¡Descarga Guyton capítulo 10 versión de impresión. y más Apuntes en PDF de Fisiología solo en Docsity!
Capítulo 10: Excitación rítmica del
corazón. Páginas 123-128.
El corazón humano tiene un sistema especial para la autoexcitación rítmica y la contracción repetitiva aproximadamente 100.000 veces al día, o 3.000 millones de veces en una vida humana de duración media. Este impresionante logro es realizado por un sistema que: 1) genera impulsos eléctricos rítmicos para iniciar la contracción rítmica del músculo cardíaco, y 2) conduce estos estímulos rápidamente por todo el corazón. Cuando este sistema funciona normalmente, las aurículas se contraen aproximadamente 1/6 de segundo antes de la contracción ventricular, lo que permite el llenado de los ventrículos antes de que bombeen la sangre a través de los pulmones y de la circulación periférica. Este sistema también es importante porque permite que todas las porciones de los ventrículos se contraigan casi simultáneamente, lo que es esencial para una generación de presión más eficaz en las cavidades ventriculares. Este sistema rítmico y de conducción del corazón se puede lesionar en las cardiopatías, especialmente en la isquemia de los tejidos cardíacos que se debe a una disminución del flujo sanguíneo coronario. La consecuencia es con frecuencia una alteración del ritmo cardíaco o una secuencia anormal de contracción de las cavidades cardíacas, con una posible alteración grave de la eficacia de la función de bomba del corazón, incluso hasta el grado de producir la muerte.
Sistema de excitación especializado y de conducción del corazón
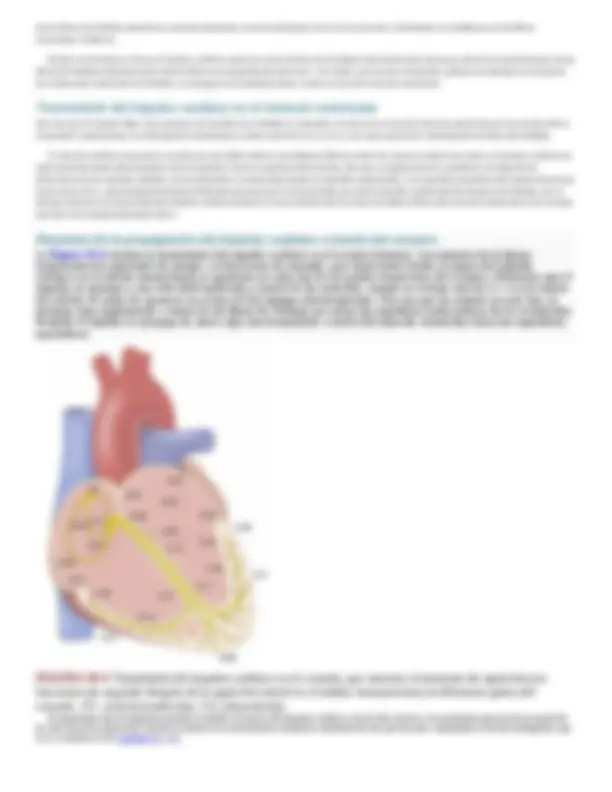
La figura 10-1 muestra el sistema especializado de excitación y conducción del corazón que controla las contracciones cardíacas. La figura muestra el nódulo sinusal (también denominado nódulo sinoauricular o SA), en el que se genera el impulso rítmico normal; las vías internodulares que conducen impulsos desde el nódulo sinusal hasta el nódulo auriculoventricular (AV); el nódulo AV, en el cual los impulsos originados en las aurículas se retrasan antes de penetrar en los ventrículos; el haz AV, que conduce impulsos desde las aurículas hacia los ventrículos, y las ramas izquierda y derecha del haz de fibras de Purkinje, que conducen los impulsos cardíacos por todo el tejido de los ventrículos.
FIGURA 10-1 Nódulo sinusal y sistema de Purkinje del corazón, que muestra también el
nódulo auriculoventricular (AV), las vías internodulares auriculares y las ramas de los haces
ventriculares.
Nódulo sinusal (sinoauricular) El nódulo sinusal (también denominado nódulo sinoauricular ) es una banda elipsoide, aplanada y pequeña de músculo cardíaco especializado de aproximadamente 3 mm de anchura, 15 mm de longitud y 1 mm de grosor. Está localizado en la pared posterolateral superior de la aurícula derecha, inmediatamente inferior y ligeramente lateral a la desembocadura de la vena cava superior. Las fibras de este nódulo casi no tienen
filamentos musculares contráctiles y cada una de ellas tiene solo de 3 a 5 μm de diámetro, en contraste con un diámetro de 10 a 15m de diámetro, en contraste con un diámetro de 10 a 15 μm de diámetro, en contraste con un diámetro de 10 a 15m para las fibras musculares auriculares circundantes. Sin embargo, las fibras del nódulo sinusal se conectan directamente con las fibras musculares auriculares, de modo que todos los potenciales de acción que comienzan en el nódulo sinusal se propagan inmediatamente hacia la pared del músculo auricular.
Ritmicidad eléctrica automática de las fibras sinusales
Algunas fibras cardíacas tienen la capacidad de autoexcitación, que es un proceso que puede producir descargas y contracciones rítmicas automáticas. Esta capacidad es especialmente cierta en el caso de las fibras del sistema especializado de conducción del corazón, entre ellas las fibras del nódulo sinusal. Por este motivo el nódulo sinusal habitualmente controla la frecuencia del latido de todo el corazón, como se analiza en detalle más adelante en este mismo capítulo. En primer lugar se va a describir esta ritmicidad automática. Mecanismo de la ritmicidad del nódulo sinusal La figura 10-2 muestra potenciales de acción registrados desde el interior de una fibra del nódulo sinusal durante tres latidos cardíacos y, a modo de comparación, un único potencial de acción de una fibra muscular ventricular. Obsérvese que entre descargas el «potencial de membrana en reposo» de la fibra del nódulo sinusal tiene una negatividad de aproximadamente –55 a –60 mV, en comparación con –85 a –90 mV para la fibra muscular ventricular. La causa de esta menor negatividad es que las membranas celulares de las fibras sinusales son permeables naturalmente a los iones sodio y calcio, y las cargas positivas de los iones sodio y calcio que entran neutralizan parte de la negatividad intracelular.
FIGURA 10-2 Descarga rítmica de una fibra del nódulo sinusal. Además, se compara el potencial de acción del nódulo
sinusal con el de una fibra muscular ventricular.
Antes de intentar explicar la ritmicidad de las fibras del nódulo sinusal, en primer lugar se debe recordar el análisis de los capítulos 5 y 9 de que el músculo cardíaco tiene tres tipos de canales iónicos de membrana que tienen funciones importantes en la generación de los cambios de voltaje en el potencial de acción. Los tipos son: 1) los canales rápidos de sodio; 2) los canales de calcio de tipo L (canales lentos de sodio-calcio), y 3) los canales de potasio. La apertura de los canales rápidos de sodio durante algunas diezmilésimas de segundo es responsable de la rápida espiga ascendente del potencial de acción que se observa en el músculo ventricular, debido a la entrada rápida de iones sodio positivos hacia el interior de la fibra. Después, la «meseta» del potencial de acción ventricular está producida principalmente por la apertura más lenta de los canales lentos de sodio- calcio, que dura aproximadamente 0,3 s. Finalmente, la apertura de los canales de potasio permite la difusión de grandes cantidades de iones potasio positivos hacia el exterior a través de la membrana de la fibra y devuelve el potencial de membrana a su nivel de reposo. Sin embargo, hay una diferencia en la función de estos canales en la fibra del nódulo sinusal porque el potencial «de reposo» es mucho menos negativo (de solo –55 mV en la fibra nodular, en lugar de los –90 mV de la fibra muscular ventricular). A este nivel de –55 mV, los canales rápidos de sodio principalmente ya se han «inactivado», lo que significa que han sido bloqueados. La causa de esto es que siempre que el potencial de membrana es menos negativo de aproximadamente –55 mV durante más de algunos milisegundos, las compuertas de inactivación del interior de la membrana celular que cierran los canales rápidos de sodio se cierran y permanecen de esta manera. Por tanto, solo se pueden abrir los canales lentos de sodio-calcio (es decir, se pueden «activar») y, por tanto, pueden producir el potencial de acción. En consecuencia, el potencial de acción del nódulo auricular se produce más lentamente que el potencial de acción del músculo ventricular. Además, después de la producción del potencial de acción, el regreso del potencial a su estado negativo también se produce lentamente, en lugar del regreso súbito que se produce en la fibra ventricular. Autoexcitación de las fibras del nódulo sinusal Debido a la elevada concentración de iones sodio en el líquido extracelular en el exterior de la fibra nodular, así como al número moderado de canales de sodio abiertos previamente, los iones sodio positivos del exterior de las fibras normalmente tienden a desplazarse hacia el interior. Por tanto, entre los latidos cardíacos, la entrada de iones sodio de carga positiva produce una elevación lenta del potencial de membrana en reposo en dirección positiva. Así, como se muestra en la figura 10-2 , el potencial «en reposo» aumenta gradualmente y se hace menos negativo entre cada
FIGURA 10-3 Organización del nódulo auriculoventricular (AV). Los números representan el intervalo de tiempo que
transcurre desde el origen del impulso en el nódulo sinusal. Los valores se han extrapolado a seres humanos.
Así, el retraso total en el nódulo AV y en el sistema de AV es de aproximadamente 0,13 s. Este retraso, añadido al retraso inicial de la conducción de 0,03 s desde el nódulo sinusal hasta el nódulo AV, hace que haya un retraso total de 0,16 s antes de que la señal excitadora llegue finalmente al músculo ventricular que se está contrayendo. Causa de la conducción lenta La conducción lenta en las fibras transicionales, nodulares y penetrantes del haz AV está producida principalmente por la disminución del número de uniones en hendidura entre células sucesivas de las vías de conducción, de modo que hay una gran resistencia a la conducción de los iones excitadores desde una fibra de conducción hasta la siguiente. Por tanto, es fácil ver por qué se tarda en excitar células sucesivas. Transmisión rápida en el sistema de Purkinje ventricular Las fibras de Purkinje especiales se dirigen desde el nódulo AV a través del haz AV hacia los ventrículos. Excepto en la porción inicial de estas fibras, donde penetran en la barrera fibrosa AV, tienen características funcionales bastante distintas a las de las fibras del nódulo AV. Son fibras muy grandes, incluso mayores que las fibras musculares ventriculares normales, y transmiten potenciales de acción a una velocidad de 1,5 a 4 m/s, una velocidad aproximadamente seis veces mayor que la del músculo ventricular normal y 150 veces mayor que la de algunas de las fibras del nódulo AV. Esta velocidad permite una transmisión casi instantánea del impulso cardíaco por todo el resto del músculo ventricular. Se piensa que la rápida transmisión de los potenciales de acción por las fibras de Purkinje está producida por un gran aumento del nivel de permeabilidad de las uniones en hendidura de los discos intercalados entre las células sucesivas que componen las fibras de Purkinje. Por tanto, los iones pasan fácilmente de una célula a la siguiente, aumentando de esta manera la velocidad de la transmisión. Las fibras de Purkinje también tienen muy pocas miofibrillas, lo que significa que se contraen poco o nada durante la transmisión de los impulsos. Conducción unidireccional a través del haz AV Una característica especial del haz AV es la imposibilidad, excepto en estados anormales, de que los potenciales de acción viajen retrógradamente desde los ventrículos hacia las aurículas. Esta característica impide la reentrada de los impulsos cardíacos por esta ruta desde los ventrículos hacia las aurículas, permitiendo solo la contracción anterógrada desde las aurículas hacia los ventrículos. Además, se debe recordar que en todas las localizaciones excepto en el haz AV el músculo auricular está separado del músculo ventricular por una barrera fibrosa continua, de la que se muestra una porción en la figura 10-3. Esta barrera normalmente actúa como aislante para impedir el paso de los impulsos cardíacos entre el músculo auricular y ventricular a través de cualquier ruta distinta a la conducción anterógrada a través del propio haz AV. (En casos infrecuentes un puente muscular anormal penetra en la barrera fibrosa en otra localización distinta al haz AV. En estas condiciones el impulso cardíaco puede entrar en las aurículas desde los ventrículos y producir arritmias cardíacas graves.) Distribución de las fibras de Purkinje en los ventrículos: las ramas izquierda y derecha del haz Después de penetrar en el tejido fibroso que está entre el músculo auricular y ventricular, la porción distal del haz AV se dirigía hacia abajo en el interior del tabique interventricular a lo largo de 5 a 15 mm hacia la punta del corazón, como se muestra en las figuras 10-1 y 10-3. Después el haz se divide en las ramas izquierda y derecha del haz, que están debajo del endocardio en los dos lados respectivos del tabique interventricular. Cada una de las ramas se dirige hacia abajo, hacia la punta del ventrículo, dividiéndose progresivamente en ramas más pequeñas. Estas ramas, a su vez, siguen un trayecto en dirección lateral alrededor de cada una de las cavidades ventriculares y hacia atrás, hacia la base del corazón. Los extremos
de las fibras de Purkinje penetran en aproximadamente un tercio del grosor de la masa muscular y finalmente se continúan con las fibras musculares cardíacas. Desde el momento en el que el impulso cardíaco entre las ramas del haz en el tabique interventricular hasta que sale de las terminaciones de las fibras de Purkinje el tiempo total transcurrido es en promedio de solo 0,03 s. Por tanto, una vez que el impulso cardíaco ha entrado en el sistema de conducción ventricular de Purkinje, se propaga casi inmediatamente a toda la masa del músculo ventricular. Transmisión del impulso cardíaco en el músculo ventricular Una vez que el impulso llega a los extremos de las fibras de Purkinje se transmite a través de la masa del músculo ventricular por las propias fibras musculares ventriculares. La velocidad de transmisión es ahora solo de 0,3 a 0,5 m/s, una sexta parte de la velocidad de las fibras de Purkinje. El músculo cardíaco envuelve el corazón en una doble espiral, con tabiques fibrosos entre las capas en espiral; por tanto, el impulso cardíaco no viaja necesariamente directamente hacia el exterior, hacia la superficie del corazón, sino que se angula hacia la superficie a lo largo de las direcciones de las espirales. Debido a esta inclinación, la transmisión desde la superficie endocárdica a la superficie epicárdica del ventrículo precisa hasta otros 0,03 s, aproximadamente igual al tiempo necesario para la transmisión por toda la porción ventricular del sistema de Purkinje. Así, el tiempo total para la transmisión del impulso cardíaco desde las ramas iniciales del haz hasta las últimas fibras del músculo ventricular en el corazón normal es de aproximadamente 0,06 s. Resumen de la propagación del impulso cardíaco a través del corazón La figura 10-4 resume la transmisión del impulso cardíaco en el corazón humano. Los números de la figura representan los intervalos de tiempo, en fracciones de segundo, que transcurren desde el origen del impulso cardíaco en el nódulo sinusal hasta su aparición en cada uno de los puntos respectivos del corazón. Obsérvese que el impulso se propaga a una velocidad moderada a través de las aurículas, aunque se retrasa más de 0,1 s en la región del nódulo AV antes de aparecer en el haz AV del tabique interventricular. Una vez que ha entrado en este haz, se propaga muy rápidamente a través de las fibras de Purkinje por todas las superficies endocárdicas de los ventrículos. Después el impulso se propaga de nuevo algo más lentamente a través del músculo ventricular hacia las superficies epicárdicas. FIGURA 10-4 Transmisión del impulso cardíaco en el corazón, que muestra el momento de aparición (en fracciones de segundo después de la aparición inicial en el nódulo sinoauricular) en diferentes partes del corazón. AV, auriculoventricular; SA, sinoauricular. Es importante que el estudiante aprenda en detalle el trayecto del impulso cardíaco a través del corazón y los momentos precisos de su aparición en cada una de las partes del corazón; es esencial un conocimiento cuantitativo detallado de este proceso para comprender la electrocardiografía, que se va a analizar en los capítulos 11 a 13.
músculo ventricular. Por el contrario, los nervios simpáticos se distribuyen en todas las regiones del corazón, con una intensa representación en el músculo ventricular, así como en todas las demás zonas. La estimulación parasimpática (vagal) ralentiza el ritmo y la conducción cardíacos La estimulación de los nervios parasimpáticos que llegan al corazón (los vagos) hace que se libere la hormona acetilcolina en las terminaciones nerviosas. Esta hormona tiene dos efectos principales sobre el corazón. Primero, reduce la frecuencia del ritmo del nódulo sinusal, y segundo, reduce la excitabilidad de las fibras de la unión AV entre la musculatura auricular y el nódulo AV, retrasando de esta manera la transmisión del impulso cardíaco hacia los ventrículos. Una estimulación vagal débil a moderada reduce la frecuencia del bombeo del corazón, con frecuencia hasta un valor tan bajo como la mitad de lo normal. Además, la estimulación intensa de los nervios vagos puede interrumpir completamente la excitación rítmica del nódulo sinusal o puede bloquear completamente la transmisión del impulso cardíaco desde las aurículas hacia los ventrículos a través del nódulo AV. En cualquiera de los casos, las señales excitadores rítmicas ya no se transmiten hacia los ventrículos. Los ventrículos pueden dejar de latir durante 5 a 20 s, pero después alguna área pequeña de las fibras de Purkinje, habitualmente en la porción del tabique interventricular del haz AV, presenta un ritmo propio y genera la contracción ventricular a una frecuencia de 15 a 40 latidos/min. Este fenómeno se denomina escape ventricular. Mecanismo de los efectos vagales La acetilcolina que se libera en las terminaciones nerviosas vagales aumenta mucho la permeabilidad de las membranas de las fibras a los iones potasio, lo que permite la salida rápida de potasio desde las fibras del sistema de conducción. Este proceso da lugar a un aumento de la negatividad en el interior de las fibras, un efecto que se denomina hiperpolarización, que hace que este tejido excitable sea mucho menos excitable, como se explica en el capítulo 5. En el nódulo sinusal, el estado de hiperpolarización hace el potencial de membrana «en reposo» de las fibras del nódulo sinusal mucho más negativo de lo habitual, es decir, de –65 a –75 mV en lugar del nivel normal de –55 a –60 mV. Por tanto, el aumento inicial del potencial de membrana del nódulo sinusal que produce la corriente de entrada de sodio y de calcio tarda mucho más en alcanzar el potencial liminal para la excitación. Este requisito retrasa mucho la frecuencia de ritmicidad de estas fibras nodulares. Si la estimulación vagal es lo suficientemente intensa es posible detener totalmente la autoexcitación rítmica de este nódulo. En el nódulo AV, el estado de hiperpolarización producido por la estimulación vagal hace que sea difícil que las pequeñas fibras auriculares que entran en el nódulo generen una corriente de una intensidad suficiente como para excitar las fibras nodulares. Por tanto, el factor de seguridad para la transmisión del impulso cardíaco a través de las fibras de transición hacia las fibras del nódulo AV disminuye. Una reducción moderada simplemente retrasa la conducción del impulso, aunque una disminución grande bloquea totalmente la conducción. La estimulación simpática aumenta el ritmo y la conducción del corazón La estimulación simpática produce esencialmente los efectos contrarios sobre el corazón a los que produce la estimulación vagal, como se señala a continuación. Primero, aumenta la frecuencia de descarga del nódulo sinusal. Segundo, aumenta la velocidad de conducción, así como el nivel de excitabilidad de todas las porciones del corazón. Tercero, aumenta mucho la fuerza de contracción de toda la musculatura cardíaca, tanto auricular como ventricular, como se analiza en el capítulo 9. En breve, la estimulación simpática aumenta la actividad global del corazón. La estimulación máxima casi puede triplicar la frecuencia del latido cardíaco y puede aumentar la fuerza de la contracción del corazón hasta dos veces. Mecanismo del efecto simpático La estimulación de los nervios simpáticos libera la hormona noradrenalina en las terminaciones nerviosas simpáticas. La noradrenalina estimula, a su vez, los receptores β 1 -adrenérgicos, que median en los efectos sobre la frecuencia cardíaca. No está del todo claro el mecanismo preciso mediante el que la estimulación β 1 -adrenérgica actúa sobre las fibras del músculo cardíaco, aunque se piensa que aumenta la permeabilidad de la membrana de las fibras a los iones sodio y calcio. En el nódulo sinusal, un aumento de la permeabilidad a sodio-calcio genera un potencial en reposo más positivo y también produce un aumento de la velocidad del ascenso del potencial de membrana diastólico hacia el nivel liminal para la autoexcitación, acelerando de esta forma la autoexcitación y, por tanto, aumentando la frecuencia cardíaca. En el nódulo AV y en los haces AV, el aumento de la permeabilidad a sodio-calcio hace que sea más fácil que el potencial de acción excite todas las porciones sucesivas de los haces de las fibras de conducción, disminuyendo de esta manera el tiempo de conducción desde las aurículas hasta los ventrículos. El aumento de la permeabilidad a los iones calcio es responsable al menos en parte del aumento de la fuerza contráctil del músculo cardíaco bajo la influencia de la estimulación simpática, porque los iones calcio tienen una función importante en la excitación del proceso contráctil de las miofibrillas.