¡Descarga El Núcleo Celular: Función, Estructura y Transporte de Moléculas - Prof. Parra y más Apuntes en PDF de Biología Celular solo en Docsity!
TEMA 5.
EL NÚCLEO CELULAR.
CONCEPTOS BÁSICOS:
♦ RNA de Transferencia (tRNA) - Constituye un 20% del total de RNA de la célula. Son moléculas relativamente pequeñas ya que constan de entre 75 y 95 nucleótidos. Pueden tener hasta un 10% de bases diferentes de las comunes.
■ Su función consiste en aportar aminoácidos durante la síntesis de las proteínas.
- Estructura tridimensional (terciaria)
- Cada molécula posee zonas de complementariedad (brazos) y otras no apareadas (bucles).
- Cada bucle tiene una función: unión al ribosoma; reconocimiento de las aminoacil tRNA sintetasas ; anticodon.
- Especificidad de los tRNA (anticodon) –
- El anticodon es una secuencia de tres nucleótidos que determina qué aminoácido se une al tRNA. El aminoácido correspondiente se une al único brazo que no tiene bucle y que se conoce como brazo aceptor del aminoácido. –
- Los tRNA activados, es decir, unidos a su aminoácido correspondiente, se denominan aminoacil-tRNA.
♦ RNA Mensajero (mRNA) - Forma alrededor del 5 % del total del RNA de la célula. Se forman en el núcleo por complementariedad a partir de un gen (transcripción). Llevan una copia del mensaje genético contenido en el DNA al citoplasma, donde se encuentran los ribosomas que lo emplearán como molde en el proceso de síntesis de proteínas (traducción). Son degradados rápidamente después de realizar su función.
♦ RNA Ribosómico (rRNA) - Constituye el 75 % del RNA celular. Tiene zonas plegadas en doble cadena. Su función es estructural, ya que se asocia a proteínas para constituir los ribosomas.
LA EXISTENCIA DEL NÚCLEO ES LA CARACTERÍSTICA PRINCIPAL que diferencia las células eucariotas de las células procariotas y en la que se encuentra el ADN. Por contener el genoma celular, el núcleo sirve de almacén de la información genética y como centro de control celular. La replicación del ADN, la transcripción y el procesamiento del ARN ocurren en el interior del núcleo, y sólo la última etapa de la expresión génica (traducción) tiene lugar en el citoplasma.
Debido a que la envuelta nuclear separa el genoma del citoplasma, la expresión génica está regulada por mecanismos exclusivos de los organismos eucariotas. Mientras que los ARNm procariotas son traducidos a la vez que ocurre la transcripción, los ARNm eucariotas sufren procesos postranscripcionales antes de ser
transportados desde el núcleo al citoplasma. La presencia de un núcleo, por tanto, permite que la expresión génica sea regulada por mecanismos postranscripcionales, como el splicing alternativo. Por tanto, la separación entre el genoma y el lugar de la traducción del ARNm desempeña un papel fundamental en la expresión génica en las células eucariotas.
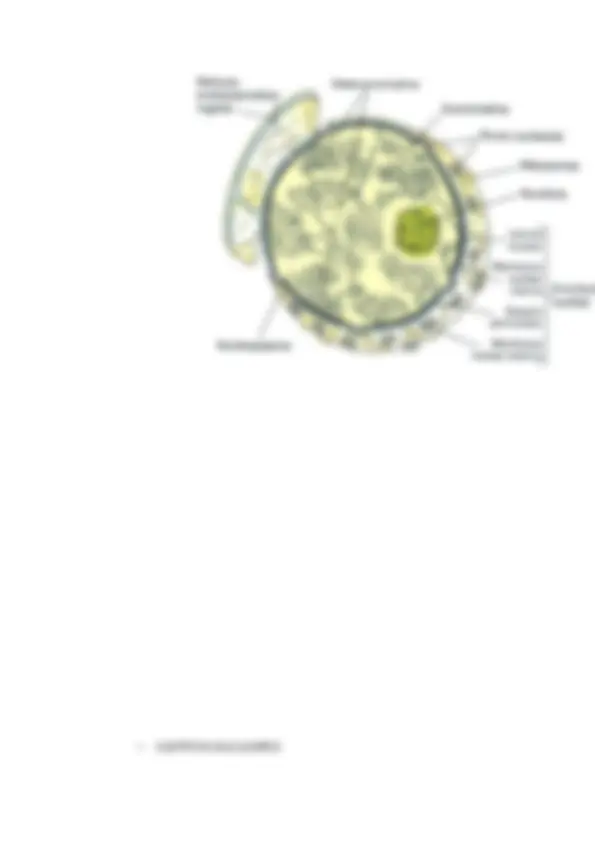
El núcleo celular tiene por lo tanto una envoltura doble. Las membranas nucleares actúan como una barrera selectiva que impide el libre paso de las moléculas entre el interior nuclear y el citoplasma, manteniéndolos como dos compartimentos metabólicamente independientes. La membrana externa es continua al RER. Es muy parecida al RER pero ésta es muy rica en proteínas que la unen al citoesqueleto. La membrana nuclear interna tiene proteínas únicas que son específicas para el núcleo, como aquellas que unen la matriz nuclear de láminas. Las membranas interna y externa se unen en los complejos de poro nuclear , siendo los únicos canales que permiten el paso de pequeñas moléculas polares y de macromoléculas a través de la envuelta nuclear.
▲ Poros nucleares : Los únicos canales en la envuelta nuclear están representados por los complejos de poro nucleares, que permiten un intercambio controlado de moléculas entre el núcleo y el citoplasma. El tráfico selectivo de proteínas y ARN a través de los complejos de poro nucleares no sólo mantiene la composición interna del núcleo sino que tiene un papel clave en la regulación de la expresión génica. El complejo del poro nuclear tiene un papel fundamental en la fisiología de todas las células eucariotas. Las moléculas de ARN que son sintetizadas en el núcleo deben ser exportadas de manera eficiente al citoplasma, donde intervienen en la síntesis de proteínas. Por otro lado, las proteínas necesarias para las funciones nucleares (p. ej., factores de transcripción) deben entrar en el núcleo procedentes de los lugares de síntesis en el citoplasma.
Nucleoporinas : proteínas del anillo interno
poro nuclear se controla por una proteína denominada Ran. Ran representa una de las diversas clases de proteínas de bajo peso molecular que se unen a GPT cuya conformación y actividad están reguladas por la unión y la hidrólisis de GTP.
•.1. El importe de proteínas a través del complejo del poro nuclear comienza cuando una importina específica se une a la señal de localización nuclear de una proteína transportadora en el citoplasma. Este complejo importina/proteína de transporte se une a continuación a proteínas presentes en los filamentos citoplásmicos del complejo del poro nuclear, y se produce el transporte mediante la unión secuencial a proteínas específicas del poro nuclear, localizadas cada vez más cerca de la cara nuclear del complejo del poro.
•.2. En la cara nuclear del complejo del poro, el complejo de proteínas de transporte/importina se separa por la unión de Ran/GTP. Esto produce un cambio conformacional en la importina, que desplaza a la proteína de transporte y la libera en el núcleo.
•.3. El complejo importina-Ran/GTP se exporta a continuación a través del complejo del poro nuclear. En el citoplasma el GTP es hidrolizado a GDP , a partir de RAN/GAP, que es un enzima con el que se encuentra la importina-Ran/GTP.
•.4. Esto libera a la importina de modo que puede unirse a una nueva proteína transportadora en el citoplasma y volver a ser transportada al interior nuclear por su propio receptor de importe (una proteína denominada NTF2 ), donde el Ran/GTP es regenerado. mejorar
•.5. Algunas proteínas permanecen en el interior del núcleo una vez transportadas desde el citoplasma, pero muchas otras viajan continuamente entre el núcleo y el citoplasma. Algunas de estas proteínas actúan como transportadores (carriers) de otras moléculas, como los ARN; otras coordinan las funciones nucleares y citoplasmáticas (p. ej., regulando la actividad de los factores de transcripción).
•.6. Las proteínas se etiquetan para ser exportadas del núcleo mediante una secuencia de aminoácidos específica, llamada señal de exportación nuclear. Las señales de exportación nuclear son reconocidas por receptores en el interior del núcleo, exportinas , que dirigen el transporte de las proteínas a través del complejo del poro nuclear al citoplasma. Al igual que las importinas, muchas exportinas pertenecen a una familia de receptores de transporte nuclear denominados carioferinas. Las exportinas se unen a Ran, necesaria tanto para la exportación nuclear como la importación nuclear. Sorprendentemente, sin embargo Ran/GTP promueve la formación de complejos estables entre las exportinas y las proteínas diana, mientras que disocia los complejos entre las
importinas y sus proteínas diana. Este efecto de la unión de Ran/ GTP sobre las exportinas dirige el movimiento de las proteínas con señales de exportación nuclear desde el núcleo al citoplasma. Por tanto, las exportinas forman complejos estables con sus proteínas diana y con Ran/GTP en el interior del núcleo. Una vez que se ha producido el transporte al lado citosólico de la envuelta nuclear, la hidrólisis de GTP y la liberación de Ran/GDP provoca la disociación de la proteína diana, que es liberada en el citoplasma, se ha separado todo por la acción de RAN/GAP. Las exportinas se reciclan a través del complejo del poro nuclear para su reutilización.
La mayoría de los ARN son exportados desde el núcleo al citoplasma. Puesto que las proteínas se sintetizan en el citoplasma, la salida de los ARNm, ARNr, ARNt y microARN (ARNmi) es un proceso fundamental en la expresión génica en las células eucariotas. La salida de los ARN a través de los complejos del poro nuclear es un proceso activo , dependiente de energía, requiriendo que los receptores de transporte interaccionen con el complejo del poro nuclear. Los ARNm son exportados por un complejo de dos proteínas y este transporte de ARNm parece ser independiente de Ran. Los ARN son transportados a través de la envuelta nuclear como complejos ribonucleoproteína (RNP). Los ARN ribosómicos se asocian en primer lugar con proteínas ribosómicas y con proteínas específicas del procesamiento de ARN en el nucléolo, y las subunidades 60S y 40S son entonces transportadas de manera independiente al citoplasma a través de un mecanismo en el que interviene la carioferina. Su exportación del núcleo está mediada por señales de exporte nuclear presentes en las proteínas del complejo de la subunidad. Los pre-ARNm y ARNm están asociados con un conjunto de al menos 20 proteínas durante su procesamiento en el núcleo y posterior transporte al citoplasma, lo que está mediado por el complejo exportador del ARNm, que tendrá que desenrollar el ARN para que pueda ser exportado. Los ARNt precursores de ARNmi son exportados del núcleo por la acción de la exportina-t, y exportina5, respectivamente (las exportinas reconocen cuando tiene que ser exportado ese ARN) que se une directamente a los ARNt.
- (^) ORGANIZACIÓN INTERNA DEL NÚCLEO INTERFÁSICO
- Nucleoplasma : composición parecida al citoplasma: agua, sales minerales...
Nucleoesqueleto de láminas : Subyacente a la membrana nuclear interna se localiza la lámina nuclear, una red fibrosa que proporciona soporte estructural al núcleo. La lámina nuclear está compuesta de proteínas fibrosas denominadas lamininas junto a algunas proteínas asociadas. Las láminas son las encargadas de dar forma al núcleo. Cuando se fosforilan pierden su estructura y como consecuencia se desorganiza la envoltura del núcleo. Esto ocurre
Por tanto, la cantidad de heterocromatina facultativa varía dependiendo de la actividad transcripcional de la célula.
Modificaciones de la heterocromatina: cuando las histonas están en estado de acetilacion, la polimerasa será capaz de transcribir esos genes. Sin embargo, si la histona está metilada, no se podrá transcribir los genes.
Eucromatina : la que se transcribe a menudo es acetilada.
Epigenética : hay caracteres heredables que no están en el adn.tienen que ver con cómo estan las histonas de una determinada región (metiladas, acetiladas).
▲ Superficie de la cromatina
■ Factorías de transcripción
Lugares donde se acumulan las enzimas que permiten que se transcriban esos genes, donde están las polimerasas. También está el ARN siendo transcrito y proteínas que se asocian a ese ARN transcrito.
▲ Extremos del cromosoma : telómeros, tienen una región especial que hace un bucle y se sella.
▲ Centrómero , se localizan muchas proteínas que hacen posible que el huso se una a ellas.
▲ Cromosoma en interfase : solo hay una cromátida.
• CUERPOS NUCLEARES
La cromatina correspondiente a cada uno de los cromosomas tiende a ocupar unas posiciones concretas en el núcleo en interfase , y que esas posiciones tienden a mantenerse durante el ciclo celular.
En determinadas situaciones celulares los cromosomas se encuentran cerca de la envoltura del núcleo (no se están transcribiendo los genes) (heterocromatina), cuando los cromosomas se encuentran más en el interior del núcleo es porque se están transcribiendo, esto quiere decir que se encuentran cerca de las factorías de transcripción, a la hora de transcribirse.