


































Prepara tus exámenes y mejora tus resultados gracias a la gran cantidad de recursos disponibles en Docsity

Gana puntos ayudando a otros estudiantes o consíguelos activando un Plan Premium

Prepara tus exámenes
Prepara tus exámenes y mejora tus resultados gracias a la gran cantidad de recursos disponibles en Docsity
Prepara tus exámenes con los documentos que comparten otros estudiantes como tú en Docsity
Encuentra los documentos específicos para los exámenes de tu universidad
Estudia con lecciones y exámenes resueltos basados en los programas académicos de las mejores universidades
Responde a preguntas de exámenes reales y pon a prueba tu preparación
Consigue puntos base para descargar
Gana puntos ayudando a otros estudiantes o consíguelos activando un Plan Premium
Comunidad
Pide ayuda a la comunidad y resuelve tus dudas de estudio
Ebooks gratuitos
Descarga nuestras guías gratuitas sobre técnicas de estudio, métodos para controlar la ansiedad y consejos para la tesis preparadas por los tutores de Docsity
En este documento encontraras una descripción de la conformación de los lipidos
Tipo: Apuntes
1 / 44

Esta página no es visible en la vista previa
¡No te pierdas las partes importantes!
Denominamos lípidos a un conjunto muy heterogéneo de biomoléculas cuya característica distintiva aunque no exclusiva ni general es la insolubilidad en agua , siendo por el contrario, solubles en disolventes orgánicos (benceno, cloroformo, éter, hexano, etc.).
En muchos lípidos, esta definición se aplica únicamente a una parte de la molécula, y en otros casos, la definición no es del todo satisfactoria, ya que pueden existir lípidos soluble en agua (como los gangliósidos, por ejemplo), y a la vez existen otras biomoléculas insolubles en agua y que no son lípidos (carbohidratos como la quitina y la celulosa, o las escleroproteínas).
Los lípidos pueden encontrarse unidos covalentemente con otras biomoléculas como en el caso de los glicolípidos (presentes en las membranas biológicas), las proteínas aciladas (unidas a algún ácido graso) o las proteínas preniladas (unidas a lípidos de tipo isoprenoide).
También son numerosas las asociaciones no covalentes de los lípidos con otras biomoléculas , como en el caso de las lipoproteínas y de las estructuras de membrana.
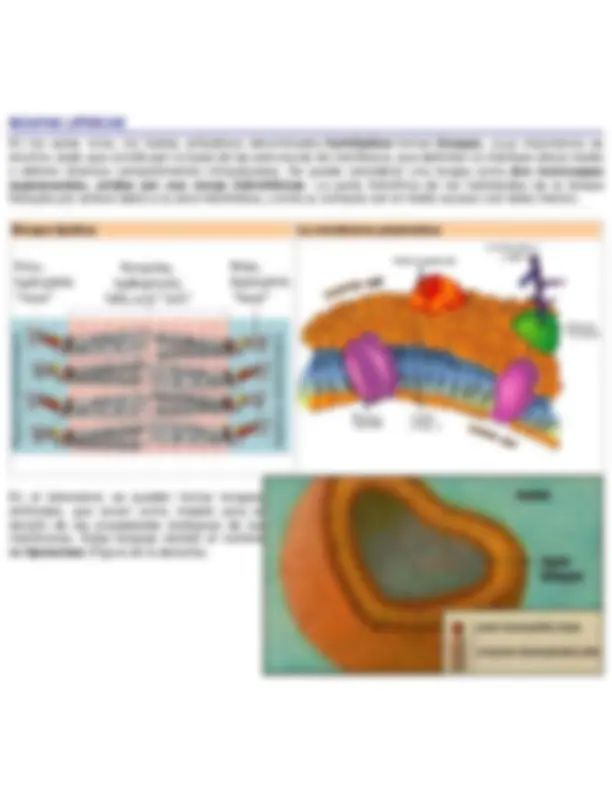
Una característica básica de los lípidos, y de la que derivan sus principales propiedades biológicas es la hidrofobicidad. La baja solubilidad de los lipídos se debe a que su estructura química es fundamentalmente hidrocarbonada (alifática, alicíclica o aromática), con gran cantidad de enlaces C-H y C-C (Figura de la izquierda). La naturaleza de estos enlaces es 100% covalente y su momento dipolar es mínimo. El agua, al ser una molécula muy polar, con gran facilidad para formar puentes de hidrógeno, no es capaz de interaccionar con estas moléculas. En presencia de moléculas lipídicas, el agua adopta en torno a ellas una estructura muy ordenada que maximiza las interacciones entre las propias moléculas de agua, forzando a la molécula hidrofóbica al interior de una estructura en forma de jaula, que también reduce la movilidad del lípido. Todo ello supone una configuración de baja entropía, que resulta energéticamente desfavorable. Esta disminución de entropía es mínima si las moléculas lipídicas se agregan entre sí, e interaccionan mediante fuerzas de corto alcance, como las fuerzas de Van der Waals. Este fenómeno recibe el nombre de efecto hidrofóbico (Figuras inferiores).
Dispersión de lípidos en medio acuoso Agregación de lípidos en medio acuoso
Los lípidos (generalmente en forma de triacilgiceroles) constituyen la reserva energética de uso tardío o diferido del organismo. Su contenido calórico es muy alto ( Kcal/gramo), y representan una forma compacta y anhidra de almacenamiento de energía.
A diferencia de los hidratos de carbono, que pueden metabolizarse en presencia o en ausencia de oxígeno, los lípidos sólo pueden metabolizarse aeróbicamente.
En algunos animales hay un tejido adiposo especializado que se llama grasa parda o grasa marrón. En este tejido, la combustión de los lípidos está desacoplada de la fosforilación oxidativa, por lo que no se produce ATP, y la mayor parte de la energía derivada de la combustión de los triacilgliceroles se destina a la producción de calor.
El medio biológico es un medio acuoso. Las células, a su vez, están rodeadas por otro medio acuoso. Por lo tanto, para poder delimitar bien el espacio celular, la interfase célula-medio debe ser necesariamente hidrofóbica. Esta interfase está formada por lípidos de tipo anfipático, que tienen una parte de la molécula de tipo hidrofóbico y otra parte de tipo hidrofílico. En medio acuoso, estos lípidos tienden a autoestructurarse formando la bicapa lipídica de la membrana plasmática que rodea la célula.
En las células eucariotas existen una serie de orgánulos celulares (núcleo, mitocondrias, cloroplastos, lisosomas, etc) que también están rodeados por una membrana constituída, principalmente por una bicapa lipídica compuesta por fosfolípidos. Las ceras son un tipo de lípidos neutros, cuya principal función es la de protección mecánica de las estructuras donde aparecen.
Los organismos pluricelulares han desarrollado distintos sistemas de comunicación entre sus órganos y tejidos. Así, el sistema endocrino genera señales químicas para la adaptación del organismo a circunstancias medioambientales diversas. Estas señales reciben el nombre de hormonas. Muchas de estas hormonas (esteroides, prostaglandinas, leucotrienos, calciferoles, etc) tienen estructura lipídica.
Funcionamiento de las hormonas esteroideas Algunos usos de las hormonas esteroideas
Estos enlaces te llevan a páginas con información sobre el uso de esteroides anabolizantes en la práctica deportiva: Enlace 1. Enlace 2.
En otros casos, los lípidos pueden funcionar como segundos mensajeros. Esto ocurre cuando se activan las fosfolipasas o las esfingomielinasas e hidrolizan glicerolípidos o esfingolípidos generando diversos compuestos que actúan como segundos mensajeros (diacilgliceroles, ceramidas, inositolfosfatos, etc) que intervienen en multitud de procesos celulares. (Ver figura inferior).
Los lípidos pueden funcionar como segundos mensajeros
La heterogeneidad estructural de los lípidos dificulta cualquier clasificación sistemática. El componente lipídico de una muestra biológica puede ser extraído con disolventes orgánicos y ser sometido a un criterio empírico : la reacción de saponificación.
La saponificación consiste en una hidrólisis alcalina de la preparación lipídica (con KOH o NaOH). Los lípidos derivados de ácidos grasos (ácidos monocarboxílicos de cadena larga) dan lugar a sales alcalinas (jabones) y alcohol, que son fácilmente extraíbles en medio acuoso. No todos los lípidos presentes en una muestra biológica dan lugar a este tipo de reacción. Se distinguen por tanto dos tipos de lípidos:
lípidos saponificables lípidos no saponificables
Los lípidos saponificables agrupan a los derivados por esterificación u otras modificaciones de ácidos grasos, y se sintetizan en los organismos a partir de la aposición sucesiva de unidades de dos átomos de carbono. En este grupo se incluyen:
ácidos grasos y sus derivados eicosanoides (prostaglandinas, tromboxanos y leucotrienos) lípidos neutros (acilgliceroles y ceras) lípidos anfipáticos (glicerolípidos y esfingolípidos).
Desde el punto de vista químico, son muy poco reactivos. Por lo general, contienen un número par de átomos de carbono. En la nomenclatura de los ácidos grasos se utilizan con más frecuencia los nombres triviales que los sistemáticos. La nomenclatura abreviada es muy útil para nombrar los ácidos grasos. Consiste en una C, seguida de dos números, separados por dos puntos. El primer número indica la longitud de la cadena hidrocarbonada, mientras que el segundo indica el número de dobles enlaces que contiene.
Los ácidos grasos saturados más abundantes son el palmítico (hexadecanoico, o C16:0) y el esteárico (octadecanoico, o C18:0). Los ácidos grasos saturados de menos de 10 átomos de C son líquidos a temperatura ambiente y parcialmente solubles en agua. A partir de 12 C, son sólidos y prácticamente insolubles en agua. En estado sólido, los ácidos grasos saturados adoptan la conformación alternada todo-anti, que da un máximo de simetría al cristal, por lo que los puntos de fusión son elevados. El punto de fusión aumenta con la longitud de la cadena.
ácido mirístico (C14:0)
ácido palmítico (C16:0)
ácido esteárico (C18:0)
ácido lignocérico (C24:0)
Los ácidos grasos de cadena impar probablemente derivan de la metilación de un ácido graso de cadena par. En ellos, la simetría del cristal no es tan perfecta, y los puntos de fusión son menores. Ejemplos son el ácido propiónico (C3:0), valeriánico (pentanoico, o C5:0) y pelargónico (nonanoico, o C9:0).
Los lípidos ricos en ácidos grasos saturados constituyen las grasas. Conviene en este punto hacer una distinción entre los términos lípidos, grasas y aceites. Grasas son aquellos lípidos que son sólidos a temperatura ambiente, mientras que aceites son aquellos lípidos que son líquidos a temperatura ambiente. Tanto los aceites como las grasas son lípidos.
Con mucha frecuencia, aparecen insaturaciones en los ácidos grasos, mayoritariamente en forma de dobles enlaces , aunque se han encontrado algunos con triples enlaces. Cuando hay varios dobles enlaces en la misma cadena, estos no aparecen conjugados (alternados), sino cada tres átomos de carbono. En la nomenclatura abreviada , se indica la longitud de la cadena y el número de dobles enlaces. La posición de los dobles enlaces se indica como un superíndice en el segundo múmero. Así, el ácido oleico (9-octadecenoico) se representa como C18:1^9 , y el linoleico (9,12-octadecadienoico) como C18:29,12, y el linolénico (9,12,15-octadecatrienoico) como C18:39,12,15.
ácido oleico (C18:1^9 ) ácido linoleico (C18:2 9,12 ) ácido linolénico (C18:3 9,12,15 )
Por lo general, las insaturaciones de los ácidos grasos son del tipo cis. Esto hace que la disposición de la molécula sea angulada, con el vértice en la insaturación. Esta angulación hace que los puntos de fusión de las ácidos insaturados sean más bajos que los de sus homólogos saturados. Los dobles
enlaces en trans distorsionan poco la simetría cristalina, que es muy parecida a la de los ácidos grasos saturados.
ácido esteárico (C18:0) ácido trans - oleico (C18:19t)
ácido cis - oleico (C18:19c)
La configuración en cis o en trans de un doble enlace en la cadena hidrocarbonada también puede indicarse en la nomenclatura abreviada. Así, el ácido araquidónico (5,8,11,14-eicosatetraenoico) se representa como C20:45c,8c,11c,14c^ ó C20:4 (5c, 8c, 11c, 14c).
Algunos ácidos grasos poliinsaturados (linoleico, linolénico y araquidónico) no pueden ser sintetizados por los animales superiores (incluído el hombre), y como su función biológica es fundamental, deben ser suministrados en la dieta. Por este motivo reciben el nombre de ácidos grasos esenciales.
Los ácidos grasos insaturados manifiestan las propiedades inherentes al doble enlace :
Con mucha menor frecuencia, aparecen en la Naturaleza ácidos grasos cuya estructura difiere en mayor o medida de la que hemos visto hasta ahora. Entre ellos podemos destacar:
Este término agrupa a una serie de compuestos derivados de ácidos grasos poliinsaturados de 20 átomos de carbono (de donde deriva su nombre), como el ácido araquidónico (Figura de la derecha).
Como la diversidad de los eicosanoides es grande, estos compuestos se clasifican en función de las enzimas que intervienen en su síntesis :
Si son productos de la ruta de la ciclooxigenasa : prostaglandinas y tromboxanos Si son productos de la ruta de la lipoxigenasa: leucotrienos
Tienen una amplia gama de actividades biológicas: intervienen en procesos alérgicos, inflamatorios, provocan la contracción del músculo liso (en la menstruación y en el parto). Son el prototipo de mediadores locales , liberados in situ ante diversos estímulos. Aunque son compuestos que funcionan como señales químicas, difieren de las hormonas en dos aspectos importantes :
Se sintetizan prácticamente en todos los tejidos , no en una glándula endocrina Químicamente son muy inestables y, por tanto, sólo actúan a nivel local
Tipos de eicosanoides:
PROSTAGLANDINAS
TROMBOXANOS
LEUCOTRIENOS
Son los primeros miembros conocidos del grupo. Las prostaglandinas (PG) se consideran derivados de un hipotético ácido prostanoico (no existe como tal en la naturaleza), de 20 átomos de C, con un anillo pentagonal entre los carbonos 8 y 12.
Existen varias familias de PG, que se denominan con una letra adicional (PGA, PGB, PGC, PGD, PGE, PGF, etc), en función de los sustituyentes del anillo ciclopentano de su estructura. A menudo, la letra mayúscula va seguida de un subíndice que indica el número de dobles enlaces presentes en la molécula, sin incluir el anillo.
Se conocen unas 20 PG, cuya función es la de regular la acción hormonal. Las PGE y PGF provocan la contracción de la musculatura lisa, en especial en el aparato reproductivo, de ahí que sean utilizadas para inducir el aborto.
Prostaglandina E 1 Prostaglandina E 2
Son derivados eicosanoides que deben su nombre a la presencia de tres dobles enlaces conjugados (Ver figura inferior). Se sintetizan a partir de la ruta de la lipoxigenasa, que es especialmente activa en los leucocitos.
Son mediadores locales que intervienen en reacciones de tipo alérgico, asmático o inflamatorio. Aparecen frecuentemente combinados con el tripéptido glutatión, como en el caso del leucotrieno C 4 (LTC 4 ).
Son ésteres de ácidos grasos con alcoholes. No tienen ningún otro tipo de componentes, por loque son moléculas muy poco reactivas.
En la Naturaleza encontramos dos tipos:
acilgliceroles
ceras
Los acilgliceroles o glicéridos son ésteres de ácidos grasos con glicerol (propanotriol). Constituyen el contingente mayoritario de los lípidos de reserva energética, y son muy abundantes en el tejido adiposo animal y en las semillas y frutos de las plantas oleaginosas.
El glicerol o propanotriol presenta tres grupos alcohólicos, y por tanto puede aparecer esterificado en una, dos o tres posiciones, dando lugar respectivamente, a monoacilgliceroles (monoglicéridos), diacilgliceroles (diglicéridos) y triacilgliceroles (triglicéridos). En su inmensa mayoría se presentan como triésteres, aunque los mono y diacilgliceroles aparecen esporádicamente como intermediarios en la biosíntesis o degradación de triglicéridos, o como segundos mensajeros hormonales. Los triglicéridos son moléculas muy hidrofóbicas, mientras que los mono y diacilgliceroles presentan carácter anfipático debido a los grupos OH no esterificados.
Diglicérido Triglicérido (2D) Triglicérido (3D)
Las tres posiciones de esterificación pueden aparecer con 3 ácidos grasos iguales, dos iguales y uno distinto, o los tres diferentes (figura de la izquierda). Por este motivo, el carbono del alcohol secundario del glicerol puede resultar asimétrico. En algunos casos (cuando los sustituyentes en C y C3 son iguales), es imposible distinguir entre el C1 y el C3 del glicerol, y por lo tanto se recurre a la numeración estereoquímica de los carbonos del glicerol , que se denota anteponiendo el prefijo sn- al número del carbono (figura de la derecha). Según esta numeración, si se sitúa el OH del alcohol secundario a la izquierda, se considera como carbono 1 al que queda arriba utilizando la proyección de Fischer. Para nombrarlos se indican los radicales de ácidos grasos, seguido de glicerol. Así, el
En presencia de agua , las colas hidrofóbicas tienden a interaccionar entre sí , creando un espacio hidrofóbico del que el agua es excluída y en el que pueden quedar atrapadas otras moléculas hidrofóbicas, mientras que la cabeza polar interacciona con el agua , y se encuentra solvatada, preservando a la parte hidrofóbica de todo contacto con el agua. Este es el llamado efecto hidrofóbico (Figura de la derecha).
El efecto hidrofóbico es el responsable de que en presencia de agua, los lípidos anfipáticos tengan la importante propiedad de la autoestructuración , que da lugar a tres tipos de estructuras distintas:
monocapas micelas bicapas
Las monocapas se forman en la interfase aire-agua. Las colas hidrofóbicas se orientan hacia el aire, mientras que las cabezas polares lo hacen hacia el agua (figura de la derecha). Las monocapas son susceptibles de compresión lateral mecánica (en el sentido de la flecha, en la figura inferior), de forma que las moléculas quedan totalmente empaquetadas en la interfase aire-agua.