentran libres en el citoplasma. Lue- go de la unión de una hormona al receptor, el complejo se trans- loca al núcleo, se separa de la chaperona y forma un dímero con otro complejo similar. Estos dímeros interactúan con secuencias específicas del DNA, disparando la transcripción génica.
Los receptores de membrana
- Los receptores ionotrópicos se encuentran en la membrana celu- lar y permiten el pasaje de iones al interior o al exterior de la cé- lula. En general, responden a neurotransmisores y controlan pro- cesos de contracción muscular y de transferencia neuronal de in- formación.
- Existen dos tipos de receptores de membrana asociados con en- zimas: los que se activan y funcionan como enzimas y los que ac-