¡Descarga Envolturas celulares y más Apuntes en PDF de Biología solo en Docsity!
TEMA 7: ENVOLTURAS CELULARES
1. LA MEMBRANA PLASMÁTICA
a. Composición b. Estructura (mosaico fluido) c. Funciones
- TRANSPORTE A TRAVÉS DE LA MEMBRANA a. Sin deformación de la membrana i. Transporte pasivo 1. Difusión simple 2. Difusión facilitada ii. Transporte activo (bombas) b. Con deformación de la membrana i. Endocitosis 1. Fagocitosis 2. Pinocitosis ii. Exocitosis iii. Transcitosis
- ESTRUCTURAS DE UNIÓN ENTRE CÉLULAS a. Adhesiones mecánicas i. Bandas de adhesión ii. Desmosomas iii. Hemidesmosomas b. Uniones estrechas c. Uniones tipo GAP d. Plasmodesmos
- LA MATRIZ EXTRACELULAR
- LA PARED CELULAR a. Vegetal b. Hongos c. Moneras
1.- LA MEMBRANA PLASMÁTICA
La membrana plasmática fue definida en 1967 por Palade como un complejo molecular que delimita un territorio celular determinado. Durante mucho tiempo sólo se pudo intuir su existencia ya que es una estructura tan delgada que escapaba a la observación mediante el microscopio óptico. Hoy sabemos que la membrana plasmática es una envoltura continua que rodea la célula estando una de sus caras en contacto con el medio extracelular y la otra con el citoplasma. La observación al microscopio electrónico revela una estructura de unos 7 nm de grosor en la que se aprecian dos bandas oscuras separadas por una banda más clara. COMPOSICIÓN La casi totalidad de la masa de la membrana plasmática está constituida por proteínas y lípidos anfipáticos ; contiene además pequeñas cantidades de glúcidos en forma de oligosacáridos unidos covalentemente a las proteínas o a los lípidos (glicoproteínas y glucolípidos). A veces aparecen en gran cantidad en la cara externa de la membrana formando el glucocálix. En la mayor parte de los casos la masa total de proteínas supera a la de lípidos. Las membranas que constituyen el sistema membranario interno característico de la célula eucariota presentan una composición y estructura muy similares a las de la membrana plasmática. LOS LÍPIDOS que aparecen formando parte de las membranas biológicas son fosfoglicéridos, esfingolípidos y colesterol (u otros esteroles afines). Todos ellos tienen en común su carácter marcadamente anfipático que los hace idóneos para este cometido. Las proporciones relativas de los distintos tipos de lípidos varían según el tipo de célula; las membranas presentes en cada reino, especie, tejido y tipo celular poseen una composición lipídica característica, e incluso, dentro de un mismo tipo celular esta composición puede variar entre los diferentes orgánulos. LAS PROTEÍNAS de membranas biológicas de orígenes diferentes varía aún más ampliamente que su composición lipídica, lo que refleja que estas proteínas deben estar especializadas en determinadas funciones que son diferentes según el tipo de célula. Como regla general, una membrana biológica posee varios centenares de proteínas diferentes, la mayoría de las cuales están especializadas en el transporte de solutos específicos a su través.
átomos más ligeros y por lo tanto más transparentes a los electrones). Por otra parte, un grosor de entre 5 y 8 nm es el que cabría esperar de una bicapa de lípidos anfipáticos con proteínas incrustadas en ella y sobresaliendo a ambos lados. Los experimentos llevados a cabo con diferentes tipos de lípidos anfipáticos indican que éstos en medio acuoso tienden espontáneamente a formar estructuras como las micelas, bicapas y liposomas, ya estudiadas anteriormente, lo que también apoya la idea de una bicapa lipídica como elemento básico estructural en las membranas celulares. El carácter fluido de las membranas se estableció mediante el marcaje de moléculas individuales de lípidos y su posterior seguimiento en el seno de la bicapa. Así pudo comprobarse que los movimientos de los lípidos dentro de cada monocapa son muy frecuentes y rápidos (difusión lateral y rotación), mientras que son mucho más raros los cambios de una a otra monocapa ("flip-flop"). El grado de fluidez de las membranas depende de la temperatura y de la composición en ácidos grasos de sus lípidos constituyentes. A temperaturas muy bajas los lípidos de membrana tienden a adoptar un ordenamiento casi cristalino (paracristalino). Por encima de una temperatura que es característica de cada membrana las moléculas de los lípidos comienzan a moverse y la membrana pasa al estado fluido. La temperatura de transición del estado paracristalino al estado fluido depende de la composición lipídica; los ácidos grasos saturados favorecen un mayor empaquetamiento de los lípidos de membrana y por lo tanto la temperatura de transición será mayor cuanto mayor sea la proporción de éstos ácidos grasos en la membrana. Por el contrario, los cambios de orientación existentes en las cadenas de los ácidos grasos insaturados dificultan el empaquetamiento de los lípidos de membrana en un ordenamiento paracristalino, por lo tanto la temperatura de transición será menor cuanto más abundante sea este tipo de ácidos grasos. Esta es la razón por la que en los organismos homeotermos, que mantienen una temperatura corporal constante y elevada, abundan los lípidos de membrana ricos en ácidos grasos saturados, mientras que en los organismos poiquilotermos, incapaces de regular su temperatura corporal, son más abundantes los lípidos de membrana ricos en ácidos grasos insaturados. Se ha comprobado que distintos tipos de células mantenidos en cultivo son capaces de alterar la composición en ácidos grasos de sus lípidos de membrana respondiendo a las variaciones de la temperatura ambiente, todo ello con el objeto de mantener la fluidez de sus membranas celulares. Por otra parte, el colesterol y esteroles afines, debido a la rigidez del sistema de anillos condensados de su
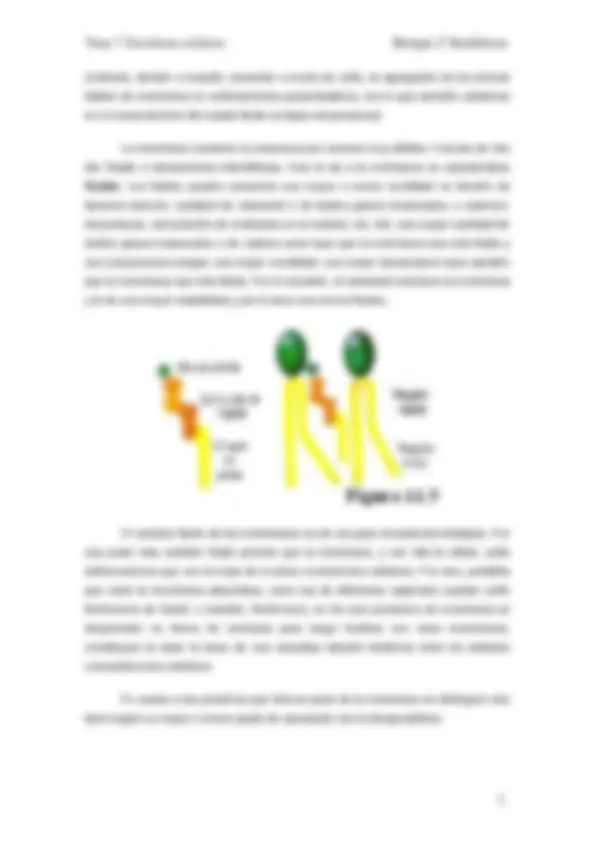
molécula, tienden a impedir, actuando a modo de cuña, la agregación de los demás lípidos de membrana en ordenamientos paracristalinos, con lo que también colaboran en el mantenimiento del estado fluido (a bajas temperaturas) La membrana mantiene su estructura por uniones muy débiles: Fuerzas de Van der Waals e interacciones hidrofóbicas. Esto le da a la membrana su característica fluidez. Los lípidos pueden presentar una mayor o menor movilidad en función de factores internos: cantidad de colesterol o de ácidos grasos insaturados, o externos: temperatura, composición de moléculas en el exterior, etc. Así, una mayor cantidad de ácidos grasos insaturados o de cadena corta hace que la membrana sea más fluida y sus componentes tengan una mayor movilidad; una mayor temperatura hace también que la membrana sea más fluida. Por el contrario, el colesterol endurece la membrana y le da una mayor estabilidad y por lo tanto una menor fluidez. El carácter fluido de las membranas es de una gran importancia biológica. Por una parte este carácter fluido permite que la membrana, y con ella la célula, sufra deformaciones que son la base de muchos movimientos celulares. Por otra, posibilita que tanto la membrana plasmática, como las de diferentes orgánulos puedan sufrir fenómenos de fusión y escisión, fenómenos, en los que porciones de membrana se desprenden en forma de vesículas para luego fundirse con otras membranas, constituyen la base la base de una compleja relación dinámica entre los distintos compartimentos celulares. En cuanto a las proteínas que forman parte de la membrana se distinguen dos tipos según su mayor o menor grado de asociación con la bicapa lipídica:
paso de sustancias, no de todas, pero sí de muchas, tanto del exterior al interior como en sentido inverso. No obstante, y a pesar de esta función limitante, la célula va a necesitar intercambios constantes con el medio que la rodea. Necesita sustancias nutritivas y tiene que eliminar productos de desecho, que serán transportados a través de la membrana y por la propia membrana. La membrana es un elemento activo que "escoge" lo que entrará o saldrá de la célula.
RECEPTORA. Algunas proteínas de la membrana plasmática van a tener esta
función, por ejemplo: receptoras de sustancias hormonales. Muchas hormonas regulan la actividad de la célula fijándose en determinados puntos de proteínas receptoras específicas. La proteína receptora va a liberar en el interior de la célula una molécula orgánica: el mediador hormonal. Esta sustancia va a actuar regulando ciertos aspectos del metabolismo celular, por ejemplo, activando determinadas enzimas o desencadenando la activación de determinados genes. Al existir diferentes proteínas receptoras en la membrana celular y al tener las células diferentes receptores, la actividad de cada célula será diferente según sean las hormonas presentes en el medio celular.
RECONOCIMIENTO. Se debe a las glicoproteínas de la cara externa de la
membrana. Así, las células del sistema inmunológico, células que nos defienden de los agentes patógenos, van a reconocer las células que son del propio organismo diferenciándolas de las extrañas a él por las glicoproteínas de la membrana. Estas sustancias constituyen un verdadero código de identidad. 2.- TRANSPORTE A TRAVÉS DE LA MEMBRANA La composición y estructura de las membranas celulares, que acabamos de estudiar, hacen que éstas, lejos de constituir meras barreras pasivas, participen de una manera activa en la regulación del tráfico de sustancias entre la célula y su entorno y entre los distintos compartimentos celulares. TRANSPORTE SIN DEFORMACIÓN DE LA MEMBRANA Las moléculas pequeñas o los iones pueden atravesar la membrana sin necesidad de que ésta sufra alteraciones en su estructura. Este proceso puede ser un transporte pasivo (sin consumo de energía) o activo (requiere energía).
1. TRANSPORTE PASIVO: Se produce a favor de gradiente de concentración. Las moléculas atravesarán la membrana de la parte más concentrada a la menos concentrada. a. Difusión simple: Cualquier molécula bien hidrófoba, bien pequeña, o bien pequeña y sin carga puede atravesar la membrana. Pasan así sustancias lipídicas como las hormonas esteroideas, los fármacos liposolubles y los anestésicos, como el éter. También sustancias apolares como el O 2 y el N 2 atmosférico y algunas moléculas polares muy pequeñas como el agua, el CO 2 , el etanol y la glicerina. b. Difusión facilitada: Las moléculas grandes o los iones necesitan la ayuda de proteínas transmembrana. i. Proteínas transportadoras, permeasas o Carriers : Transportan glúcidos, aminoácidos y nucleósidos. Se unen en un lado de la membrana a la molécula que deben transportar, sufren un cambio conformacional y liberan la molécula al otro lado. Se trata de un transporte selectivo, ya que los centros de unión de cada transportador son específicos para una molécula concreta. ii. Proteínas canal: Forman un canal a través de la membrana. Pasan así ciertos iones, como el Na+, el K+^ y el Ca2+. Es un mecanismo rápido pero menos selectivo que el anterior. 2. TRANSPORTE ACTIVO: Transporte en contra del gradiente de concentración. Las células lo utilizan para mantener su composición. Requiere energía que es proporcionada por una reacción acoplada (hidrólisis de ATP). Lo realizan proteínas transmembrana, gracias a cambios conformacionales controlados por la hidrólisis de ATP, que pasan sustancias del lado menos concentrado al más concentrado. Este tipo de proteínas reciben el nombre de “bombas”. Las más conocidas son la bomba de glucosa (impulsada por 2Na+), la bomba de Na+/K+, la bomba de Ca2+, y la bomba de H+. Las bombas iónicas mantienen el gradiente iónico a través de la membrana plasmática (potencial de membrana).
3.- ESTRUCTURAS DE UNIÓN ENTRE CÉLULAS
Las membranas de algunas células sufren modificaciones específicas. Tal es el caso de las células del epitelio intestinal que presentan numerosas microvellosidades encaminadas a aumentar la superficie de absorción. En otros casos aparecen estructuras de unión entre células encaminadas a estabilizar la formación y funcionamiento de los tejidos. Entre ellas destacaremos las adhesiones mecánicas , las uniones estrechas , las uniones tipo GAP y los plasmodesmos. 1) Adhesiones mecánicas : Se producen en tejidos sometidos a fuertes tensiones, como el epitelial o el muscular cardíaco. Proporcionan fuerza mecánica a la unión. a) Bandas de adhesión: Se dan cerca de los extremos apicales de células epiteliales. Unas proteínas transmembrana, las cadherinas , unen ambas células. En el interior celular las cadherinas se unen a haces de filamentos de actina (citoesqueleto) mediante otro tipo de proteínas llamadas cateninas. b) Desmosomas: Son uniones puntuales que forman como botones a distintos niveles entre las membranas plasmáticas de células contiguas. La unión se produce gracias a proteínas transmembrana tipo cadherinas que se anclan, en la cara interna de las membranas a una especie de disco denominado placa desmosómica. A esta placa se unen los filamentos intermedios del citoesqueleto. c) Hemidesmosomas : Tipo de unión que se produce entre la zona basal de las células epiteliales y la matriz extracelular sobre la que se asientan. Se asemejan a medio desmosoma, en el que los filamentos intermedios se unen a la placa con proteínas transmembrana (integrinas) que a su vez enlazan con el tejido conectivo subyacente. 2) Uniones estrechas : Sellan células adyacentes en los epitelios y actúan como barreras al paso de moléculas o iones a través de las superficies laterales de éstas. Están formadas por una red de hebras de proteínas transmembrana ( claudinas y ocludinas ) que se distribuyen a lo largo de todo el perímetro de la célula y que se unen a otras iguales en las células vecinas (similar al plástico que une un pack de latas de refresco). Suelen aparecer asociadas a otros tipos de uniones formando los **complejos de unión.
- Uniones tipo GAP (hendidura o nexus):** Formadas por proteínas transmembrana denominadas conexinas que se reúnen en grupos de seis formando estructuras hexagonales llamadas conexones. Los conexones forman un canal acuoso entre las células que permiten el paso de iones inorgánicos y pequeñas moléculas solubles en agua. Acoplamiento eléctrico: Las uniones en hendidura son
abundantes en el músculo cardíaco y liso. La despolarización de un grupo de células musculares rápidamente se extiende a las células adyacentes, causando contracciones coordinadas de todo el músculo. Acoplamiento metabólico: Muchas hormonas actúan elevando las concentraciones intracelulares de AMP cíclico, que inicia una vía de señalización dentro de la célula. El AMP cíclico rápidamente pasa a través de las uniones en hendidura y así, el estímulo hormonal en una célula puede propagarse a un grupo de células. 4) Plasmodesmos: Canales de comunicación entre células vegetales. Función equivalente a las uniones tipo GAP (intercambio de iones y moléculas pequeñas). La membrana plasmática de una célula es continua con la de su vecina y las paredes celulares, unidas por una sustancia cementante, quedan interrumpidas en estos puntos. Desmosoma Unión tipo GAP
Estos componentes proceden de la actividad secretora de la célula y se disponen en capas sucesivas que se van depositando desde fuera hacia dentro, de manera que las más recientes son las que quedan en contacto con la membrana plasmática. La primera capa en depositarse es la llamada lámina media , constituida exclusivamente por cemento (pectina). Su función es unir y cementar células adyacentes. A continuación, entre la lámina media y la membrana plasmática, se depositan las tres capas de la pared primaria , con moléculas de celulosa dispuestas en retículo y abundancia de cemento. Tanto la lámina media como la pared primaria pueden crecer en extensión a medida que crece la célula. Cuando la célula deja de crecer, pueden depositarse, aunque no siempre sucede, un número variable de capas que conforman la pared secundaria , con moléculas de celulosa muy abundantes dispuestas en haces paralelos y una cantidad menor de cemento. La pared secundaria queda depositada entre la pared primaria y la membrana plasmática. Una vez depositada la pared secundaria la célula ya no puede seguir creciendo. La pared celular vegetal, dada su gran rigidez, cumple una doble función de naturaleza estructural. Por una parte, constituye un verdadero exoesqueleto que protege a la célula impidiendo que sea deformada o dañada por esfuerzos mecánicos procedentes del exterior. Por otra, dado que el medio en que viven las células vegetales suele ser hipotónico con respecto a su interior, la pared celular protege a dichas células frente a fenómenos osmóticos desfavorables, impidiendo que penetre un exceso de agua por ósmosis que podría provocar la lisis celular por rotura de la membrana plasmática, la cual, por ser mucho menos resistente, no soportaría la presión hidrostática. Además de desempeñar estas funciones con carácter general, la pared celular puede sufrir modificaciones que conllevan una adaptación a funciones más específicas. La impregnación en lignina, ( lignificación ) o en sales minerales ( mineralización ) comunican a la pared una mayor resistencia mecánica sin que por ello pierda su permeabilidad; esto es lo que ocurre en tejidos conductores o de sostén que deben soportar grandes esfuerzos mecánicos. Por otra parte, determinados tejidos de la planta deben ser impermeables para evitar la desecación por exceso de transpiración en ambientes muy secos; esto se consigue mediante la impregnación de las paredes celulares de dichos tejidos en sustancias como la cutina ( cutinización ) o suberina ( suberificación ). En muchos casos las paredes celulares presenten diferenciaciones que facilitan la comunicación entre células vecinas. Las punteaduras son zonas delgadas de la pared, formadas sólo por lámina media y una pared primaria muy fina, que
permiten el paso a su través de agua y todo tipo de sustancias disueltas. Los plasmodesmos son conductos citoplasmáticos muy finos que comunican células vecinas; la membrana de ambas células se continúa a través del plasmodesmo. MONERAS Adosada a la membrana plasmática por su cara externa se encuentra una pared celular que no es de naturaleza celulósica como la de la célula vegetal sino que está formada por una macromolécula compleja denominada peptidoglucano. El peptidoglucano es una macromolécula gigante que a modo de saco rodea a toda la célula; está formado por cadenas paralelas de un heteropolisacárido complejo unidas transversalmente por cadenas de aminoácidos. Tal entramado molecular confiere una extraordinaria resistencia mecánica a la pared celular, que constituye un auténtico exoesqueleto cuya misión es proteger a la célula procariota frente a fenómenos osmóticos desfavorables a la par que mantener su forma y rigidez características. Algunas especies bacterianas presentan una envoltura lipídica adicional que rodea exteriormente a la pared celular (Gram-). El esqueleto de la pared celular bacteriana está constituido por un heteropolímero, el peptidoglucano mureína. El mismo, y las enzimas que intervienen en su síntesis, son una característica general de todas las eubacterias. Las arqueobacterias no poseen mureína. Esta macromolécula esta formada por una secuencia alternante de N-acetil- glucosamina (NAG) y el ácido N-acetilmurámico (NAM) unidos mediante enlaces ß- 1,4. La cadena es recta y no ramificada, constituyendo la estructura básica de la pared celular (su " backbone "). El ácido N-acetilmurámico es un éter resultante de la unión del oxhidrilo del C3 de la molécula de N-acetil-glucosamina con el oxhidrilo del ácido láctico. El grupo ácido del láctico enlaza con una pequeña cadena peptídica. Entre los aminoácidos típicos de esta cadena se encuentran la L-alanina, ácido D-glutámico, ácido m-diaminopimélico o la L-lisina o D-alanina.
osmóticos. Además, la pared celular permite la interacción con el medio externo ya que algunas de sus proteínas son adhesinas y receptores. Algunos de sus componentes tienen una alta capacidad inmunogénica. La pared celular es una estructura característica de los hongos y está compuesta por glucanos, quitina y glicoproteínas. Al no estar presentes los componentes de la pared celular fúngica en el ser humano, esta estructura es una diana excelente para la terapia antifúngica. Los polisacáridos más importantes son la quitina (polímero de n-acetil glucosamina), el manano (polímero de manosa) y el glucano (polímero de glucosa). ENLACES http://recursos.cnice.mec.es/biosfera/alumno/2bachillerato/La_celula/contenidos5.htm http://www.bionova.org.es/biocast/tema11.htm http://www.iespando.com/web/departamentos/biogeo/2BCH/PDFs/07Membranas.pdf http://www.genomasur.com/lecturas/Guia06.htm http://www.biologia.edu.ar/bacterias/micro4.htm