¡Descarga FISIOLOGIA MÉDICA II y más Apuntes en PDF de Biología solo en Docsity!
TEMA 17. APRENDIZAJE Y MEMORIA: CONCEPTOS, CATEGORÍAS
Y SISTEMAS NEURALES
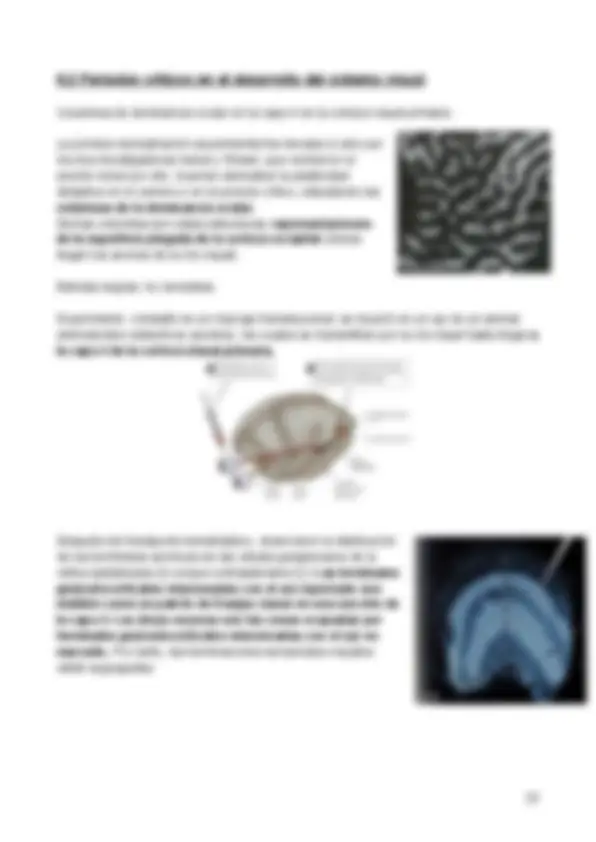
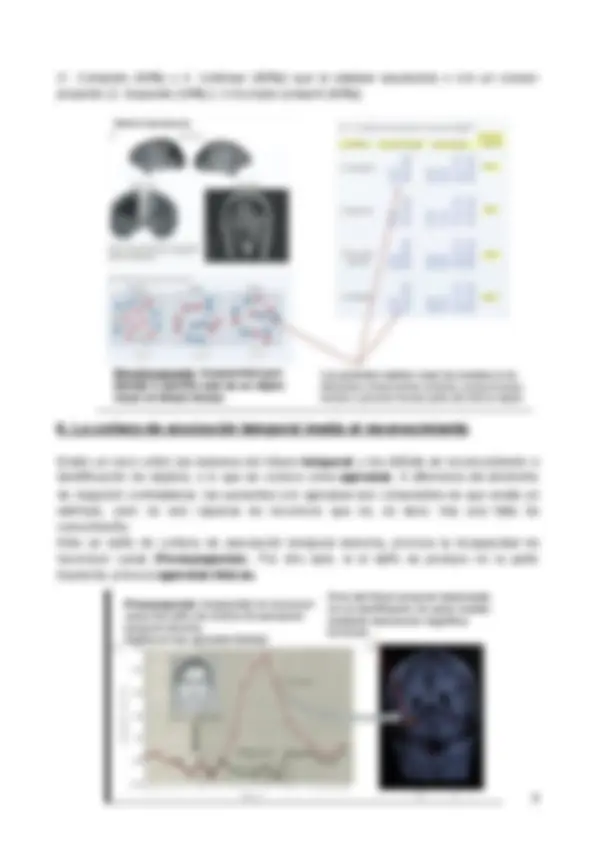
El “brainbow” es una técnica en la que se utiliza un único marcador fluorescente sobre un grupo de neuronas y donde, de manera aleatoria, éstas tomarán distintos colores.
En la imagen se observa un pliegue que se trata del hipocampo (en el lóbulo temporal), esencial para la memoria. “Hipocampo” significa caballito de mar (se trata de nombre científico de la especie) y este nombre se le atribuye debido a que el hipocampo completo de un ratón se asimila a la silueta de un caballito de mar.
1. Aprendizaje y memoria
El hipocampo , que se encuentra en el lóbulo temporal medial , es una estructura clave para la memoria declarativa , también llamada memoria explícita (es decir, la memoria de hechos y eventos que podemos expresar conscientemente). Si hay daño en el hipocampo o en estructuras cercanas del lóbulo temporal medial , puede haber una pérdida significativa de memoria declarativa , como ocurre en algunos tipos de amnesia.
La habilidad de almacenar información a través de la experiencia y la capacidad de recordarla tanto de forma consciente como inconsciente es una de las funciones complejas más fascinantes del cerebro debido a que el aprendizaje es esencial para la vida de una persona.
En función de la experiencia y la capacidad que tenemos de recoger, almacenar y olvidar información, nuestra capacidad de vivir es mayor o menor (como es el caso de pacientes amnésicos).
El aprendizaje de la memoria es la modificación de las conexiones sinápticas entre neuronas. Puesto que esta eficacia sináptica se puede modificar, si ésta aumenta, se mantendrá la información almacenada durante más tiempo, mientras que si se pierde, ocurre el olvido. Esto compone la memoria a nivel celular.
● Memoria declarativa: consiste en el tipo de memoria disponible en la conciencia, por lo que es íntegramente dependiente del lóbulo temporal cerebral. De ella dependen las palabras y sus significados, recuerdos de la historia o las ocurrencias diarias.
● Memoria no declarativa o procedimental: No depende del lóbulo temporal debido a que normalmente no está disponible en la conciencia. Su funcionamiento consiste en la relación entre estímulos similares previos y recientes y de ella dependen las actividades motoras, la asociación, la preparación o la resolución de problemas.
Clasificación de la memoria por el tiempo que es efectiva: ● Memoria inmediata: consiste en la retención de información durante unos segundos, como sería una cifra de 4 números que leemos en la calculadora hasta que la escribimos en el papel.
● Memoria a corto plazo / memoria trabajadora: consiste en la memoria que dura entre unos segundos hasta unos minutos, como sería recordar dónde hemos dejado algo o una palabra específica.
● Memoria a largo plazo: esta memoria dura días, años o incluso toda la vida y es el caso de recuerdos como dónde estábamos cada uno de nosotros el día que se declaró la pandemia del Covid-19.
Término idiot savant utilizado por primera vez para describir esta condición por el médico británico John Langdon Down, conocido por su descripción del síndrome de Down. Los síndrome de Down suelen desarrollar Alzheimer con 30/40 años.
1.2.Consolidación de memoria: importancia de la asociación
1.2.1. Condicionamiento clásico
Es el caso del experimento de los perros de Pavlov. Consiste en la generación de una nueva respuesta a un estímulo originalmente neutro, como sería el sonido de una campana en este caso, que se induce al asociar de forma repetitiva el estímulo neutro con otro estímulo que produce una respuesta, como es la comida. Es decir, el experimento de los perros de Pavlov consiste en que Pavlov hacía sonar una campanilla antes de darles de comer a los perros, por lo que con el tiempo, los perros al escuchar la campanilla comenzaban a salivar, ya que comenzaron a asociar el sonido de la campanilla con comida.
Este tipo de condicionamiento afecta a la memoria no declarativa o procedimental.
1.2.2. Condicionamiento operante
El condicionamiento operante consiste en la modificación de la probabilidad en la generación de una respuesta comportamental al asociarla a una recompensa o a un castigo. Fue investigación de Skinner. Es decir, se trata de un cambio en el comportamiento según esperemos recibir una recompensa o un castigo. Por ejemplo, un niño sonríe de forma accidental y cuando la madre la coge en brazos (recompensa para el niño), el niño relaciona sonreír con que lo cojan en brazos, por lo que cuando se acerque la madre sonreirá para que ésta la coja en brazos. El condicionamiento clásico se aprende y el operante ocurre de forma espontánea.
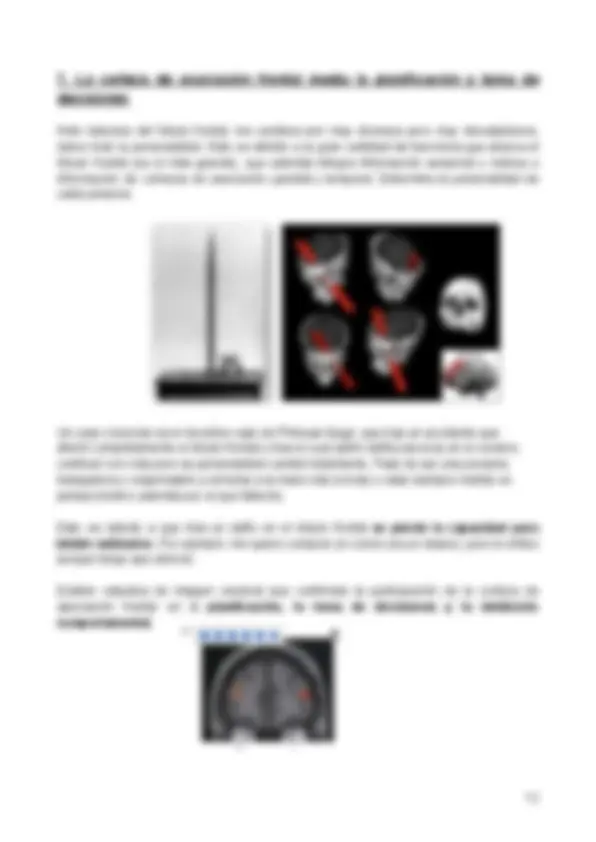
1.3. Sistemas cerebrales implicados en la memoria declarativa
Mediante el estudio de lesiones cerebrales en pacientes amnésicos, se observó la importancia del lóbulo temporal, más concretamente del hipocampo a la hora de la memoria. Estos estudios están corroborados en animales mediante la generación de lesiones en el lóbulo temporal de estos.
En pacientes con amnesia se puede ver que daños en el lóbulo temporal, especialmente la parte anterior, producen amnesia, por una relación entre el daño en el hipocampo y la pérdida de la memoria.
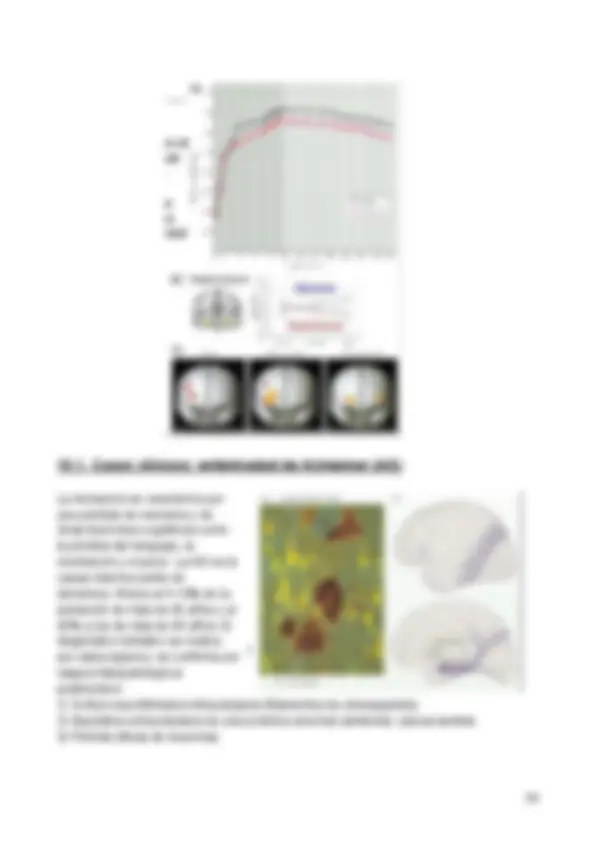
En cuanto a estudios en animales que corroboran los resultados podemos encontrar el siguiente: Se trata de un experimento con una balsa y un agua no transparente para que se observe. Se deja a la rata en la balsa para que nade y busque la plataforma (sumergida a ras del agua y no a la vista). Después de repetir el experimento 10 veces, la rata sabe dónde se encuentra la balsa. Esto ocurre porque el cuerpo tiene un sistema de posicionamiento , es decir, el gps del cerebro. Sin embargo, si a la rata le dañamos el hipocampo, nunca es capaz de encontrar la balsa porque el posicionamiento del animal en el espacio deja de funcionar. Esto muestra la importancia del lóbulo temporal y en particular del hipocampo en modelos animales.
1.4. Premios Nobel de fisiología
John O’Keefe: descubrió en 1971 que ciertas células nerviosas del cerebro se activaban cuando una rata se encontraba en un lugar específico en un lugar. Otras células nerviosas se activaban en otros lugares. Propuso que estas “células de lugar” constituyen un mapa interno de los alrededores. Estas células de lugar están localizadas en el hipocampo.
May-Britt Moser y Edvard Moser: descubrieron en 2005 que otras células nerviosas en la parte del cerebro de la corteza entorrinal se activan cuando la rata pasa por ciertas localizaciones. Juntas, estas localizaciones forman una red hexagonal donde cada célula de cada hexágono que forma la red reacciona en un patrón particular. Juntas, estas células forman un sistema de coordenadas que permiten la navegación espacial. El posicionamiento de la célula del hipocampo se proyecta en la malla hexagonal de la corteza entorrinal. Esta malla es una representación del lugar donde estamos y las células del hipocampo se proyectan sobre esta malla para que recordemos nuestra posición.
2. Gran conectividad entre hipocampo y corteza
El hipocampo recibe mucha información cortical, de hecho, una buena parte de la energía del cerebro en reposo que se gasta es la conexión entre la corteza y el hipocampo. Esto ocurre porque la corteza está continuamente organizando planes y comparándolos con la memoria del hipocampo para contrastarlos y volver a reorganizarlos en el hipocampo. Se utiliza constantemente la información de la memoria para organizar correctamente los planes. Esto ocurre porque las cortezas de asociación proyectan sus axones sobre el
3. Funciones superiores y plasticidad sináptica
El encéfalo adulto debe poseer la plasticidad necesaria para aprender habilidades nuevas y establecer memorias.
El aprendizaje y la memoria se deben a cambios en la eficacia de las sinapsis existentes.
En encéfalos muy complejos, es difícil atribuir un cambio conductual a modificaciones en las propiedades sinápticas de neuronas específicas. Este es el caso del ser humano ya que poseemos 10^11 neuronas y están muy interconectadas entre ellas. Por ello, es mucho más fácil realizar estudios en invertebrados debido a que son más sencillos. Como hay 100 sinapsis por neuronas. Hay 10^14 sinapsis totales.
3.1. Plasticidad sináptica en invertebrados: modificación conductual
Eric Kandel, Nobel de fisiología y medicina en el año 2000 dijo que la memoria del aprendizaje era igual de esencial en todos los animales, por lo que se podía estudiar en especies más simples en vez de en el ser humano. Él realizó sus estudios sobre la babosa de mar de la especie
Aplysia californica, que tiene únicamente 20000 neuronas y que a pesar de ser un valor mucho menor que el de los seres humanos, tiene el mismo funcionamiento.
Debido a la presunción de que la plasticidad sináptica es tan fundamental que sus bases moleculares y celulares están conservadas en sistemas nerviosos de organismos muy diferentes, se pudo estudiar la plasticidad conductual en la babosa, es decir, su habituación y sensibilización.
Frecuencias a las que no te puedes habituar: llanto niño.
La babosa tiene pocas neuronas e identificables, por lo que son muy fáciles de estudiar porque hay neuronas con incluso 1 mm de diámetro. Por ello, lo que hacía Eric Kandel en su estudio era tocar el sifón y observar cómo se movían las aletas del animal. Al cabo de un rato el animal ya no notaba el estímulo. Esto se conoce como la habituación a un estímulo, que es lo que nos pasa a nosotros cuando nos ponemos calcetines y dejamos de notar que los llevamos.
En el experimento, cuando el animal se habituaba, lo asociaban a un shock eléctrico (sensibilización como base fundamental de la asociación en la babosa) para que la babosa volviera a notar el estímulo antes de volver a habituarse. Si esto se repite continuamente se puede llegar a volver permanentemente sensibilizado o habituado.
Circuito neuronal simple de la contracción de la aleta de Aplysia. La actividad sináptica de este circuito se modifica durante la habituación y la sensibilización
3.1.1. Habituación
En la piel del sifón hay una neurona sensorial que envía una transmisión glutamatérgica. La neurona sensorial sigue respondiendo cada vez que hay contacto, pero el potencial sináptico de la motoneurona es lo que desciende cada vez más.
Cuando hay contacto con el sifón la aleta se contrae.
En la habituación, la transmisión sináptica glutamatérgica está disminuida por una reducción de la cantidad de vesículas sinápticas disponibles, lo que se conoce como depresión sináptica. Es decir, las vesículas sinápticas están preparadas, pero conforme hay más estímulos la cantidad de ellas comienza a disminuir, por lo que cada vez su liberación es menor y tiene lugar la habituación.
1. El mecanismo responsable del aumento de la transmisión glutamatérgica durante la sensibilización a corto plazo consiste en la modificación postraduccional de proteínas.
Motoneurona: verde / Neurona sensorial: azul / interneurona facilitadora: amarillo
Cuando hay habituación, las vesículas sinápticas de la neurona sensitiva se gastan y no llegan a la motoneurona. Con el choque eléctrico en la cola, se activa la interneurona facilitadora y hace un contacto previo con la neurona sensitiva, que libera serotonina (neurotransmisor rojo) en vesículas. Las moléculas de serotonina llegan al receptor de serotonina acoplado a proteína G, que fosforila la adenilato ciclasa, provocando que se libere AMPc. Este AMPc activa a la PKA, que fosforila otras proteínas modificando su función. Cuando se activa la PKA, fosforila un canal de potasio, haciendo que baje su probabilidad de apertura. Al no salir potasio, el potencial de reposo de la célula se eleva y la neurona se despolariza. Este aumento del potencial de membrana hace que entre calcio por canales de calcio dependientes de voltaje y se reclutan vesículas sinápticas diferentes a las iniciales, que liberan glutamato (neurotransmisor azul) sobre la motoneurona. Por tanto, se restaura la transmisión sináptica. En este modelo tiene que haber 2 tipos de vesículas, las iniciales y las que se reclutan después.
Se agotan vesículas sinápticas Choque eléctrico: sinapsis interneurona serotoninérgica Proteína G - adenilato ciclasa - AMPc - PKA PKA fosforila canal de potasio: + Vm se abren canales de calcio dependientes de voltaje se reclutan otras vesículas sinápticas.
2. Mecanismo responsable del aumento de la transmisión glutamatérgica durante la sensibilización a largo plazo: modificación de la expresión génica y de síntesis de proteínas.
No solo se modifican los canales de K+ sino que el AMPc se une a CREB y se modifica la expresión génica de las proteínas.
A largo plazo, se activa un factor de transcripción (CREB), que modifica la expresión de los genes haciendo que la sensibilización sea permanente por los continuos choques eléctricos. Debido a la liberación de AMPc provocada por la sinapsis serotoninérgica de la interneurona facilitadora, se activa un factor de transcripción (CREB) que modifica la transcripción génica en la neurona sensorial. Gracias a la expresión génica modificada por CREB, se activa la enzima ubiquitina hidroxilasa, que degrada la subunidad reguladora de la PKA: en lugar de estar activa solo durante un corto periodo de tiempo la PKA, se activa de forma permanente y ya no es un efecto transitorio. La duración de esta sensibilización puede comprender la vida entera del animal. Con la expresión génica también se generan proteínas no identificadas responsables del crecimiento de la sinapsis: aparecen nuevas espinas dendríticas en mamíferos para que la unión sináptica sea más potente que antes (ya que se realizan más sinapsis).
4. Plasticidad sináptica a largo plazo en sistema nervioso de mamíferos
Algunos patrones de actividad sináptica en circuitos encefálicos del cerebro de mamíferos producen potenciación a largo plazo (LTP) de las sinapsis, mientras que otros patrones generan depresión a largo plazo (LTD). La LTP se ha estudiado con mayor detalle en las sinapsis excitadoras del hipocampo: área del encéfalo especialmente importante para la formación y recuperación de algunas formas de memoria.
En el experimento midieron potenciales de acción por segundo en vez de Hercios, pero ambas unidades de medida son equivalentes.
4.1. Plasticidad sináptica a largo plazo en las sinapsis del hipocampo
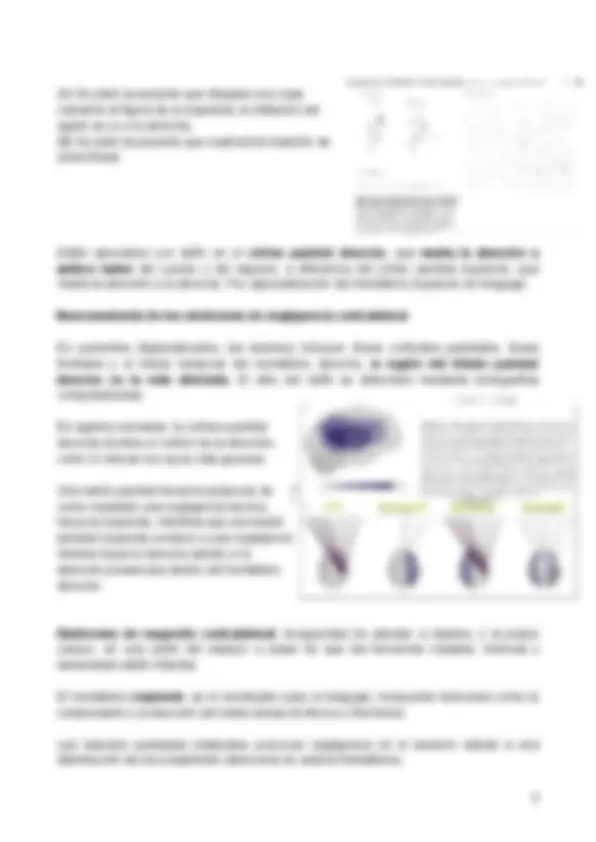
Como realizar estudios en animales completos es muy complicado, se usan como alternativa los estudios en rodajas de hipocampo, ya que se mantiene su estructura y el circuito neuronal a lo largo del lóbulo temporal. En este otro experimento, el objetivo es estudiar la potenciación sináptica entre CA3 y CA1, para ver si esta es específica de la vía que estimulan. Para ello, se realiza un registro intracelular que mide las sinapsis excitadoras entre axones de células piramidales en CA3 (colateral de Schaffer) y dendritas de piramidales de CA1.
El experimento consiste en el registro intracelular de las células piramidales de CA1 para ver el tamaño de los potenciales sinápticos excitadores (EPSPs) que se producen cuando estimulamos con 2 electrodos 2 vías diferentes (colaterales de Schaffer), ambas provenientes de células piramidales de CA3.
La diferencia entre ambas vías es que la vía 1 se estimula a alta frecuencia durante un período de tiempo y la vía 2 siempre es un estímulo igual, de frecuencia baja.
En primer lugar, se estimulan las dos vías a baja frecuencia y ven que el potencial sináptico tiene un valor equivalente. Después, estimulan sólo la 1a vía a alta frecuencia y ven que esta se queda potenciada durante 1h, mientras que la 2a sigue estimulándose a baja frecuencia. Con esto se demuestra que la modificación de la eficacia sináptica es específica para la vía que estamos analizando, es decir, que si alteramos sólo una vía, veremos que la respuesta sólo cambia en esa vía y no en la otra. Además, también explica que si la estimulación es de alta frecuencia, se produce LTP (aumento de los EPSPs).
5. Mecanismos moleculares que subyacen a la LTP
Los antagonistas de los receptores de glutamato del tipo NMDA (N-metil diaspartato) bloquean la LTP y no tienen efecto sobre la transmisión sináptica de baja frecuencia. Esto se debe a las propiedades biofísicas singulares del receptor NMDA: permeable a Ca2+ y bloqueado por Mg2+ extracelular.
El glutamato se une a 2 tipos de receptores:
**- Receptor AMPA: inotrópico permeable a sodio
- Receptor NMDA: ionotrópico permeable a sodio y calcio.** En condiciones normales no funciona bien, ya que se abre poco.
El magnesio extracelular presente en la sangre y tejidos se siente atraído por la carga negativa de la célula postsináptica y tapa el receptor NMDA, haciendo que entre poco sodio. Si se estimula a alta frecuencia, liberamos más glutamato, se producen más potenciales sinápticos y estos se van sumando. El potencial membrana de la postsináptica estará más despolarizado que si el estímulo fuera a baja frecuencia, por lo que el magnesio que estaba mantenido por las cargas electrostáticas, se repele y deja de bloquear, por lo que entra mucho calcio. Cuando entra el calcio en la neurona postsináptica, activa 2 proteínas quinasas: PKC y calcio-calmodulina quinasa II (CaMKII) En el citoplasma de la neurona postsináptica tenemos vesículas que secuestran receptores AMPA adicionales. Gracias a la activación de las quinasas, se liberan a la membrana los receptores AMPA de la vesícula, por lo que tendremos más receptores de tipo AMPA que de NMDA.
Por tanto, si estimulamos a baja frecuencia, como ahora tenemos más canales AMPA, la respuesta será mayor que al inicio sin estos canales adicionales. La respuesta por tanto se potencia y esta es la base del aprendizaje.
- un estímulo a alta frecuencia es el que nos permite que se potencie la sinapsis y que por tanto, desarrollemos el aprendizaje y la memoria.
- La entrada de Ca2+ por los receptores de NMDA sirve como señal intracelular para inducir la LTP.
- Activación de dos tipos de proteínas quinasas: CaMKII y PKC que promueven la inserción de receptores AMPA adicionales.
- La CaMKII es muy importante: enzima más abundante en las sinapsis del hipocampo y si se bloquea, se impide la LTP.
6. Cambios en la expresión génica durante la LTP
Cambios postraduccionales en fase temprana y cambios en la expresión génica en la fase más tardía (inhibida por bloqueo de síntesis proteica) de la plasticidad sináptica.
La PKC y la CaMKII no sólo producen la liberación de los receptores AMPA, sino que también fosforilan para sintetizar proteínas de crecimiento sináptico.
- PKC y CAMKII fosforilan subunidad de proteína G, haciendo que se sintetice AMPc
- AMPc activa a PKA