Vista previa parcial del texto
¡Descarga HERPESVIRUS y más Apuntes en PDF de Microbiología solo en Docsity!
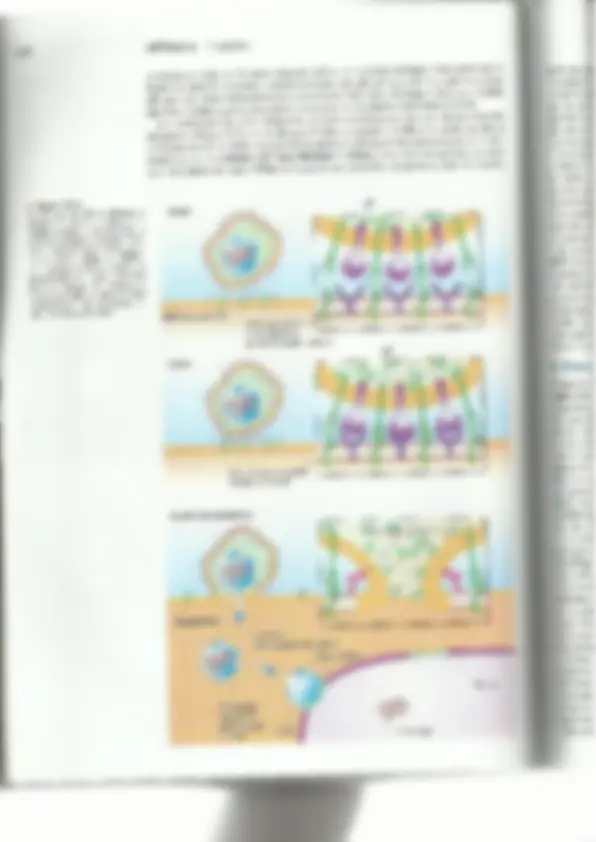
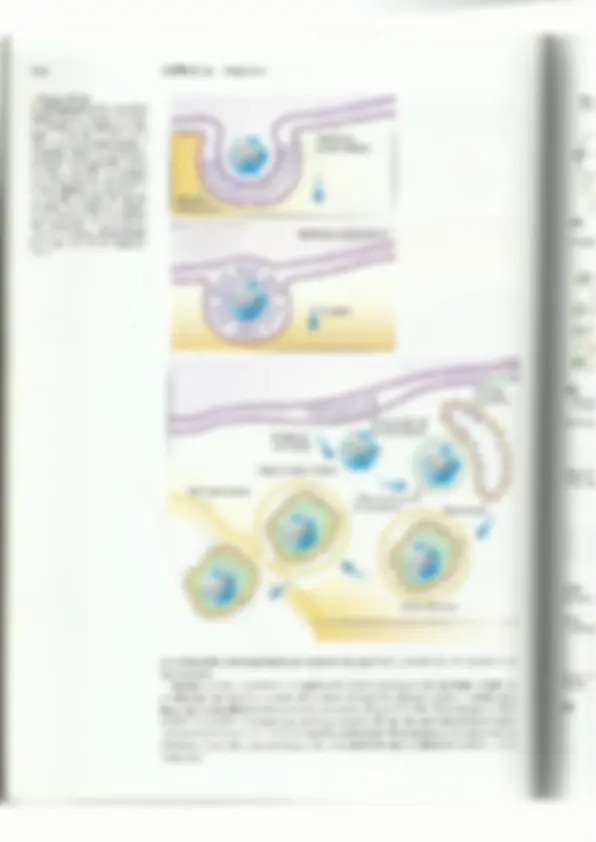
15.4 Ciclo de vida de los herpesvirus 417 Pruebas de diagnóstico más comunes para las infeccio- nes por herpesvirus E Muestra Pruebas de laboratorio Lesiones cutáneas Cultivo viral y amplificación del DNA Lesiones genitales Cultivo viral Lesiones cutáneas Amplificación del DNA Sangre o suero Serología para diagnosticar una enfermedad similar a la mononucleosis 3 Qin Cultivo viral, serología y amplificación del DNA £ para diagnosticar enfermedad neonatal Amplificación del DNA para diagnosticar infec- ciones sistémicas en inmunocomprometidos Amplificación del DNA para diagnosticar infec- ción del sistema nervioso central EBV Sangre o suero Serología para diagnosticar enfermedades seme- jantes a la mononucleosis Sangre o suero Serología para diagnosticar la sexta enfermedad Lesiones del sarcoma de Kaposi Amplificación del DNA Cultivo de la herida Cultivo viral a partir del hisopado de las heridas o biopsia o líquido del sistema nervioso central (se requieren instalaciones BSL-4) Líquido cefalorraquídeo Amplificación del DNA, ELISA Sangre o suero m Estructura y clasificación de los herpesvirus Los herpesvirus son partículas grandes, envueltas y pleo- 'mórficas que tienen un diámetro de 150 a 300 nm cuando se as examina con microscopio electrónico. Los viriones de los herpesvirus comprenden más de 30 proteínas codificadas por el virus y también contienen componentes celulares. La cu- bierta viral se obtiene a través de un proceso de doble envol- tura. Ésta es frágil y se daña fácilmente por el calor, la dese- cación, el alcohol al 70% o la alteración por jabón o deter- gentes. Para que los herpesvirus sean infecciosos, la envoltu- ta debe estar intacta. Como es extremadamente sensible al yy Figura 15-4 daño. el virus generalmente se transmite mediante el contac- Fotomicrografía que muestra los cuerpos de inclusión intranucleares que o directo con superficies mucosas o secreciones de una per- produce el VZV en cultivos de tejidos (500%). sona infectada (como la boca, los labios o los genitales). Los herpesvirus se secan y se dañan cuando están expuestos al aire y por ello no pueden ser transmitidos mediante los asientos de los inodoros u otros objetos inanimados, una creencia errónea. Los viriones tienen genomas de DNA bicatenario (ds DNA) que varían entre 125 y 230 pa- tes de kilobases. El DNA del genoma está rodeado por una cápside icosahédrica, y una cubierta protcinácea amorfa circunda a la nucleocápside. El tegumento Consiste al menos en 8 proteínas virales diferentes (Figura 15-5). Hay 8 herpesvirus que infectan a los seres humanos en forma natural. Los adultos de mediana edad generalmente tienen anticuerpos contra la mayoría de éstos, con excepción del HHV-8 (que es común en los hombres HIV>). 418 CAPÍTULO 15 Herpesvirus Figura 15-5 a) Estructura esquemática de los herpesvirus. Hay más de Proyecciones una docena de proteínas integrales de membrana diferentes glucoproteicas en la envoltura viral. El tegumento contiene al menos 8 pro- teínas que tienen funciones importantes una vez que el vi- rus entra en el huésped. b) Fotomicrografía electrónica de varios virus de la familía Herpesviridae. Cubierta Nucleocáps3= Envoltura La familia Herpesviridac se divide en 3 subfamilias: Alfaherpesvirinac, Betahcrpestio nac y Gammaherpesvirinae. Las subfamilias se diferencian unas de otras por propiedad= como el tipo de células en las cuales se establece la latencia viral y las características d= su crecimiento en cultivos celulares (Cuadro 153). Los virus se designan de acuerdo la especie de su huésped natural y se los enumera en cl orden en el cual se los identi por ejemplo, el virus herpes simple tipo 1 (HSV-1) fue el primer herpesvirus que se ide lificó en los seres humanos. Es probable que en el futuro el esquema de clasificación de 420 CAPÍTULO 15 — Herpesvirus la envoltura viral con la membrana plasmática de la célula huésped. Esta actividad de fusión requiere de la acción de un receptor para 2B, 2D, gH-eL y gD. Las glucoproteínas gB, gH y gL están estructuralmente conservadas entre todos los herpesvirus y es probable que desempeñen papeles semejantes y esenciales en la entrada viral (Figura 15-6) Las nucleocápsides de los herpesvirus se hallan cubiertas por una capa de protcínas del tegumento (Figura 15-5). A medida que el virus se denuda y se libera la nucleocápside en el citoplasma de la célula, algunas de las proteínas del tegumento permanecen en el cito- plasma (p. ej., la proteína del “cancelamiento” celular, virus host shutoff-vhs), en tanto que otras proteínas como VPL6 son transportadas al núcleo. La proteína viral vhs desem- w Figura 15-6 La gB y 9C del HSV se adhieren al receptor de unión al proteglucano celular. Después, la gD del virión se une al correceptor de entrada HVEM. Esto facilita la fusión de las mem- branas viral y celular. A continua- ción, el genoma viral se denuda y es transportado al núcleo. (Adaptado de E. K. Wagner, HSV Replication, http://www.dbc.uci.edu/-faculty/ wagner/hsváf.html. Ilustración ori- ginal de Imagecyte.com.) roteoglucanos de la superficie celular (heparán sulfato) Familia de receptoréS HVEMTNF/NGF Fusión de membrana peña un papel en la degradación rápida de los RNA mensajeros celulares (mRNA) en el citoplasma, lo cual causa la represión o la regulación negativa de la sínte- sis de proteínas del huésped. En consecuencia, los 'ÍmRNA virales se acumulan en el citoplasma y sOn tra- ducidos de manera preferente. La VP16 se une al DNA y es un activador de la transcripción. La nucleocápside se acopla a la membrana nuclear y libera el genoma en el núcleo, donde se transcribe y se replica. Los genomas de los herpesvirus están com- puestos por DNA lincal bicatenario. En condiciones experimentales, el dsDNA es infeccioso. Su longitud y su complejidad difieren para cada virus. Los extre- mos del genoma consisten en secuencias repetitivas. El genoma de algunos herpesvirus contiene secuch- cias repetitivas únicas grandes y pequeñas (Figura 15-7). Estas secuencias codifican más de 50 productos proteicos distintos, como las enzimas para la replica- ción del DNA, las glucoproteínas que son importantes para la determinación del rango de huéspedes, las pro- tefnas para las defensas contra el huésped y dradas del genoma representan en la Figura cias repetitivas que se localizan en los extres m Replicación: infección productiva o lítica Después de la entrada, infección latente. La infección product recientemente en el núcleo se vuelve circul lar dependiente de DNA y regulado por las proteí por las células y por el virus. Para iniciar l se une a uno de los tres orígenes de rep! con la ayuda de una proteína de unión tinuación, el complejo de helicasa y pri ala proteína U, 42, se une a los cebadores 15-3). En algún momento, el DNA se re] círculo rodante, del cua] resultan conca circular (Figura 15-8). Un sello distintivo del genoma de los 'mas que se requieren para incrementar timidina cinasa, la ribonucleótido reductasa, dina trifosfatasa) y para replicar el DNA del virus se replique en enzimas mencionadas son dianas para de los genes del HSV-1 y del HI lares. Casi el 50% de los genes doras que participan en el estableci Como en los poxvirus, la “anscripción temporal y secuencial que llev: nos inmediatos (02), tempranos (B) y t viral que se acumula en las células después de l; genes 0%. Los genes 0 codifican las pi papel en la transcripción viral. Los genes B y factores virales de transcripción 15.4 Ciclo de vida de los herpesvirus DNA monocatenario y sintetiza cebadores de RNA. La DNA polimerasa viral U, 30, de RNA y comienza la síntesis de DNA (Figura plica mediante un mecanismo de replicación en témeros lineales de DNA a partir del un molde [SV-2 no es necesario p: sistema inmunitario del huésped (p. ej., las virocinas). de los herpesvirus se produce de una manera e 3 clases de genes virales: tempra- ardíos (y). La protcína viral VP16 (la 16”. proteína la infección) activa la expre: roteínas de unión al DNA, las cuales desempeñan un codifican factores de la replicación del DNA adicionales. Los genes y codifican a las proteínas a la transcripción de 421 a | UL Us 2 WN E $ 5 HSvA, UL Us, 2 Hsv2 A 2 U Y E . 6-08 2 E uu 2 HHVO 3 A 0 50 100 150 Pares de kilobases T Guadros = secuencias repetitivas | Us = secuencias únicas cortas 'ecuencias repetitivas largas al Figura 15-7 Comparación de los genomas de los herpesvirus. El genoma contiene secuer es nicas (1) y secuencias repetidas invertidas (recuadros). (Adaptado de Murray, P.R., et al. Medical Microbiology, Third Edition. Mosby Elsevier Health Science, 1998.) las protcínas de la cápside. Las regiones recua- 15-7 las secuencias repetitivas. Las secuen- mos del genoma viral intervienen en el proce- so de circularización y el empaquetamiento del DNA en el virión. el virus inicia una infección lítica, productiva, O establece una tiva tiene lugar cuando el genoma viral que entró lar y es transcrito por la RNA polimerasa celu- reguladoras nucleares codificadas a replicación del DNA del HSYV, la proteína UL? licación (ORT) posibles, La U, 9 desenrolla al DNA. al DNA monocatenario denominada ICP8. A con- ¡masa que consiste en U,5, U,8 y U, 52 se une al mida herpesvirus es que codifica la mayoría de las enzi- a reserva de nucleótidos en una célula (p. ej.. la la uracilo DNA glucosilasa y la desoxiuri- genoma. Esta característica permito que el células de división lenta o que no se dividen, como las neuronas, Las los fármacos antivirales. Aproximadamente, el 50% ara la replicación en cultivos celu- del HSV-1 y del HSV-2 puede codificar proteínas regula- miento de la latencia y en la supresión O ev asión del n de los El virus se une a receptores específicos sobre la superti- cie de la célula huésped La envoltura yla y la mem- aia AE se po Fusión Adhesión Entrada dela cubierta La nucleocápside y las proteínas de la | cubierta entran en el | citoplasma celular La nucleocápside migra a través del citoplasma hasta el poro nuclear Membrana OM MANA Transcripción y traducción Síntesis de proteína - Proteínas 11123A — (temprana inmediata) Síntesis de proteínas b (temprana) DNA circularizado Proteínas 11123B' p as Síntesis de iétodo “de Ñ proteínas g (tardía) circulo rodante' Proteínas 11123G Ñ me Glucoproteínas W y Golgi Proteínas 0 de la cubierta 15.4 Ciclo de vida de los herpesvirus — 423 1 Figura 15-9 Después de la entrada, el genoma se circulari- za y tanto las proteínas del tegumento viral como factores celulares participan en la trans- cripción del genoma viral. La transcripción y la traducción se producen en 3 fases: (1) tem- prana inmediata; B) temprana, y y) tardía. (Adaptado de J. Cohen y W. G. Powderly. Infectious Diseases, Second Edition. Mosby, 2003.) 424 a Figura 15-10 La morfogénesis de los herpesvirus requiere de los procesos de desen- volvimiento y reenvoltura. La mem- brana nuclear envuelve al virión y esto transloca la nucleocápside al citoplasma, donde el virus madura. La mayoría de las proteínas del te- gumento (p. ej. aTIF, vhs) se agrega a las nucelocápsides y la envoltura final es adquirida por brotación en las vesículas derivadas del aparato de Golgi que contienen glucopro- teínas. (Adaptado de E. K. Wagner, HSV. Replication, http://ww.dbe, uci.edu/«faculty/wagner/hsváf. html. Ilustración original de Imagecyte. com.) CAPÍTULO 15 Herpesvirus J Membrana nuclear externa y Membrana nuclear interna; 4 : Envoltura Vesícula exocitótica Translocación de nn la nucleocápside Pérdida de la envoltura Cápside citoplasmática 3 o a os PS eTIF y vhs (proteínas del tegumento) Reenvoltura Membrana celular mo antisentido y desempeñarían de tal modo un papel en la prevención de la apoptosis de las neuronas. Mientras el virus está latente, la replicación viral no prosigue y las partículas virales no se detectan. El virus sc esconde del sistema inmunitario durante meses e incluso años hasta que se reactiva en una infección productiva (Figura 15-11b). Una vez que el VZW. el HSV-1 y el HSV-2 se reactivan, se dirigen a través de las vías nerviosas hacia la super ficic de la piel (Figura 15-11c). Los detalles moleculares de la latencia y reactivación son diferentes para cada grupo de herpesvirus y se demostró que es difícil de analizar a nivel molecular. Reintes de infes ) Reacti Superfic de la pie ibra — erviosa lirus — activadi